¡Descarga Microbiología y más Apuntes en PDF de Microbiología solo en Docsity!
MICROBIOLOGÍA
TEMA 1: DESARROLLO HISTÓRICO DE LA MICROBIOLOGÍA
1. CONCEPTO DE MICROBIOLOGÍA Y MICROORGANIMOS
La microbiología es la rama de la biología que se encarga del estudio de los
microorganismos, su diversidad, evolución e incluso su papel en la naturaleza, en
suelos, aguas, tierras.
Los microorganismos son seres vivos que son tan pequeños que no se pueden ver a
simple vista, solo con el uso de microscopios. Estos microorganismos pueden ser tanto
unicelulares como pluricelulares y además son muy distintos entre ellos tanto
evolutivamente como funcional y estructuralmente, todos ellos solo tienen en común
su pequeño tamaño.
Dentro de los microorganismos se incluyen los virus aunque no tengan estructura
celular.
2. DESARROLLO HISTÓRICO: PRINCIPALES HITOS
2.1. Técnicas microscópicas
En el siglo XVIII Leeuwenhoek consiguió ver los primeros microorganismos, las
bacterias, gracias a un microscopio muy simple con un sola lente.
En el mismo siglo, Hook, otro científico que se dedicaba a la talla de lentes, observó
con más claridad cosas más grandes, pero no llegó al tamaño de Leeuwenhoek.
Con estos dos descubrimientos, se empezó a hacer evidente que todo nuestro
alrededor estaba lleno de
microorganismos de distintos tipos.
Microscopio simple de Leeuwenhoek.
2.2. TÉCNICAS DE CULTIVO DE MICROORGANISMOS
Era evidente pues la existencia de microorganismos gracias a los descubrimientos de
Hook y Leeuwenhoek y al desarrollo de técnicas microscópicas para verlos, sin
embargo hasta el siglo XIX no se desarrollaron técnicas de cultivo de microorganismos
debido a que la microbiología es una ciencia difícil de manejar debido a su pequeño
tamaño.
En la mitad del siglo XIX, se logró hacer el primer cultivo axogénico o puro, el cual de
obtiene aislando un microorganismo y logrando que cree mucha descendencia.
Con esto se consiguió generar suficiente biomasa para estudiar las características de
algunos microorganismos.
Louis Pasteur consiguió, a través de una serie de experimentos consiguió demostrar
que los microorganismos no se crean por generación espontánea, si no que surgían a
través de esporas que se encuentran en todos los sitios.
El cultivo axogénico fue conseguido por Robert Koch, fue uno de los padres de la
microbiología y fundó la escuela alemana.
3. CLASIFICACIÓN Y DIVISIÓN DE LOS MICROORGANISMOS
Los microorganismos son muy diversos, tras su descubrimiento se clasificaban en
pequeños animales y plantas que se podían mover o no, pero con el tiempo se
descubrieron que eran diferentes entre sí y más aún se diferenciaban de los
macroscópicos.
Previamente los microorganismos se clasificaron en grupoel protistas; en la primera
mitad del s. XX los microorganismos también se podían dividir en el tipo de células que
los componían:
EUCARIOTAS YPROCARIOTAS, basándose en esto, Whittaker en 1960 dividió los
organismo en 5 reinos:
Plantae
Animalia
Fungi
Protista
Monera
Con el paso del tiempo , comparando la secuencia de genes muy conservados y viendo
como han ido cambiando en los distintos individuos:
El gen 16S en procariotas y el 18S eucariotas, y comparándolos se obtuvieron los
primeros arboles universales con tres linajes:
Procariotas: Bacterias verdaderas.
Arqueas: Organismo con apariencia bacteriana pero con metabolismo
semejante al de los eucariotas.
Eucariotas: protistas como los hongos, animales y plantas.
El antecesor común, también llamado LUCA, se sitúa en un punto entre bacterias y
arqueas, ya que los eucariotas son mucho más recientes.
También cabe hablar de la importancia de los virus, organismos acelulares muy
utilizados en la biotecnología y en la ingeniería genética.
Microorganismos
4. IMPORTANIA DE LOS MICROORGANISMOS EN LA BIOTECNOLOGÍA
La fermentación se da gracias a levaduras, y gracias a este proceso obtenemos
alimentos como queso, yogures, lácteos, vino y cerveza.
También son las bacterias las encargadas de fabricar sustancias imprescindibles
para la salud humana como la insulina.
Conservantes.
Antibióticos y vacunas.
Organismos transgénicos para abastecer a toda la población.
Biocombustibles.
PCR, eliminación de residuos, sistemas CRISPR-CAS...
