¡Descarga Mecanismos de Transcripción y más Resúmenes en PDF de Biología Molecular solo en Docsity!
CAPÍTULO 13
Mecanismos de
Transcripción
U
P A ESTE PUNTO, HEMOS ESTADO CONSIDERANDO mantenimiento de la ge- CONTORNO
nome, es decir, cómo se organiza, protege y replica el material
genético. Pasamos ahora a la cuestión de cómo es ese material
genético expresadoes decir, cómo la serie de bases del ADN dirige la
producción de los ARN y las proteínas que realizan funciones celulares y
definen la identidad celular. En los siguientes capítulos, describimos los
procesos básicos responsables de la expresión génica: transcripción,
procesamiento de ARN y traducción.
La transcripción es, química y enzimáticamente, muy similar a la replicación del
ADN (capítulo 9). Ambos involucran enzimas que sintetizan una nueva hebra de
ácido nucleico complementaria a una hebra molde de ADN. Hay algunas diferencias
importantes, por supuesto; más notablemente, en el caso de la transcripción, la
nueva hebra está hecha deribonucleótidos en vez de desoxirribonucleótidos (ver el
Capítulo 5). Otras características mecánicas de la transcripción que difieren de la
replicación incluyen las siguientes.
ARN polimerasas y la transcripción
Ciclo, 430
El ciclo de transcripción en bacterias,
Transcripción en eucariotas, 448
Transcripción por ARN polimerasas
I y III, 462
Visite el contenido web para ver tutoriales estructurales y animaciones interactivas
† Polimerasa de ARN (la enzima que cataliza la síntesis de ARN) no
necesita una cartilla; más bien, puede iniciar la transcripción de novo (aunque in
vivo, la iniciación está permitida sólo en ciertas secuencias, como veremos).
† El producto de ARN no permanece emparejado con el ADN molde.
hebra: la enzima desplaza la cadena en crecimiento solo unos pocos nucleótidos
detrás de donde se agrega cada ribonucleótido (Fig. 13-1). Este desplazamiento
es fundamental para que el ARN realice sus funciones (p. Ej., Como suele ser el
caso, para que se traduzca para producir su producto proteico). Además, debido
a que esta liberación sigue muy de cerca el sitio de polimerización, múltiples
moléculas de ARN polimerasa pueden transcribir el mismo gen al mismo tiempo,
cada una siguiendo de cerca a la otra. Por tanto, una célula puede sintetizar una
gran cantidad de transcripciones de un solo gen (u otra secuencia de ADN) en
poco tiempo. Es importante señalar que a medida que el producto de ARN se
disocia de la plantilla de ADN justo detrás de cada ARN polimerasa que avanza,
las dos cadenas de ADN se vuelven a recocer (fig. 13-1).
† La transcripción, aunque muy precisa, es menos precisa que la replicación.
(Se produce un error en 10,000 nucleótidos agregados, en comparación con uno en 10 millones para la replicación). Esta diferencia refleja la falta de amplios mecanismos de corrección de pruebas para la transcripción, aunque existen dos formas de corrección de pruebas para la síntesis de ARN.
Traducido del inglés al español - www.onlinedoctranslator.com
430 Capítulo 13
Dúplex de ADN 5' 3'
5 ' 3 '
ARN 5 ' (^) hebra de plantilla
FIGURA 13-1 Transcripción de ADN en ARN. La figura muestra, en ausencia de las enzimas involucradas, cómo se desenrolla la doble hélice de ADN y se construye una hebra de ARN en la hebra molde. También muestra cómo la transcripción de ARN se disocia de la plantilla de ADN unos pocos nucleótidos detrás del punto de síntesis, y cómo las hebras de ADN se reasocian. En la figura, la transcripción procede de izquierda a derecha.
Tiene sentido que la célula se preocupe más por la precisión de la replicación que de la transcripción. El ADN es la molécula en la que se almacena el material genético y la replicación del ADN es el proceso mediante el cual se transmite ese material genético. Por lo tanto, cualquier error que surja durante la replicación puede ser fácilmente catastrófico: se vuelve permanente en el genoma de ese individuo y se transmite a las generaciones posteriores. La transcripción, por el contrario, produce sólo copias transitorias y normalmente varias de cada región transcrita. Por lo tanto, un error durante la transcripción rara vez causará más daño que hacer que una de las muchas transcripciones transitorias sea defectuosa.
Más allá de estas diferencias mecánicas entre la replicación y la
transcripción del ADN, una profunda diferencia refleja los diferentes propósitos
a los que sirven estos procesos. La transcripción copia selectivamente solo
ciertas partes del genoma y hace entre uno y varios cientos, o incluso miles, de
copias de una sección determinada. Por el contrario, la replicación debe copiar
todo el genoma y hacerlo una vez (y sólo una vez) en cada división celular
(como vimos en el capítulo 9). La elección de qué regiones transcribir no es
aleatoria: hay secuencias de ADN específicas que dirigen el inicio de la
transcripción al comienzo de cada región y otras al final que terminan la
transcripción.
No solo se transcriben diferentes partes del genoma en diferentes grados, sino que también se puede regular la elección de qué parte transcribir y en qué medida. Por tanto, en diferentes células, o en la misma célula en diferentes momentos, se podrían transcribir diferentes conjuntos de genes. Por lo tanto, por ejemplo, dos células genéticamente idénticas en un ser humano, en muchos casos, transcribirán diferentes conjuntos de genes, dando lugar a diferencias en el carácter y la función de esas dos células (por ejemplo, una podría ser una célula muscular y la otra una neurona). O una determinada célula bacteriana transcribirá un conjunto diferente de genes, dependiendo del medio en el que esté creciendo. Estas cuestiones de regulación transcripcional se tratan en la Parte 4.