TEMA 2
MORFOLOGÍAS
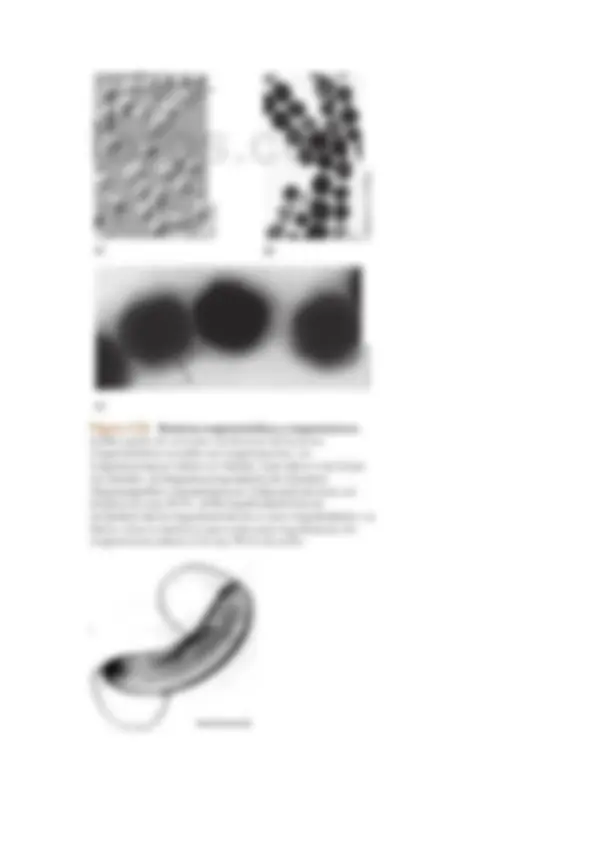
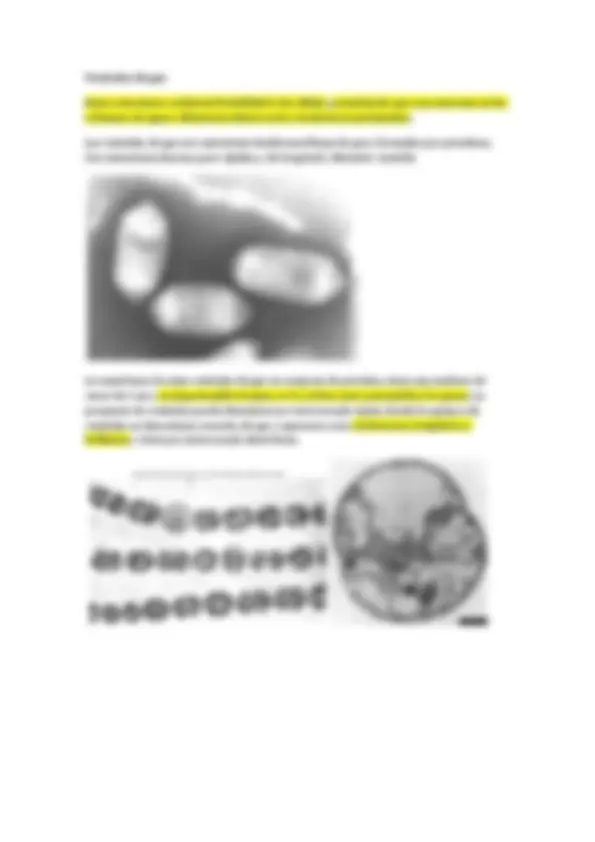
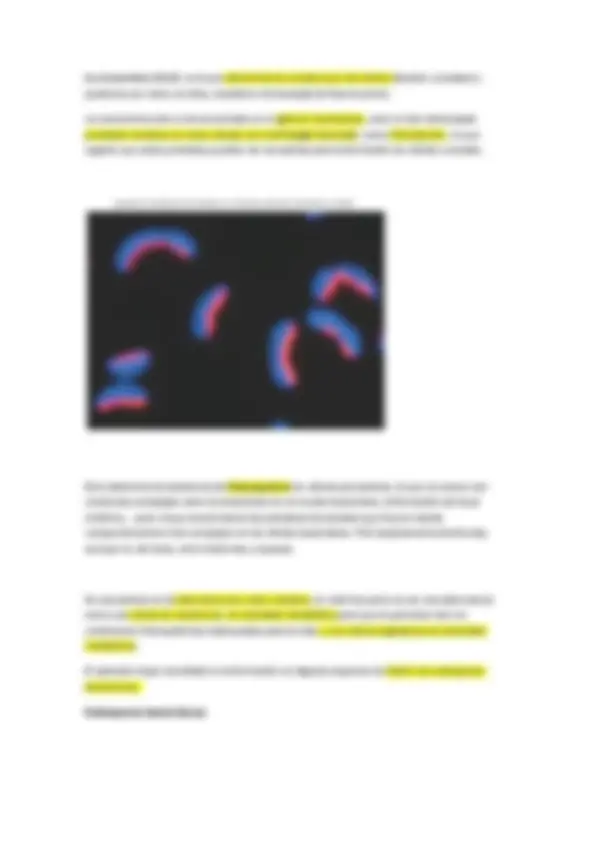
Las células de muchos procariotas se mantienen juntas después de la división celular formando grupos, y estas asociaciones frecuentemente son características de diferentes géneros.
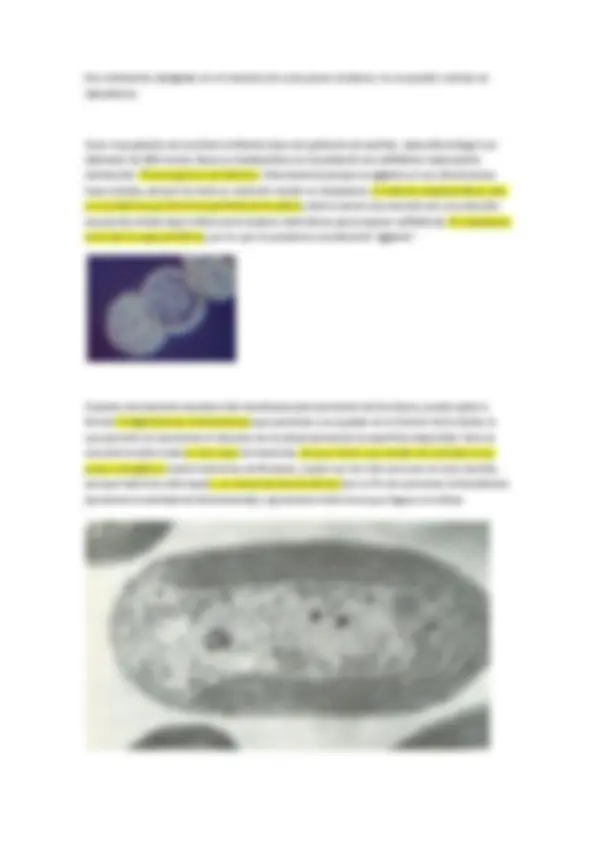
Por ejemplo, algunos cocos forman larcas cadenas (Streptococcus), otros se disponen en agrupaciones cúbicas tridimensionales (Sarcina) y otros forman conjuntos en forma de racimo de uvas (Staphylococcus).