POLIMERASAS DE ARN Y EL CICLO DE TRANSCRIPCIÓN
Las ARN polimerasas se presentan en diferentes formas pero comparten muchas características
La ARN polimerasa realiza esencialmente la misma reacción en todas las células, desde las bacterias hasta los humanos. Por tanto, no es sorprendente que las enzimas de estos organismos compartan muchas características, especialmente en aquellas partes de la enzima directamente implicadas en la catalización de la síntesis de ARN. Desde bacterias hasta mamíferos, las ARN polimerasas celulares están formadas por múltiples subunidades (aunque algunos fagos y orgánulos codifican enzimas de una sola subunidad que realizan la misma tarea, como veremos en el Cuadro 13-2). La tabla 13-1 muestra
432 Capítulo 13
FIGURA 13-2 Comparación de las estructuras cristalinas de las ARN polimerasas procariotas y eucariotas. (a) Estructura de la enzima del núcleo de la ARN polimerasa de Thermus aquaticus. Las subunidades están coloreadas de la siguiente manera: (azul) B; (púrpura) B 0 ; (amarillo y verde) los dosa subunidades; (rojo)v. El Mg2þ ion (bola roja) marca el sitio activo aquí y en la parte b (Seth Darst, The Rockefeller University, com. pers.). (b) Estructura del ARN Pol II de levaduraSaccharomyces cerevisiae. Las subunidades están coloreadas para mostrar su relación con las de la enzima bacteriana (véase el cuadro 13-1). Por lo tanto, RPB1 (violeta); RPB (azul); RPB3 (verde) RPB11 (amarillo); RPB6 (rojo). (De Cramer P. et al. 2001.Ciencias 292: 1863.) Imágenes preparadas conMolScript, BobScript y Raster3D.
y RPB1 y RPB2 para la enzima eucariota). El sitio activo, que está formado por
regiones de estas dos subunidades, se encuentra en la base de las pinzas dentro de
una región denominada “hendidura del centro activo” (v. Fig. 13-2). El sitio activo
funciona de acuerdo con el mecanismo catalítico de iones de dos metales para la
adición de nucleótidos propuesto para todos los tipos de polimerasa (ver Capítulo
9). En este caso, sin embargo, el sitio activo contiene solo un Mg fuertemente unido 2
þ ion, y el segundo Mg2þ se introduce con cada nuevo nucleótido en el ciclo de
adición y se libera con el pirofosfato.
Hay varios canales que permiten que el ADN, el ARN y los ribonucleótidos entren
y salgan de la hendidura del centro activo de la enzima. Discutiremos estos más
adelante al considerar los mecanismos de transcripción.
La transcripción por ARN polimerasa procede en una serie de pasos
Para transcribir un gen, la ARN polimerasa pasa por una serie de pasos bien
definidos agrupados en tres fases: iniciación, alargamiento, y terminación.Aquí,
y en la Figura 13-3, resumimos las características básicas de cada fase.
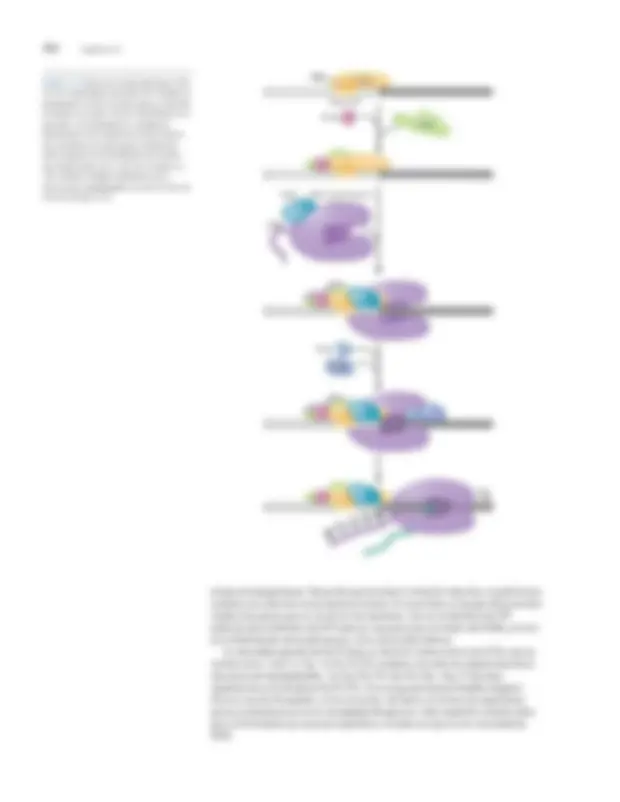
Iniciación Apromotor es la secuencia de ADN que inicialmente se une a la ARN polimerasa (junto con cualquier factor de iniciación requerido). Una vez formado, el complejo promotor- polimerasa sufre los cambios estructurales necesarios para que proceda la iniciación. Como en el inicio de la replicación, el ADN alrededor del punto donde comenzará la transcripción se desenrolla. Los pares de bases se rompen, produciendo una "burbuja de transcripción" de ADN monocatenario. Una vez más, como la replicación del ADN, la transcripción siempre ocurre en 5 0 -a-3 0 dirección: el nuevo ribonucleótido se añade a los 3 0 final de la cadena de crecimiento. Sin embargo, a diferencia de la replicación, solo una de las cadenas de ADN actúa como plantilla sobre la que se construye la cadena de ARN. Debido a que la ARN polimerasa se une a los promotores en una orientación definida, la misma hebra siempre se transcribe a partir de un promotor dado.
La elección del promotor determina qué tramo de ADN se transcribe y
es el paso más común en el que se impone la regulación.
Mecanismos de transcripción 433
ARN
polimerasa FIGURA 13-3 Las fases del ciclo de transcripción: inicio, alargamiento y terminación. La figura muestra el esquema general del ciclo de transcripción. Las características que se muestran son válidas tanto para casos bacterianos como eucariotas. Otros factores necesarios para la iniciación, elongación y la terminación no se muestran aquí, pero se describen en el texto. El nucleótido de ADN que codifica el comienzo de la cadena de ARN se denomina sitio de inicio de la transcripción y se designa como
“ÞPosición de 1 ”. Las secuencias en la dirección en la que procede la transcripción se denominan aguas abajo del sitio de inicio. Asimismo, las secuencias que preceden al sitio de inicio se denominan secuencias cadena arriba. Cuando se hace referencia a una posición específica en la secuencia ascendente, se le da un valor negativo. A las secuencias aguas abajo se les asignan valores positivos.
río arriba ADN
río abajo
promotor
vinculante (cerrado complejo)
promotor "derritiendo" (abierto complejo)
inicial transcribiendo complejo ARN
alargamiento
ARN
polimerasa termina y lanzamientos ARN
terminación
alargamiento
iniciación
Mecanismos de transcripción 435
FIGURA 13-4 Holoenzima de ARN polimerasa de Thermus aquaticus. En gris se muestra la enzima central (la misma enzima que se muestra en la figura 13-2a). loss 70 La subunidad se muestra en violeta (regiones 2, 3 y 4; consulte la figura 13-6). A la derecha está la región 2; en la parte superior, región 3; y en la parte inferior, la región 4. Como se describe más adelante en el texto, ess regiones 2 y 4 que reconocen las regiones –10 y –35 del promotor, respectivamente. (DeMurakami KS y col. 2002.Ciencias 296: 1280.) Imagen preparada con MolScript, BobScript y Raster3D.
cada uno de 6 nucleótidos, separados por un tramo inespecífico de 17 a 19 nucleótidos (fig. 13-5a). Las dos secuencias definidas están centradas, respectivamente, a 10 pb y 35 pb cadena arriba del sitio donde comienza la síntesis de ARN. Por tanto, las secuencias se denominan:35 (menos 35) y -10 (menos 10)
regiones, o elementos, de acuerdo con el esquema de numeración descrito en la
Figura 13-3, en el que el nucleótido de ADN que codifica el comienzo de la cadena
de ARN se designa þ1. Aunque la gran mayoría de s 70 los promotores contienen
regiones -35 y -10 reconocibles, las secuencias no son idénticas. Al comparar
muchos promotores diferentes,secuencia de consenso pueden derivarse (para un
análisis de cómo se obtienen, consulte el Recuadro 13-1, Secuencias de consenso).
La secuencia de consenso refleja las regiones preferidas –10 y –35, separadas por el
espaciado óptimo (17 pb). Muy pocos promotores tienen esta secuencia exacta,
pero la mayoría se diferencia de ella solo por unos pocos nucleótidos.
Los promotores con secuencias más cercanas al consenso son generalmente "más fuertes" que aquellos que no coinciden tan bien. Por la fuerza de un promotor, entendemos cuántas transcripciones inicia en un tiempo determinado. Esa medida está influenciada por qué tan bien se une el promotor a la polimerasa inicialmente, qué tan eficientemente apoya
a
(17-19 pb)
B
Elemento UP (^) - 35 - 10 + 1
C "Extendido –10"
FIGURA 13-5 Características de los promotores bacterianos. Se muestran varias combinaciones de elementos promotores bacterianos. Los detalles de cómo cada elemento contribuye a la unión y función de la polimerasa se describen en el texto.
D discriminado
436 Capítulo 13
} TÉCNICAS
BBUEY 13-1 Secuencias de consenso
Las secuencias de ADN de los sitios de unión reconocidos por una proteína administrada pueden no ser siempre exactamente iguales. Del mismo modo, un tramo de aminoácidos que confiere a una proteína una función particular puede ser ligeramente diferente en diferentes proteínas. Una secuencia consenso es, en cada caso, la aversión a la secuencia que tiene una posición en el nucleótido (o aminoácido) que se encuentra más comúnmente allí en diferentes ejemplos. Por tanto, la secuencia de consenso para los promotores enE. coli reconocida por la ARN polimerasa que contiene s 70 se muestra en la figura (Cuadro 13-1 Fig. 1). Esta secuencia de consenso se derivó alineando 300 secuencias que se sabe que funcionan comos 70 promotores y determinando la base más común encontrada en cada posición en los hexámeros –35 y –10. A continuación, ese nucleótido se elige como el nucleótido de elección en esa posición en el consenso; su frecuencia relativa y las frecuencias con las que los otros tres nucleótidos ocurren en cada posición se representan en el gráfico. Tenga en cuenta que no existe un consenso significativo entre los 17-19 nucleótidos que se encuentran en la región entre -35 y -10. En ese ejemplo, cada secuencia promotora individual se había identificado previamente, por lo que alinear las secuencias es tri-
frasco. Pero considere un ejemplo bastante diferente. En este caso, no se ha identificado ningún sitio de unión para la proteína de unión al ADN en cuestión. Sin embargo, se sabe que varias regiones de un cromosoma contienen sitios de unión en algún lugar dentro de sus longitudes. Se utiliza un algoritmo informático que escanea cada una de las secuencias de estas regiones cromosómicas, buscando un sitio de unión potencial común a todas ellas. Un segundo enfoque para derivar la secuencia de consenso para una proteína de unión al ADN cuando el sitio de unión aún no se conoce aprovecha los métodos químicos para sintetizar grandes conjuntos de fragmentos cortos de ADN de secuencia aleatoria. La proteína de interés se mezcla con la población de moléculas de ADN, y los ADN a los que se une se recuperan y secuencian. Una comparación de las secuencias unidas revela fácilmente el consenso, porque cada uno de los fragmentos es muy corto. Este último método (a menudo llamado SELEX) se usa ampliamente para definir sitios de unión para proteínas de unión a ADN previamente no caracterizadas. SELEX se describe con más detalle en el Capítulo 7.
CAJA 13-1 FIGURA 1 Secuencia de consenso del promotor y consenso de espaciamiento. (Redibujado, con autorización, de Alberts B. et al.2002. Biología molecular de la célula, Cuarto
T T GRAMO A C A T A T A A T
nucleótidos
- 10 ed., pág. 308, figura 6.12. # Garland Science / Taylor & Francis Books LLC.)
isomerización, y con qué facilidad puede escapar la polimerasa. La correlación entre
la fuerza del promotor y la secuencia explica por qué los promotores son tan
heterogéneos: algunos genes necesitan expresarse más que otros, y es probable
que los primeros tengan secuencias más cercanas al consenso.
Un elemento de ADN adicional que se une a la ARN polimerasa se encuentra en
algunos promotores fuertes, por ejemplo, los que dirigen la expresión de los genes
del ARN ribosómico (ARNr). Esto se llamaElemento UP (véase la figura 13-5b) y
aumenta la unión de la polimerasa al proporcionar una interacción específica
adicional entre la enzima y el ADN.