Morfologías curiosas:
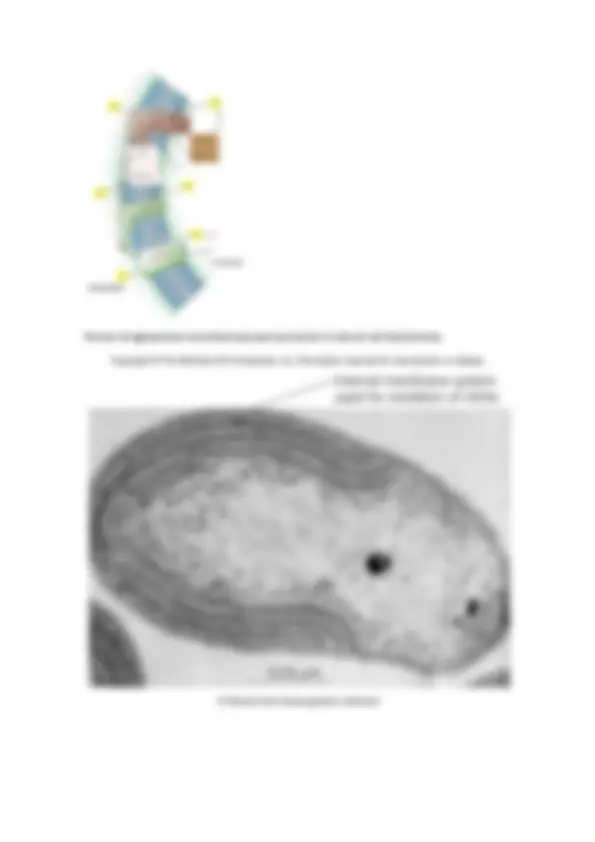
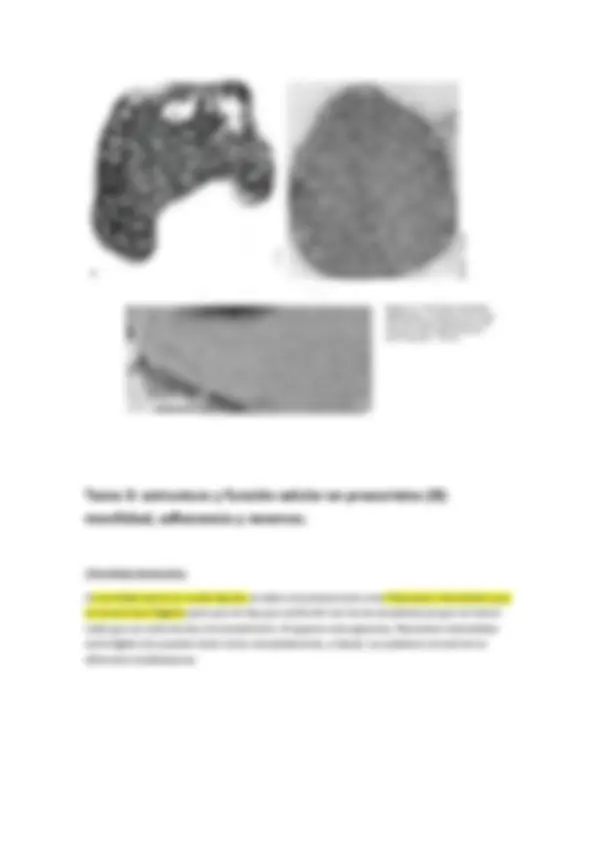
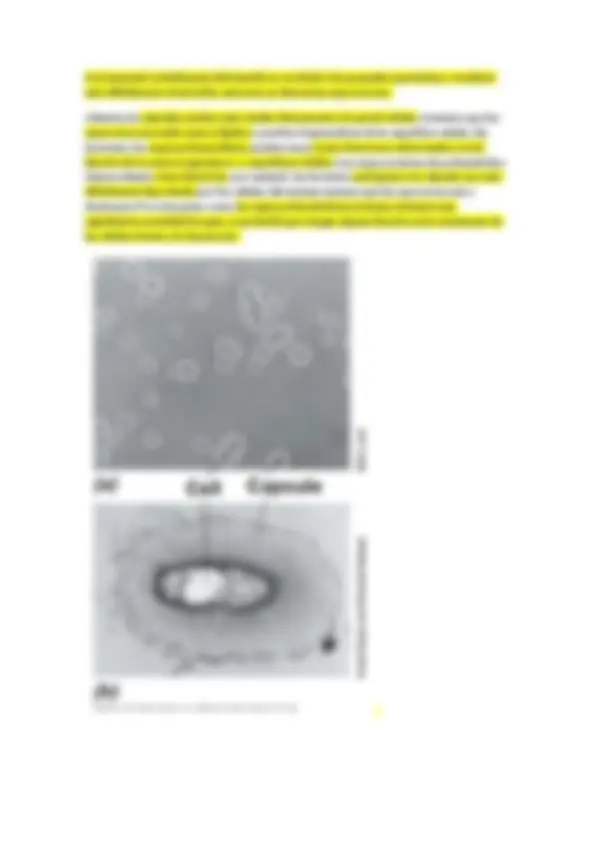
-Células cuadradas (haloarquea)
Solo se dan en arqueas, son perfectamente cuadrados.
En contraste de fases (a) nos permite observar puntos brillantes que son vacuolas gaseosas, que son como flotadores y les permite eso, flotar, en un medio acuático.
En microscopía electrónica (e) de transmisión vemos que las vacuolas gaseosas se situan en la periferia de la célula.
Esta forma cuadrada es muy plana, son como pequeñas laminillas y por esa morfología y lo extraordinariamente fino que son las láminas que forman y las vacuolas de gas, estas bacterias tienden a pegarse en la superficie del agua, que es donde encuentran su hábitat óptimo para desarrollarse.