Otra clase de s 70 los promotores carecen de una región -35 y, en cambio, tienen un elemento llamado "extendido -10" (véase la figura 13-5c). Esto comprende una región estándar -10 con un elemento de secuencia corta adicional en su sentido ascendente
frecuencia de nucleótidos (%)
438 Capítulo 13
FIGURA 13-7 sya las subunidades reclutan la enzima del núcleo de la ARN polimerasa hacia el promotor. El dominio carboxi-terminal del a subunidadaCTD) reconoce el UPelement (si está presente), mientras que s las regiones 2 y 4 reconocen las regiones –10 y –35, respectivamente (véase la figura 13-6). En esta figura, la ARN polimerasa se muestra en una representación esquemática bastante diferente a la presentada en figuras anteriores. Esta representación es particularmente útil para indicar superficies que tocan el ADN y las proteínas reguladoras, y la usamos nuevamente en algunas figuras del capítulo 18 cuando consideramos la regulación de la transcripción en bacterias.
αNTD
αCTD (^) σ4 σ^ σ
Elemento UP - 35 - 10
subunidad, llamada aCTD (Figura 13-7). losaCTD está conectado al aNTD por un
enlazador flexible. Por tanto, aunque elaNTD está incrustado en el cuerpo de la
enzima, la aCTD puede alcanzar el elemento aguas arriba y puede hacerlo incluso
cuando ese elemento no se encuentra inmediatamente adyacente a la región -35,
sino más arriba.
los s La subunidad se coloca dentro de la estructura de la holoenzima de tal
manera que hace factible el reconocimiento de varios elementos promotores.
Por lo tanto, las regiones de unión al ADN apuntan lejos del cuerpo de la
enzima, en lugar de estar incrustadas. Además, el espacio entre esas regiones
es consistente con la distancia entre los elementos de ADN que reconocen:
s las regiones 2 y 4 están separadas por 75 Å cuando s está unido a la holoenzima,
y esta es aproximadamente la misma distancia que la que hay entre los centros de
los elementos –10 y –35 de una s 70 promotor (véase la figura 13-7). Este
espaciamiento bastante grande de los dominios proteicos es acomodado por la
región entres regiones 2 y 4, es decir, por región 3, especialmente región
3.2, también llamado s3/4 enlazador (véanse las figuras 13-4 y 13-6).
La transición al complejo abierto implica cambios estructurales
en la ARN polimerasa y en el ADN promotor
La unión inicial de la ARN polimerasa al ADN promotor en el complejo
cerrado deja el ADN en forma bicatenaria. La siguiente etapa en la
iniciación requiere que la enzima se involucre más íntimamente con el
promotor, en el complejo abierto. La transición del complejo cerrado al
abierto implica cambios estructurales en la enzima y la apertura de la
doble hélice del ADN para revelar las hebras plantilla y no plantilla. Este
"derretimiento" ocurre entre las posiciones –11 yþ2, con respecto al sitio
de inicio de la transcripción.
En el caso de la enzima bacteriana que llevas 70 , esta transición, a menudo llamada isomerización, no requiere energía derivada de la hidrólisis de ATP y, en cambio, es el resultado de un cambio conformacional espontáneo en el complejo de ADN enzimático a una forma más energéticamente favorable. Como señalamos anteriormente, dos bases en la cadena sin plantilla del elemento -10 (A 11 y T 7 ) se saltan de sus interacciones de apilamiento de bases y en su lugar se insertan en los bolsillos dentro
los s proteína donde hacen interacciones más favorables. Al estabilizar la forma
monocatenaria del elemento –10, estas interacciones impulsan la fusión de la
región promotora (v. Fig. 13-8).
La isomerización es esencialmente irreversible y, una vez completa, generalmente garantiza que la transcripción se iniciará posteriormente (aunque la regulación aún puede imponerse después de este punto en algunos casos). La formación del complejo cerrado, por el contrario, es fácilmente reversible: la polimerasa puede disociarse del promotor tan fácilmente como hacer la transición al complejo abierto.
Para visualizar los cambios estructurales globales dentro de la polimerasa
que acompañan a la isomerización, necesitamos examinar la estructura de la
holoenzima con más detalle. Un canal corre entre las tenazas de la garra
Mecanismos de transcripción 439
a B
RPC
5 ' –12 - 11^ - 10^ - 9^ - 8^ - 7^ 3 '
T A T A A T
A T A T T A
σ 4 σ 2 3 '^ 5 '
- 10 elementos
- 35 elemento dsDNA ssDNA
- 10 elemento
5 ' –12 A^ - 10^ - 9^ - 8 T
σ 2 - 11^ - 7^ 3 '
5 ' –12 - 11^ - 10^ - 9^ - 8^ - 7^ 3 '^ T^ T^ A^ A
T –
T A T A A T
A (^) T A T A T T A 3 '^ A 3 ' 5 ' (^) T T A –11 ningún promotor se derrite sin transcripción
A 5 ' promotor de fusión transcripción T–
A–
FIGURA 13-8 Reconocimiento y fusión de los elementos –10 por s región 2. s la región 2 tiene dos bolsillos que unen cada uno una base volteada de la hebra sin plantilla del elemento –10. Estas reacciones de unión energéticamente preferidas impulsan la fusión del promotor y, por tanto, la transición del complejo cerrado al abierto, sin necesidad de hidrólisis de ATP. (a) Los magnificados vista de s 2 de Taq RNA polimerasa, con el dsDNA del complejo cerrado (azul) y la porción de ssDNA del complejo abierto (amarillo). El corte revela las dos bases volteadas, A y T (amarillo), en los bolsillos de encuadernación. Las flechas rojas muestran cómo las bases invertidas se relacionan con los mismos nucleótidos en el complejo cerrado. (Imagen reproducida, con autorización, de Feklistov A. y Darst SA 2011. Celda 147: 1257, figura 6C, pág. 1265. # Elsevier.) (B) La imagen mostrada en el panel a se representa aquí esquemáticamente para mostrar más claramente la interacción entres región 2 y la hebra sin plantilla de la 210, específicamente las bases volteadas que impulsan la fusión del ADN en esta región. (Adaptado, con autorización, de Liu X., Bushnell DA y Kornberg RD 2011.Celda 147: 1218, figura 1, pág. 1219. # Elsevier.)