-Existen más morfologías diferentes a las cuatro principales como
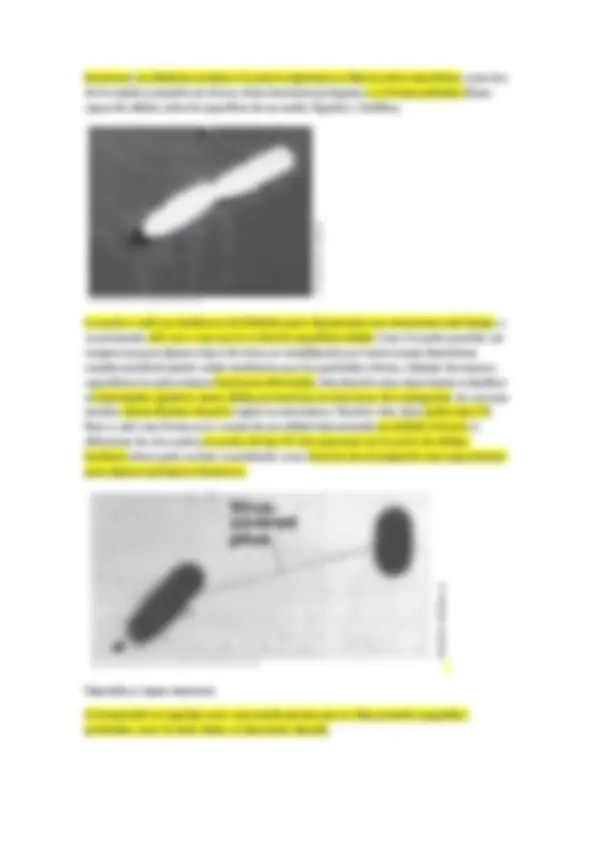
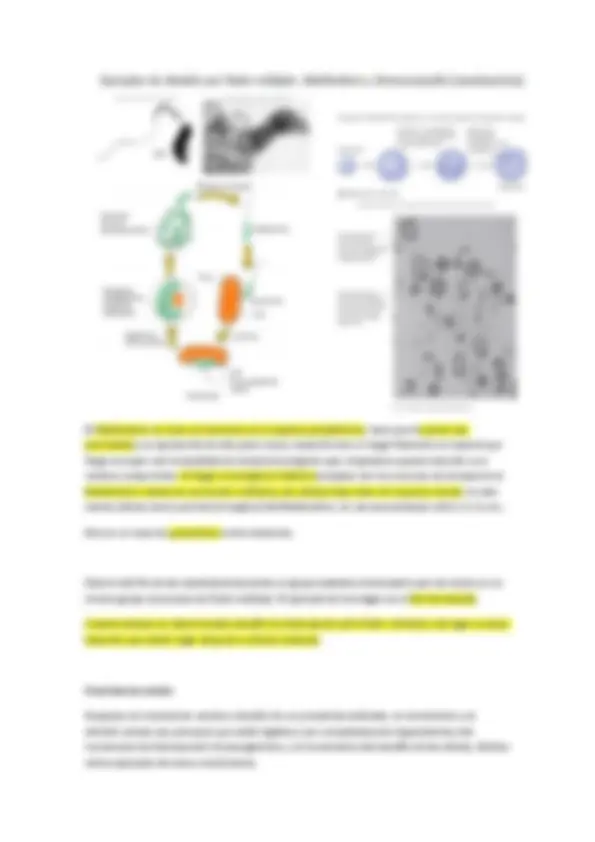
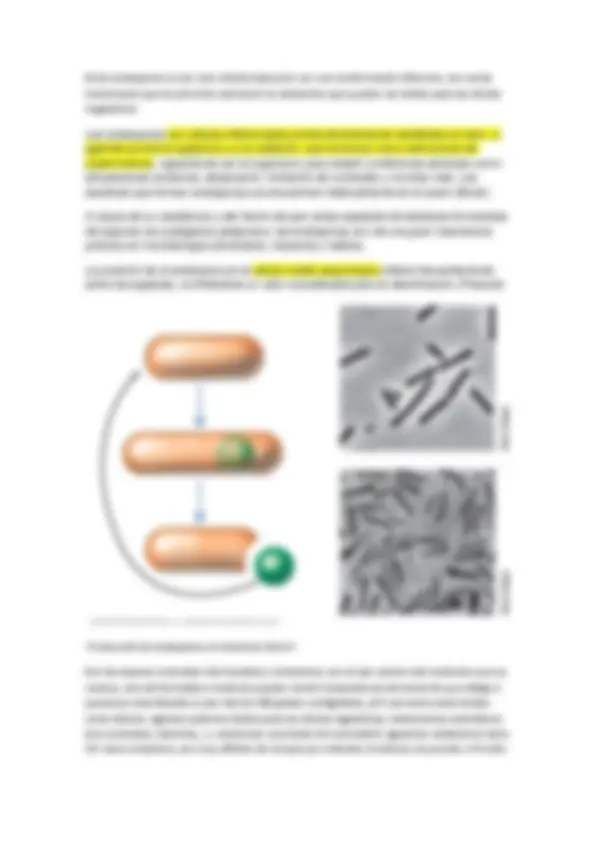
Prostecadas: con varias prostecas, en el cual en un extremo de la prosteca existen yemas para dar a células hijas, tienen sentido de reproducción.
TAMAÑOS
Los procariotas tienen células menos complejas y con arquitecturas más pequeñas, pero esto solo es mayoritario, existen casos a parte.
Cianobacterias, bacterias multicelulares, bastante grandecitas
El que las células procarioticas sean más pequeñas por lo general que las eucariotas tiene la lógica por la relación superficie/volumen. Presencia en las células eucarióticas de compartimentos internos con tareas determinadas y especializadas que ocupan un volumen
Son simbiontes obligados en el intestino de unos peces coralinos, no se pueden cultivar en laboratorio.
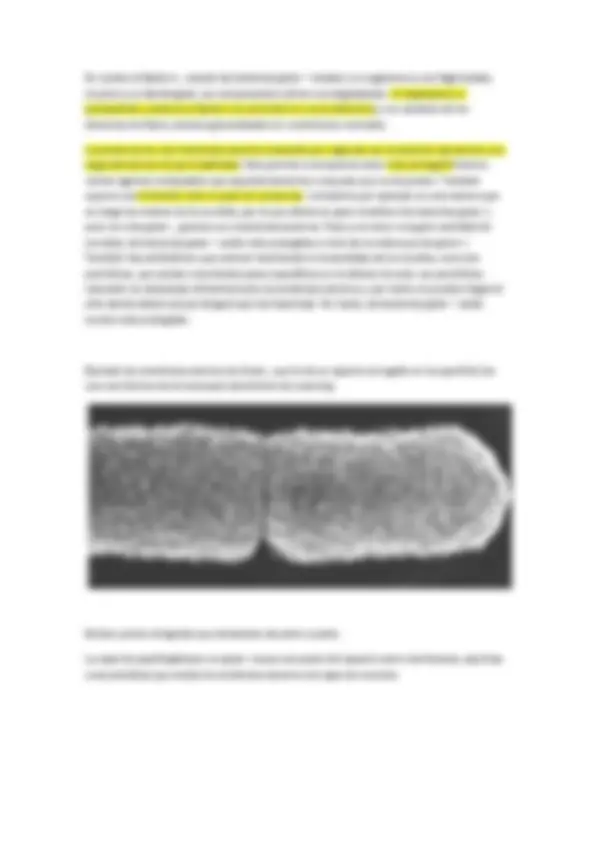
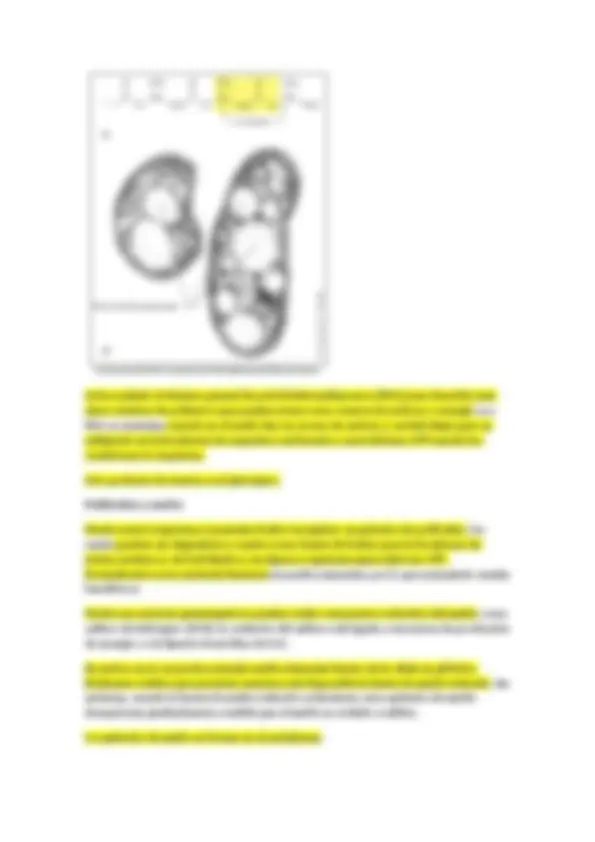
Coco muy grande con puntitos brillantes (que son gránulos de azufre), cada esfera llega a un diámetro de 500 micras. Basa su metabolismo en la oxidación de sulfhídrico hasta azufre elemtental. Thiomargarita namibiensis. Esta bacteria aunque es gigante en sus dimensiones hace trampa, porque no todo su volumen celular es citoplasma, el materia citoplasmático solo es una lámina que forma la periferia de la esfera, dentro existe una vacuola con una solución acuosa de nitrato (que utiliza como aceptor electrónico para respirar sulfhídrico). El citoplasma será solo la capa periférica, por lo que no podemos considerarla “gigante”.
Cuando una bacteria necesita más membrana para aumentar las funciones, puede optar a formar invaginaciones membranosas que penetran y se quedan en el interior de la célula, lo que permite sin aumentar el volumen de la célula aumentar la superficie disponible. Esto se encuentra sobre todo en dos tipos de bacterias, las que tienen que oxidar de sustratos muy pocos energéticos (como bacterias nitrificantes, suelen ser las más comunes en este sentido, aunque habrá de más tipos) y en bacterias fotosintéticas (con el fin de aumentar la fotosíntesis (aumenta la cantidad de fotosistemas) y aprovechar toda la luz que llegue a la célula.
Otra forma de aumentar la superficie disponible es producir prostecas, que suelen ser más largas o más abundantes (número de prostecas característico de cada especie) en medios pobres en un determinado nutriente. Caulobacter y Hypnomicrobium aumentan las prostecas cuando hay poco fosfato, la prosteca es digamos un orgánulo especializado en la captación de fósforo.
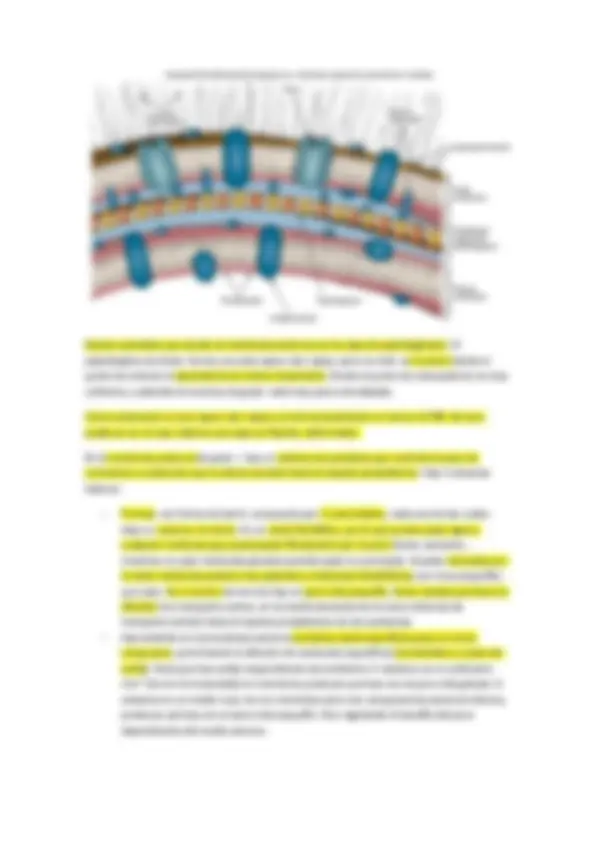
La membrana citoplasmática
Repaso de membrana citoplasmática en bacterias y arqueas (sacado del Brock)
La membrana citoplasmática es una estructura fina que rodea la célula. Esta estructura vital es la barrera que separa el interior celular (el citoplasma) del exterior. Si la membrana ser rompe, se destruye la integridad celular. También es una barrera con elevada permeabilidad selectiva que capacita a la célula para concentrar metabolitos específicos y para excretar productos de desecho.
-Composición
La estructura general de las membranas biológicas es la de una bicapa fosfolipídica. Como se ha indicado anteriormente, los fosfolípidos contienen componentes hidrofóbicos (ácidos grasos) e hidrofílicos (glicerol-fosfato). Los ácidos grasos se orientan hacia el interior formando un ambiente hidrofóbico y las porciones hidrofílicas expuestas a la fase acuosa exterior o al citoplasma.
-Proteínas de membrana:
Las principales proteínas de la membrana citoplasmática tienen zonas hidrofóbicas en las regiones que se incluyen en la membrana y zonas hidrofílicas en las regiones que contactan con el medio exterior y con el citoplasma. La superficie más externa de la membrana citoplasmática se orienta hacia el medio y en algunas bacterias interacciona con diversas proteínas que unen sustratos o procesan grandes moléculas para su transporte a la célula ( proteínas periplasmáticas ). La cara interna de la membrana citoplasmática se orienta al citoplasma y contacta con proteínas implicadas en reacciones que generan energía y otras funciones celulares importantes.
Proteínas integrales de membrana firmemente asociadas a ella, otras solamente tienen una porción anclada en la membrana y presentan regiones fuera de la membrana que se orientan hacia dentro o hacia fuera de la célula. Finalmente, hay otras, llamadas periféricas, que no están en absoluto embebidas en la membrana pero que se asocian fuertemente con sus superficies (por lo general estas proteínas interaccionan con proteínas integrales de membrana que son importantes en procesos celulares relacionados con el metabolismo energético y el transporte).
Bicapas de lípidos en todos los seres vivos de naturaleza celular, que tiene una serie de componentes como fosfolípidos. Acompañada de otros tipos de lípidos, como de tipo esterol, moléculas planas con muchos anillos que dan consistencia a la membrana (como hopanoides).