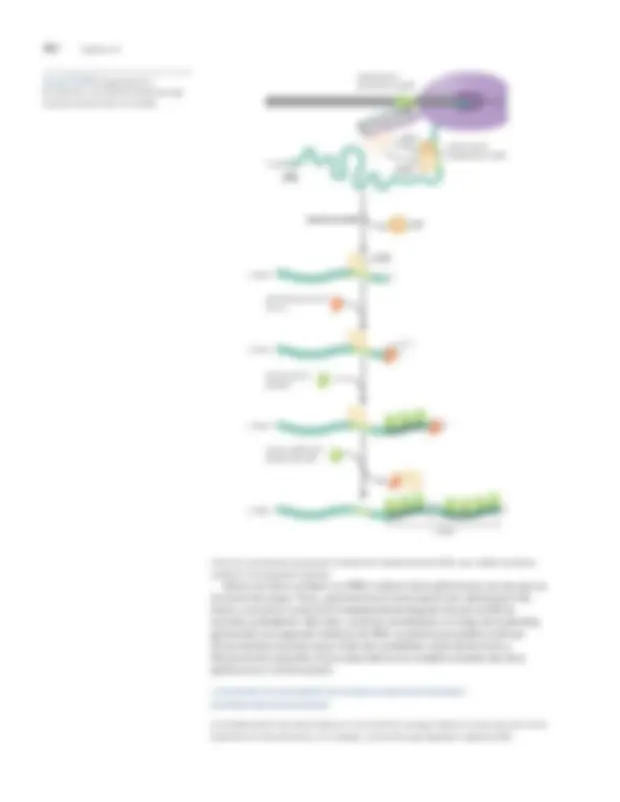
enzima, como se describió anteriormente (véase la figura 13-2). El sitio activo de la enzima, que está formado por regiones tanto de laB y B 0 subunidades, se encuentra en la base de las tenazas dentro de la hendidura del centro activo. Hay cinco canales en la enzima, como se muestra en la ilustración del complejo abierto en la figura 13-9. El canal de captación de NTP (no se muestra en la figura; ver el título de la figura) permite que los ribonucleótidos ingresen al centro activo. El canal de salida de ARN permite que la cadena de ARN en crecimiento abandone la enzima a medida que se sintetiza durante el alargamiento. Los tres canales restantes permiten la entrada y salida del ADN de la enzima, de la siguiente manera.
El ADN corriente abajo (es decir, el ADN delante de la enzima, aún por
transcribir) entra en la hendidura del centro activo en forma de doble hebra a
través del canal de ADN corriente abajo (entre las pinzas). Dentro del centro activo
© 2011 Elsevier
© 2011 Elsevier
Mecanismos de transcripción 441
ataque químico al NTP entrante. El requisito de tales interacciones específicas entre
la enzima y el nucleótido iniciador probablemente explica por qué la mayoría de las
transcripciones comienzan con el mismo nucleótido. La estructura del complejo
abierto muestra que els El enlazador de la región 3/4 interactúa con la hebra
plantilla, organizándola en la conformación y ubicación correctas para permitir la
iniciación. De acuerdo con esto, en experimentos que utilizan una ARN polimerasa
que contiene uns 70 derivado que carece de esta parte de s, la iniciación requiere
concentraciones mucho más altas de lo normal de uno o ambos de los dos
primeros ribonucleótidos.
Durante la transcripción inicial, la ARN polimerasa permanece estacionaria y
atrae el ADN corriente abajo hacia sí mismo
Como hemos descrito, durante la transcripción inicial, la ARN polimerasa produce y
libera transcripciones cortas de ARN de 10 nucleótidos (síntesis abortiva) antes de
escapar del promotor, entrar en la fase de alargamiento y sintetizar la transcripción
adecuada. Durante mucho tiempo no estaba claro cómo el sitio activo de la enzima
se transloca a lo largo de la plantilla de ADN durante los ciclos iniciales abortivos de
transcripción. Se propusieron tres modelos generales (como se muestra en la figura
13-10 y se describe a continuación).
1. El modelo de "excursión transitoria" propone ciclos transitorios de
translocación directa e inversa de la ARN polimerasa. Por tanto, se cree que
la polimerasa abandona el promotor y se transloca un poco a lo largo de la
plantilla de ADN, sintetizando una transcripción corta antes de abortar la
transcripción, liberando la transcripción y volviendo a su ubicación original
en el promotor.
- "Inchworming" invoca un elemento flexible dentro de la polimerasa que permite que un módulo en la parte frontal de la enzima, que contiene el sitio activo, se mueva hacia abajo, sintetizando una transcripción corta antes de abortar y retraerse al cuerpo de la enzima que aún se encuentra en el promotor..
3. "Scrunching" propone que el ADN aguas abajo de la polimerasa estacionaria,
unida al promotor, se desenrolla y se introduce en la enzima. FIGURA 13-10 Mecanismo de transcripción inicial.
Durante la transcripción inicial, el centro activo de la ARN polimerasa se transloca hacia adelante en relación con la plantilla de ADN y sintetiza transcripciones cortas antes de abortar, luego repite este ciclo hasta que escapa del promotor. Se han propuesto tres modelos para dar cuenta de esto y se muestran en la figura. Según el primero de ellos, excursiones transitorias (que se muestran en la parte superior), la polimerasa se mueve a lo largo del ADN. En la segunda, una pulgada (que se muestra en el medio), la parte frontal de la enzima se mueve a lo largo del ADN, pero debido a una región flexible dentro de la enzima, la parte posterior de la enzima puede permanecer estacionaria en el promotor. En el tercer modelo de crujido (que se muestra en la parte inferior), la enzima permanece estacionaria y atrae el ADN hacia sí misma. Las diferencias entre estos modelos se explican en el texto, al igual que la evidencia que respalda el aplastamiento como la verdadera imagen de lo que sucede. (Modificado con autorización de Kapanidis AN et al.
- Ciencias 314: 1144-1147, figura 1a.
AAAS.)
"Excursiones transitorias" RNAP (NTP)norte (PÁGINASI)norte
ARN abortivo
"Pulgada" (NTP)norte (PÁGINASI)norte
ARN abortivo
"Aplastando" (NTP)norte (PÁGINASI)norte
ARN abortivo - 35 –10 +1^ © 2006 AAAS
442 Capítulo 13
El ADN así acumulado dentro de la enzima se aloja como
protuberancias monocatenarias.
Ahora se cree que el tercer modelo:estrujarrefleja lo que realmente sucede.
Esta conclusión se basa en una serie de hallazgos, incluidos experimentos que
utilizan análisis de una sola molécula que permiten medir las posiciones de
diferentes partes de la polimerasa en relación entre sí y con el ADN molde
durante la transcripción inicial. Estos experimentos muestran que durante la
transcripción inicial, la polimerasa permanece estacionaria en el promotor,
desenrolla el ADN corriente abajo y atrae ese ADN hacia sí mismo. Solo el
modelo de aplastamiento es consistente con estos resultados.
El escape del promotor implica romper las interacciones entre la polimerasa y el promotor y el núcleo de la polimerasas Interacciones
Como hemos visto, durante la transcripción inicial, tiene lugar el proceso de
iniciación abortiva y se generan y liberan transcripciones cortas (de 9
nucleótidos o menos). La polimerasa logra escapar del promotor y entrar en la
fase de elongación solo una vez que ha logrado sintetizar una transcripción de
una longitud umbral de 10 o más nucleótidos. Una vez que esta longitud, la
transcripción no puede acomodarse dentro de la región donde se hibrida con
el ADN y debe comenzar a enhebrarse en el canal de salida del ARN (fig. 13-9).
El escape del promotor se asocia con la ruptura de todas las interacciones
entre la polimerasa y los elementos del promotor y entre la polimerasa y
cualquier proteína reguladora que opere en el promotor dado (capítulo 18).
No está claro por qué la ARN polimerasa debe pasar por este período de iniciación abortiva antes de lograr el escape, pero una vez más una región de la s El factor parece estar involucrado, actuando como un imitador molecular. En este caso, es el enlazador de la región 3/4 e imita el ARN. Esta región des se encuentra en el medio del canal de salida de ARN en el complejo abierto (véase la figura 13-9), y para una cadena de ARN que puede ser mayor de 10 nucleótidos, esta región desdebe ser expulsado desde ese lugar, un proceso que puede tomar la enzima varios intentos.
La expulsión del s el enlazador de región 3/4 probablemente representa s estar
más débilmente asociado con la enzima alargadora que con el complejo abierto; de
hecho, a menudo se pierde por completo del complejo de alargamiento.
El aplastamiento se invierte al escapar: el ADN desenrollado durante el
aplastamiento se rebobina, con el colapso concomitante de la burbuja de
transcripción de un tamaño de 22 a 24 nucleótidos a 12-14 nucleótidos (fig. 13-3). Se
cree que este proceso proporciona la energía requerida por la polimerasa para
romper la polimerasa (promotor y núcleo)s interacciones asociadas con el escape.
Por lo tanto, el aplastamiento está lejos para almacenar y movilizar energía durante
el inicio de la transcripción, y su liberación al escapar es lo que permite que la
polimerasa se libere del promotor y se desprenda.s factor del núcleo.
En el Cuadro 13-2, Las ARN polimerasas de una sola subunidad, vemos cómo estas ARN polimerasas simples, a pesar de carecer de un s subunidad, experimentan un cambio estructuralmente comparable en la transición del complejo de iniciación al de alargamiento.
La polimerasa de alargamiento es una máquina
procesadora que sintetiza y corrige el ARN
El ADN pasa a través de la enzima alargadora de una manera muy similar a su paso
a través del complejo abierto (fig. 13-9). Por lo tanto, el ADN de doble hebra ingresa
al frente de la enzima entre las pinzas. En la apertura de la hendidura catalítica, las
hebras se separan para seguir diferentes caminos a través de la enzima antes de
salir por sus respectivos canales y volver a formar un
444 Capítulo 13
Polimerasa de ARN ADN
ARN
a 0 (no trasladado) C +1 (translocado hacia adelante, enlazado a NTP)
B +1 (translocado hacia adelante) D -1 (translocado inverso)
FIGURA 13-11 Plantilla y transcripción dentro del complejo alargador de ARN polimerasa.
La figura muestra diagramas esquemáticos de las posiciones relativas del ARN y la plantilla de ADN dentro de la ARN polimerasa en varios estados del proceso de transcripción. (a) La polimerasa no translocada (0) muestra la cadena de ARN emparejada con la cadena de ADN de la plantilla para una recta base 9. (b) Polimerasa translocada hacia adelante ( þ1) muestra la situación en la que la enzima se ha trasladado una base hacia adelante. (c) La polimerasa translocada hacia adelante con límites NTP muestra el ADN y el ARN en la misma posición que enb con el límite NTP entrante. (d) La polimerasa de translocación inversa (–1) muestra la situación cuando la enzima se transloca hacia atrás en una base como lo hace durante la edición hidrolítica. (Redarrow) Una posición dentro de la polimerasa, la misma en todas las partes de la figura. Consulte el texto para obtener más detalles. Para mayor claridad, la polimerasa mostrada no tiene una orientación diferente a la de la Figura 13-9, con el canal de salida de ARN hacia abajo. (Figuras basadas en imágenes cortesía de Richard Ebright.)
a lo largo del alargamiento: a medida que se separa 1 pb por delante de la enzima de procesamiento, se forma 1 pb detrás de ella. Además de sintetizar la transcripción, la ARN polimerasa realiza dos funciones de corrección de pruebas en esa transcripción en crecimiento. El primero de estos se llama edición pirofosforolítica. En esto, la enzima utiliza su sitio activo, en una simple reacción inversa, para catalizar la eliminación de un ribonucleótido incorrectamente insertado, mediante la reincorporación de PPi. La enzima puede incorporar otro ribonucleótido en su lugar en la cadena de ARN en crecimiento. Tenga en cuenta que la enzima puede eliminar las bases correctas o incorrectas de esta manera, pero pasa más tiempo sobre los desajustes que las coincidencias y, por lo tanto, elimina las primeras con más frecuencia. En el segundo mecanismo de corrección de pruebas, llamadoedición hidrolítica,
la polimerasa retrocede en uno o más nucleótidos (véase la figura 13-11d) y escinde
el producto de ARN, eliminando la secuencia que contiene el error.