Las membranas de las arqueas tienen propiedades similares a las de cualquier otro ser vivo en cuanto a permeabilidad, a pesar de su composición química diferente. La permeabilidad de la membrana limita el paso a muy pocas moléculas que pueden atravesarla por difusión pasiva. En cuanto a moléculas cargadas, agua… son iguales a las membranas eucariotas o bacteriales. Las consecuencias de la composición química peculiar de las membranas arqueales son la temperatura y la resistencia. Por ello podemos encontrar arqueas en ambientes extremófilos. Su resistencia a la desagregación por pH extremo o temperaturas tochas es mucho mayor. Las membranas que están compuestas por monocapas son mucho más estables térmicamente hablando que las bicapas. En las arqueas hipertermófilas predomina totalmente la membrana monocapa.
Los lípidos que tienen son siempre los mismos indiferentemente de si son extremófilas o no. Lo que varía es la proporción.
-Existen varias funciones asociadas a las membranas procarióticas
1.- Paso de sustancias
2.- Generación de energía a través de ATPsas (dependientes de H+^ o de Na+)
3.- Anclaje de sistemas de movilidad celular: rotores flagelares, fimbrias, etc …
4.- Biosíntesis de componentes de la membrana y de la pared celular (ocasionalmente de cápsulas y otras estructuras de superficie): proteínas, lípidos, polisacáridos, LPS …
5.- Secreción de proteínas al exterior (sistemas Sec, y otros)
6.- Segregación de copias durante la replicación de plásmidos y cromosomas.
7.- Transducción de señales externas para el control de la expresión génica (sistemas de dos componentes) o la conducta celular (MCPs y quimiotactismo).
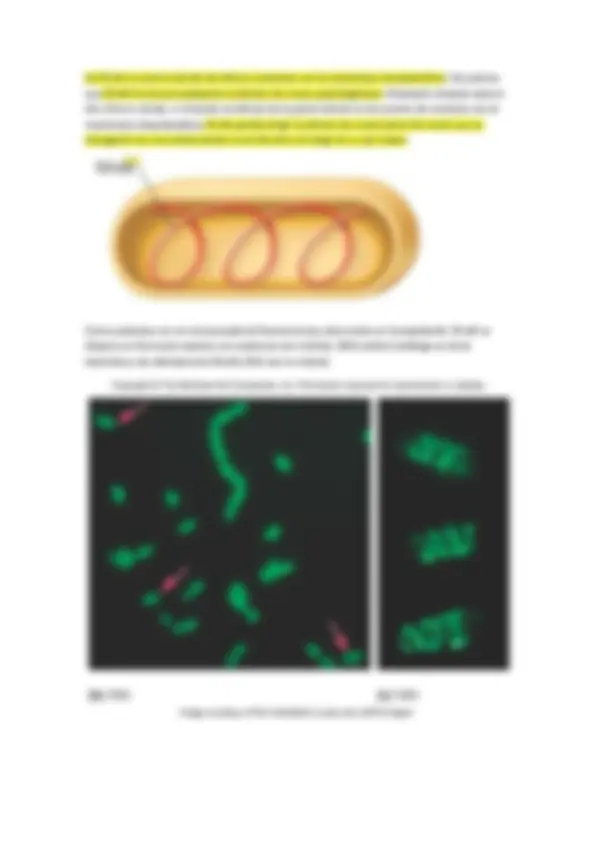
8.- Sede de proteínas implicadas en procesos de mantenimiento de la morfología celular, distribución polarizada de funciones, septación , etc… (FtsZ, MreB, crescentina etc…”citoesqueleto procariótico” ).
La cuestión es que la membrana de procariotas contiene unos 200 tipos diferentes de proteínas en su membrana, y el contenido total (lo que representa la proteína ligada a membrana en procariotas) es de hasta un 70% de su masa. Hay mucha más proteína y mucho más variada en un procariota que en un eucariota. La membrana es el órgano principal, metabólico, de las procariotas. De ahí que la relación superficie-volumen sea tan crítica.
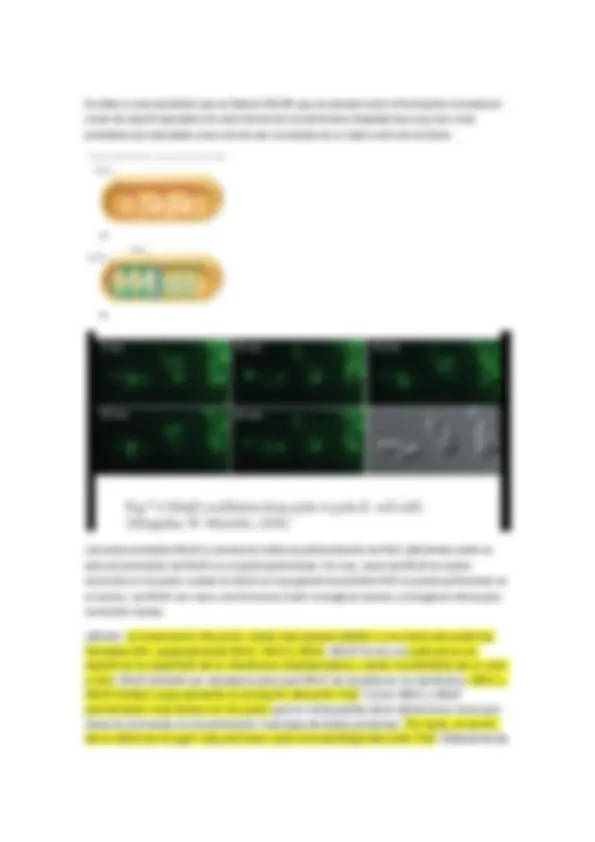
Podemos ver la membrana de Escherichia Coli. LH son los pigmentos membrana, en verde oscuro están las cadenas de transporte… La cuestión es que los fotosistemas también están en la membrana. Según las circunstancias de producción (producir un gradiente de protones a costa de la hidrólisis de ATP o cualquier otra estrategia) la bacteria actuará de una forma u otra.