Mecanismos de transcripción 445
La edición hidrolítica es estimulada por Grefactores, que mejoran la función de edición hidrolítica y sirven como factores estimulantes de la elongación; es decir, aseguran que la polimerasa se alargue de manera eficiente y ayuden a superar la "detención" en secuencias que son difíciles de transcribir. Esta combinación de funciones es comparable a las impuestas a la ARN polimerasa II eucariota por el factor de transcripción TFIIS (véase más adelante en este capítulo y la figura 13-22). Otro grupo de proteínas, las proteínas Nus, se une a la polimerasa en la fase de elongación y promueve, de formas todavía bastante indefinidas, los procesos de elongación y terminación (para ejemplos de regulación durante el alargamiento, véase el capítulo 18). Una de las proteínas Nus bacterianas, NusG, está muy conservada también en Archaea y Eucariotas (donde se llama Spt5; ver discusión posterior).
La ARN polimerasa puede detenerse y debe eliminarse
En determinadas circunstancias, una ARN polimerasa que se alarga puede
detenerse y dejar de transcribir. Una causa común de detención es una hebra de
ADN dañada. Las consecuencias de la detención pueden ser catastróficas si el gen
que se transcribe es esencial, ya que la polimerasa detenida no producirá ningún
producto, y esa misma enzima obstaculizará el paso de otras polimerasas que
intenten transcribir el mismo gen.
Para hacer frente a esta situación, la célula tiene una maquinaria que elimina la polimerasa detenida y al mismo tiempo recluta enzimas reparadoras (en particular, la endonucleasa Uvr (A) BC); la reparación que sigue se llama reparación acoplada a la transcripción, que analizamos en el capítulo 10. Tanto la eliminación de la polimerasa como el reclutamiento de la enzima reparadora se llevan a cabo mediante una única proteína llamada TRCF.
TRCF tiene actividad ATPasa. Se une al ADN de doble hebra aguas arriba de
la polimerasa y utiliza el motor ATPasa para translocar a lo largo del ADN hasta
que encuentra la ARN polimerasa estancada. La colisión empuja a la
polimerasa hacia adelante, ya sea permitiéndole reiniciar el alargamiento o,
más a menudo, provocando la disociación del complejo ternario de ARN
polimerasa, ADN molde y transcrito de ARN. Esto termina la transcripción de
esa enzima, pero deja paso a las enzimas reparadoras y a otra ARN polimerasa.
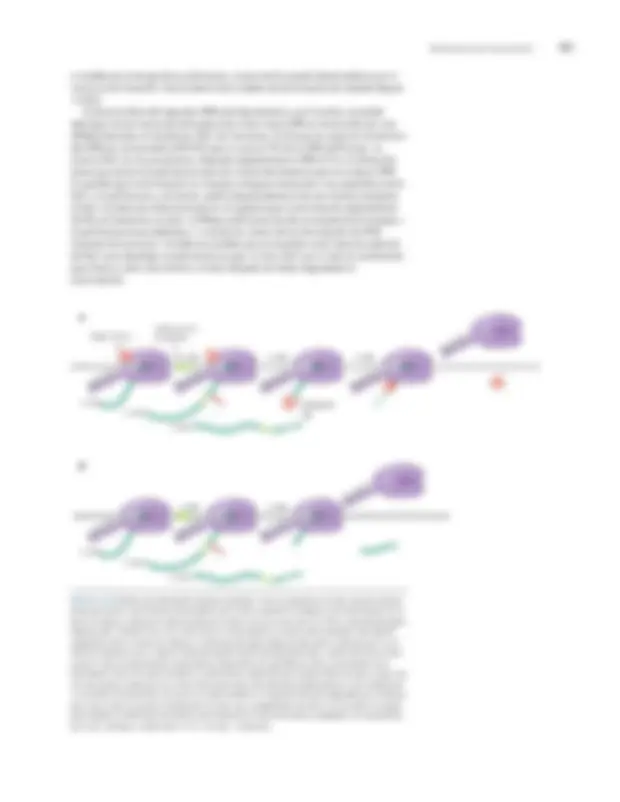
La transcripción se termina por señales dentro de la secuencia de ARN
Ya hemos visto una forma en la que se puede terminar la transcripción. Cuando la ARN polimerasa se detiene durante el alargamiento, puede desprenderse del ADN mediante la acción del translocador TRCF (discutido anteriormente). Esta terminación es provocada por el ADN dañado o por otros obstáculos imprevistos. Pero la terminación es una función normal e importante en los extremos de los genes. Allí, secuencias llamadas terminadores desencadenar la polimerasa en alargamiento para que se disocie del ADN y libere la cadena de ARN que ha creado. En las bacterias, los terminadores vienen en dos tipos:Dependiente de Rho y Rho-independiente. El primero, como su nombre indica, requiere una proteína llamada Rho para inducir la terminación. El segundo provoca la terminación sin la participación de otros factores. Nos ocupamos de cada tipo de terminador por turno. Los terminadores dependientes de Rho tienen elementos de ARN bastante mal definidos llamados rodera sitios (discutidos más adelante), y para que funcionen requiere la acción del factor Rho. Rho, que es una proteína en forma de anillo con seis subunidades idénticas, se une al ARN monocatenario cuando sale de la polimerasa (fig. 13-12). La proteína también tiene actividad anATPasa y, una vez unida a la transcripción, Rho utiliza la energía derivada de la hidrólisis de ATP para inducir la terminación. Queda por determinar el mecanismo preciso de terminación, y los modelos incluyen lo siguiente: Rho empuja la polimerasa hacia adelante en relación con el ADN y el ARN, lo que da como resultado la terminación de una manera análoga a la terminación por
Mecanismos de transcripción 447
pareja simetría FIGURA^ 13-13^ Secuencia de un terminador independiente de Rho. En la parte superior está la secuencia, en el ADN, del terminador. A continuación se muestra la secuencia del ARN y la imagen inferior muestra la estructura de la horquilla del terminador. El terminador en cuestión es deltrpatenuador, discutido en el Capítulo 18. Los recuadros muestran mutaciones aisladas en la secuencia que interrumpen el terminador. (Adaptado de Yanofsky C. 1981. Naturaleza 289: 751–758.)