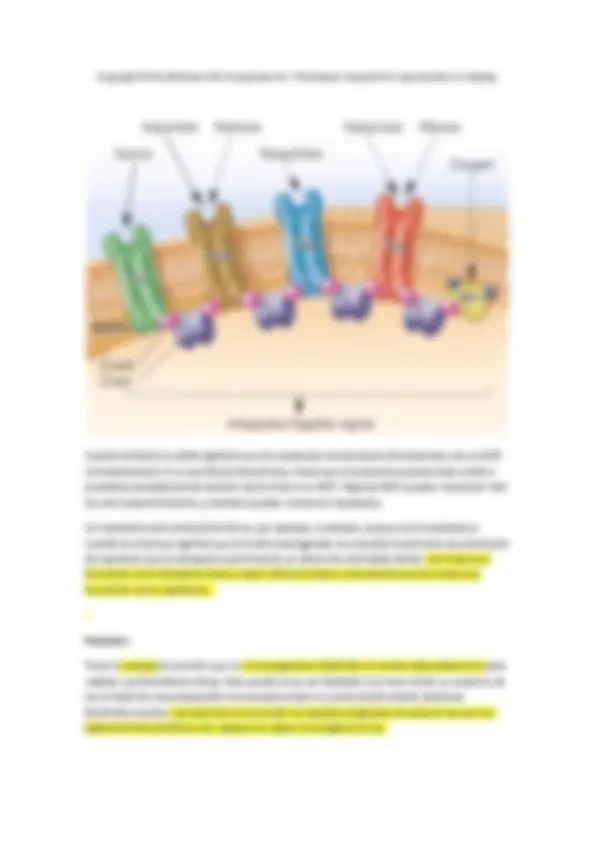
Paso de sustancias. La membrana plasmática como barrera de permeabilidad La mayoría de sustancias no penetran de modo pasivo en la célula y, por lo tanto, deben ser transportadas. Por tanto existe una necesidad de proteínas transportadoras, que a parte de transportar sustancias a través de la membrana son capaces de acumular solutos dentro de la célula contra un gradiente de concentración. Si los solutos únicamente entrasen por difusión jamás se alcanzarían las concentraciones intracelulares necesarias para que las reacciones bioquímicas tengan lugar.
Las funciones de paso a través de membrana se podrían producir por transporte activo (esquemas A y B), o por procesos de translocación de grupo (esquema C).
- Dependientes de gradientes iónicos:
Si la energía que se consume proviene de la disipación de gradiente, entonces hablamos del esquema A, simportadores o antiportadores. También tenemos los sistemas uniportadores. La energía que consumen viene de un gradiente iónico previo. Los transportes antiportes transportan dos sustancias, una de ella a favor de gradiente y la otra en contra (si fuesen las dos en contra no habría energía para el intercambio). En el simporte, tanto la que viaja a favor como la que viaja en contra, van hacia el mismo lado de la membrana. El transporte uniporte solo transporta una molécula cargada, un ión potasio por ejemplo. El paso es en contra de gradiente de concentración, pero va a favor de gradiente de carga (lo que conduce el gradiente es la carga eléctrica).
- Sistemas ABC, dependientes de ATP:
Los sistemas ABC son capaces de hidrolizar ATP, son muy extendidos en el mundo procariótico, puesto que los usan para acumular nutrientes en la célula, y son específicos. En este caso, los sistemas ABC tienen la composición de la imagen B. Una parte del sistema consiste en proteínas que se relacionan con la membrana por la cara citoplásmica, y que son las que se encargan de hidrolizar el ATP, cuya hidrólisis va a servir para el transporte. ABC=ATP Binding Cassette. Además, existe un tercer componente de estos sistemas, una proteína que no está asociada al canal ni la proteína que hidroliza ATP. Se encuentra o bien asociada (laxamente) a la membrana citoplásmica por fuera, o libre por el espacio periplásmico, este tipo de proteína se denomina proteína periplásmica de unión. La proteína periplásmica de unión, no tiene actividad enzimática, solo se une de manera muy afín al sustrato que se va a transportar, aunque se encuentre en concentraciones extremadamente bajas. Una vez que se ha unido el sustrato correspondiente, el complejo interacciona con el respectivo componente transmembranal que transporta el sustrato gracias a la energía liberada por la hidrólisis del ATP.
Si eliminamos estas proteínas de unión al soluto, estos sistemas pierden efectividad TOTALMENTE. Estos sistemas son muy comunes. Podemos encontrar unos 60 genes en el genoma bacteriano que codifican estos sistemas ABC. Por aclarar, las tres partes del sistema ABC son la proteína periplásmica, el transportador (lo que vendría a ser el canal) y el dominio de unión al nucleótido (lo que une e hidroliza el ATP).
Esquema A
Una pequeña proteína llamada HPr, la enzima que la fosforila (enzima I) y la enzima IIa son proteínas citoplasmáticas. Por el contrario, la enzima IIb se sitúa sobre la superficie interior de la membrana y la enzima IIc es una proteína integral de membrana. Tanto HPr como la enzima I son componentes inespecíficos del sistema fototransferasa y participan en el transporte de varios azúcares. En cambio, la enzima II es específica para cada azúcar individual.
La energía requerida por el sistema fosfotransferasa deriva del fosfoenol piruvato, que es un compuesto con alta energía.
Estos sistemas son sistemas que en último término también gastan energía, por lo que rompen un enlace ricoenergético, pero suponen una ventaja muy grande para las células que los usan, y es que el carbohidrato entra y aparece en el citoplasma en forma activada. Para entrar en las vías del metabolismo central (glucolisis o cualquiera), los carbohidratos requieren una fosforilación. Eso a un eucariota le supone gastar un ATP, pero es que en procariota matamos dos pájaros de un tiro. Romper el fosfoenolpiruvato para fosforilar el carbohidrato tiene un gasto neto de un ATP. Usamos un sustrato fermentado que no serviría para gran cosa para activar el carbohidrato.
Las bacterias fermentadoras obligadas obtienen poca energía de los sustratos que fermentan, de manera que usan sistemas fosfotransferasa para captar del medio ATP.
El paso de sustancias a través de la membrana se complica en ciertos grupos de bacterias (gram -), porque no tienen en su superficie una única membrana, sino por dos. Todos los pasos de sustancias se complican, porque de esas dos membranas solo una está energizada (contiene ATPasas, tiene un gradiente iónico a su través…). La membrana externa no está energizada. El paso de sustancias a través de las dos membranas es mucho más complejo. Necesitamos una proteína ligada a la membrana externa, un sistema que conecte con esa proteína y con la membrana citoplasmática, y luego un sistema tipo ABC que transporta el sideróforo del espacio intermembrana al interior.
Esquema C
Tenemos otras funciones residentes en la membrana, el control de la expresión génica que funcionan como sistemas de dos componentes. El componente ligado a la membrana externa, que se fosforila (una quinasa) y una vez se ha fosforilado transmite la señal que se transmite hasta el DNA mediante una segunda proteína que se fosforila a costa de la primera. La primera es la quinasa sensora, que percibe el cambio ambiental. La otra proteína es el regulador de respuesta. Tiene dos estados: uno sin fosforilar y otro fosforilado, y es la que interacciona con la transcripción del material genético promoviéndolo o bloqueándolo según la señal que le venga. Interviene también una fostatasa que elimina un segundo fosfato del regulador de respuesta.
Otro sistema es el sistema que controla el quimiotactismo celular (movimiento de flagelo en función de señales atrayentes o repelentes para la célula).
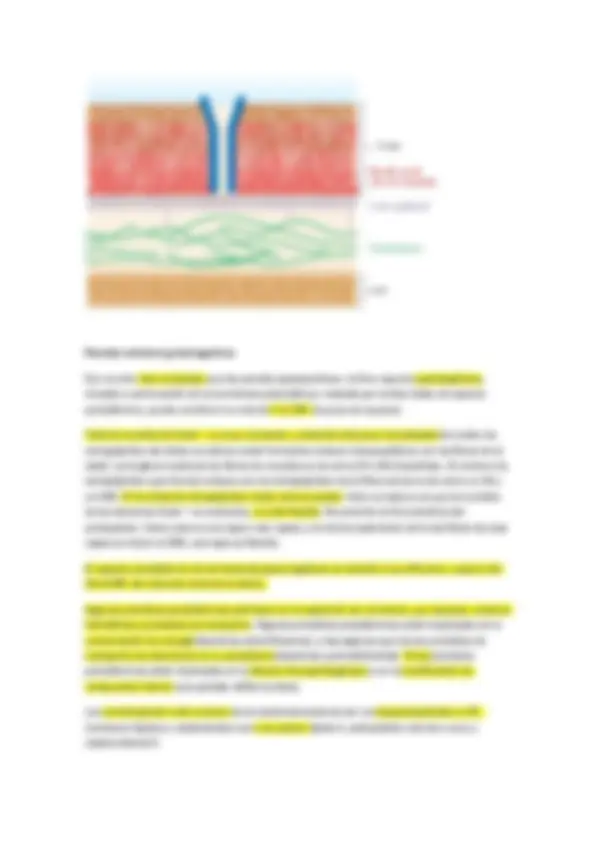
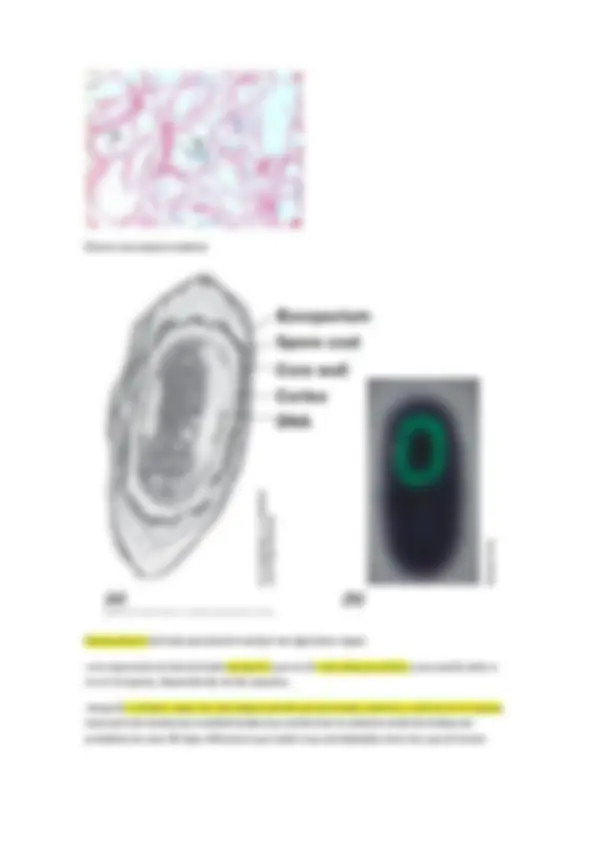
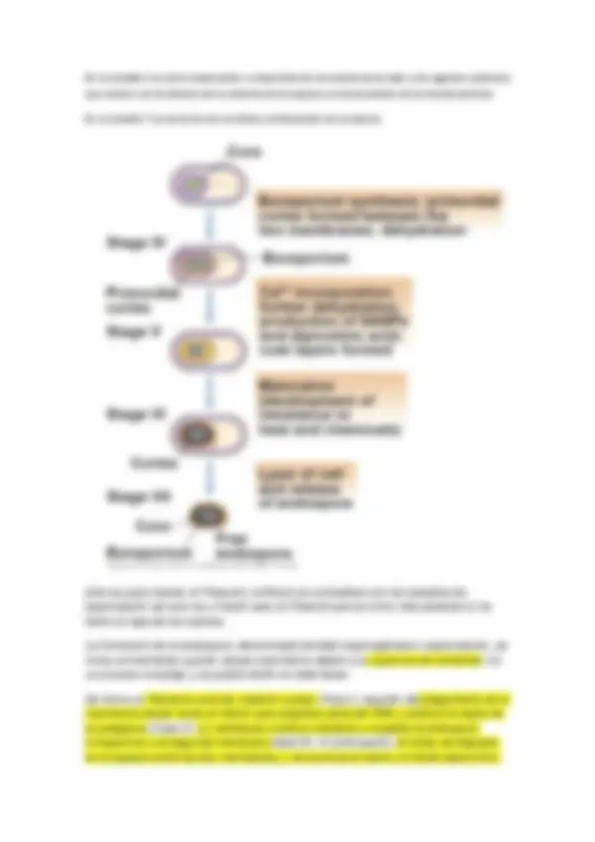
Estudio de la pared celular en procariotas
No hay que confundirla con la membrana citoplásmica (hay casos en los que la pared hay una ultima capa membranosa)
La pared es un componente casi universal en procariotas (99% de arqueas y bacterias que conocemos, hay casos particulares en los que arqueas y bacterias no presentan pared celular y esos organismos viven con limitaciones debido a esta ausencia)
Finalidad primaria de protegerse de la lisis osmótica que les ocurriría en la mayoría de hábitats donde se encuentran. Y otorgar a las células la forma característica, en ausencia de pared celular en forma de protoplastos serían esferas.
La mayoría de arqueas que no tienen pared celular son del género Thermoplasma
La mayoría de bacterias que no tienen pared celular son del género Mycoplasma (se puede encontrar en la saliva de mamíferos.
Uno de los pocos casos en los que encontramos esteroles en la membrana es en los mycoplasmas (hacen la membrana un poco más resistente a base de incorporar esteroles de su hospedador)
Las Arqueas presentan paredes celulares completamente diferentes a las de las bacterias