CCCAGCCCGCCTAATGAGCGG GCTTTTTTTTGAACAAAAGGG
ADN TCGGGCGGATTACTCGCCCGA AAAAAAACTTGTTTT
5 ' 3 '
ARN 5 ' CCCAGCCCGCCUAAUGAGCGGGCUUUUUUUU 3 '
transcripción doblada para formar una horquilla de terminación
A A U U C
GRAMO A C GRAMO A, U GRAMO C A U
CG CG CG
A A, U, C A, U GRAMO (^) C 5 ' CCC A UUUUUUUU 3 '
GRAMO supresión
La horquilla funciona como un terminador eficiente sólo cuando va seguida de un tramo de pares de bases A: U, como hemos descrito. Esto se debe a que, en esas circunstancias, en el momento en que se forma la horquilla, la cadena de ARN en crecimiento se mantendrá en la plantilla en el sitio activo solo por pares de bases A: U. Debido a que los pares de bases A: U son los más débiles de todos los pares de bases (más débiles incluso que los pares de bases A: T), se interrumpen más fácilmente por los efectos del ciclo del tallo en la polimerasa de transcripción y, por lo tanto, el ARN se disociará más fácilmente (fig. 13-14).
5' 3'^ UUUUUUU
FIGURA 13-14 Terminación de la transcripción. Un modelo de cómo podría funcionar el terminador independiente de Rho. (Arriba) La horquilla se forma en el ARN (fig. 13-11) tan pronto como esa región ha sido transcrita por la polimerasa (la enzima no se muestra aquí). (Medio) Esa estructura de ARN interrumpe la polimerasa justo cuando la enzima transcribe el tramo de ADN rico en AT aguas abajo. (Abajo) No está claro exactamente cómo la horquilla interrumpe la transcripción de la polimerasa (consulte el texto para ver modelos alternativos), pero las interacciones débiles entre la transcripción y el ADN de la plantilla (Us en la transcripción y As en la plantilla) parecen dar lugar a esa liberación. transcripción más fácil. (Adaptado de Platt T. 1981.Celda 24: 10-23.)
5' 3' UUUUUU^ U
UUUUUUU 5 ' 3 '
448 Capítulo 13
TRANSCRIPCIÓN EN EUCARIOTAS
Como ya hemos comentado, la transcripción en eucariotas se realiza mediante polimerasas estrechamente relacionadas con las ARN polimerasas que se encuentran en procariotas. Esto no es de extrañar, ya que el proceso de transcripción en sí es idéntico en los dos casos. Sin embargo, existen diferencias en la maquinaria utilizada en cada caso, una de las cuales ya hemos visto: mientras que las bacterias tienen solo una ARN polimerasa, todos los eucariotas tienen al menos tres diferentes (Pol I, II y III; y las plantas también tienen una Pol IV y una Pol V). Además, mientras que las bacterias requieren solo un factor de iniciación adicional (s), Se requieren varios factores de iniciación para una iniciación eficaz y específica del promotor en eucariotas. Estos se llamanfactores de transcripción general (GTF).
In vitro, los factores de transcripción generales son todo lo que se requieren,
junto con Pol II, para iniciar la transcripción en una plantilla de ADN (sin histonas).
Sin embargo, in vivo, el molde de ADN en células eucariotas se incorpora a
nucleosomas, como hemos visto en el capítulo 8. En estas circunstancias, los
factores de transcripción generales no son suficientes por sí solos para unir
secuencias promotoras y provocar una expresión significativa. Más bien, se
requieren factores adicionales, incluidas las proteínas reguladoras de unión al ADN,
el llamado complejo mediador y, a menudo, enzimas modificadoras de la cromatina.
Primero consideramos el mecanismo básico por el cual Pol II y los factores de transcripción generales se ensamblan en un promotor para iniciar la transcripción in vitro. Luego consideramos las funciones de los componentes adicionales necesarios para promover la transcripción in vivo.
Los promotores del núcleo de la ARN polimerasa II se componen de combinaciones de diferentes clases de elementos de secuencia
El eucariota promotor central se refiere al conjunto mínimo de elementos de secuencia requeridos para el inicio preciso de la transcripción por la maquinaria Pol II, medido in vitro. El promotor Acore tiene típicamente de 40 a 60 nucleótidos de longitud y se extiende hacia arriba o hacia abajo desde el sitio de inicio de la transcripción. La figura 13-15 muestra la ubicación, en relación con el sitio de inicio de la transcripción, de los elementos que se encuentran en los promotores del núcleo Pol II. Estos son el elemento de reconocimiento TFIIB (BRE), el elemento TATA (o caja), el iniciador (Inr) y los elementos impulsores descendentes (conocidos como DPE, DCE y MTE). Normalmente, un promotor incluye algún subconjunto de estos elementos. Así, por ejemplo, los promotores normalmente tienen un elemento TATA o un elemento DPE, no ambos. A menudo, un promotor que contiene TATA también contiene un DCE. El Inr es el
TFIIB TBP TFIID TFIID TFIID TFIID
- 37 - 32 –31 - 26 - 2 + 4 + 6 + 11 + 16 + 21 + 28 +30 +32 + BRE TATA Inr DCEI DCEII DPE DCEIII
GGGCGCCCCCA TATAAAATT CCANTCC CTTCTT ATT CTGT^ Automóvil club británicoGT G CGTG AGC
FIGURA 13-15 Promotor del núcleo Pol II. La figura muestra las posiciones de varios elementos de ADN en relación con el sitio de inicio de la transcripción (indicado por la flecha sobre el ADN). Estos elementos, descritos en el texto, son los siguientes: (BRE) Elemento de reconocimiento TFIIB; (TATA) Caja TATA; (Inr) elemento iniciador; (DPE) elemento promotor corriente abajo; y (DCE) elemento central aguas abajo. Otro elemento, MTE (elemento del motivo diez), descrito en el texto, no se muestra en esta figura, pero está ubicado aguas arriba del DPE. También se muestran las secuencias de consenso para cada elemento (determinadas de la misma manera que se describe para los elementos promotores bacterianos; ver Cuadro 13-1) y (arriba) el nombre del factor de transcripción general que reconoce cada elemento.