¡Descarga Nucleo y ciclo celular y más Apuntes en PDF de Biología Celular solo en Docsity!
NÚCLEO CELULAR (1)
PRINCIPALES ELEMENTOS DEL NÚCLEO. El núcleo celular está rodeado por la envuelta nuclear , compuesta por una membrana externa y otra interna. La externa es continua con el RER y presenta ribosomas en su cara citoplásmica. La interna descansa sobre la lámina nuclear. La mayor parte del interior del núcleo, lo que se denomina nucleoplasma , por analogía con el citoplasma, está constituída por cromatina. La cromatina transcripcionalmente activa se denomina eucromatina y está menos condensada que la heterocromatina , en la que la transcripción es muy difícil. El máximo grado de condensación es el que se observa en los cromosomas. El resto del nucleoplasma está ocupado por el nucleolo y la matriz nuclear (“nucleoesqueleto”), red de proteínas que mantiene la forma del núcleo y compartimentaliza la cromatina
LÁMINAS NUCLEARES. Son proteínas que pertenecen a la familia de los filamentos intermedios. En mamíferos hay tres tipos de láminas, A, B y C, que en su conjunto forman una red de nominada lámina nuclear. Las láminas interaccionan entre sí, con la cromatina, con proteínas de la membrana nuclear interna (lámina B) y con los complejos del poro. Cuando las láminas se fosforilan en sus extremos, la lámina se desorganiza (circunstancia que se produce durante la mitosis); los monómeros se disocian, si bien la lámina B queda anclada a la membrana de las vesículas que resultan de la fragmentación de la envuelta.
RELACIONES ENTRE EL NÚCLEO Y EL CITOPLASMA. El citoesqueleto y la lámina nuclear se mantienen comunicados por cadenas de proteínas enlazadoras, capaces de transmitir señales mecánicas desde el exterior celular hasta la propia cromatina. Algunas de dichas señales pueden modificar la propia transcripción del DNA.
IMPORTACIÓN Y EXPORTACIÓN DE PROTEÍNAS (información adicional)
TIPOS DE TRANSPORTE
NÚCLEO CELULAR (2): Transporte núcleo-citoplásmico
INTERCAMBIO ENTRE EL CITOPLASMA Y EL NÚCLEO. El citosol envía al núcleo las proteínas constitutivas de este (histonas, láminas, matriz, etc.), y las enzimas y factores necesarios para la duplicación (replicación) y transcripción del DNA. Por su parte, el núcleo exporta RNAs, que salen unidos a proteínas
COMPLEJO DEL PORO NUCLEAR (información adicional)
La entrada y salida de proteínas depende de los siguientes elementos:
- Secuencias de señalizacíon: La de entrada se llama secuencia de localización nuclear (NSL) y la de salida, secuencia de exportación nuclear (NES).
- Proteínas que interaccionan con el cargo, que se denominan importinas y exportinas.
- La proteína G- monomérica Ran: Regula la afinidad de las importinas y las exportinas con el cargo, permitiendo la liberación de este en el lugar apropiado (núcleo o citoplasma). Los GEFs predominan en el núcleo, por lo que es allí donde la la mayoría de Ran está en la forma Ran-GTP. Los GAPs son propios del citosol, por lo que en este predomina la forma Ran- GDP.
NUCLEOLO. El nucleolo es el lugar de formación de los ribosomas. En el se encuentra la mayor parte del DNA que transcribe el rRNA, si bien uno de los rRNAs de la subunidad mayor del ribosoma se transcribe fuera del nucleolo.
NÚCLEO CELULAR (4)
CICLO CELULAR (1)
DIAGRAMA DE LAS FASES DEL CICLO CELULAR.
PUNTOS DE
CONTROL DEL
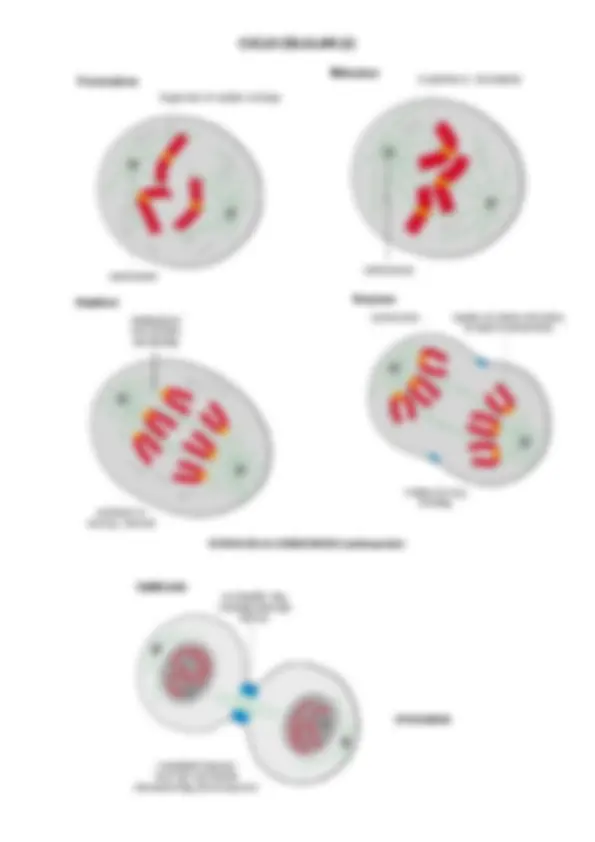
CICLO CELULAR. ETAPAS DE LA CARIOCINESIS (1)
CICLO CELULAR (2)
ETAPAS DE LA CARIOCINESIS (continuacIón)
CITOCINESIS
CICLO CELULAR (4)
FOSFORILACIONES Y DESFOSFORILACIONES EN UN COMPLEJO
CICLINA-Cdk (en este ejemplo, la M-Cdk.). La Cdk de la fase M y su ciclina forman un complejo inactivo (1), que sufre dos fosforilaciones inhibidoras consecutivas (2) y una fosforilación activadora (3). La activación de la Cdk tiene lugar por una fosfatasa, que elimina los fosfatos inhibidores (en este caso Cdc25), que se activa por la Cdk de la etapa anterior del ciclo (S-Cdk). La propia M-Cdk activa es capaz de aumentar la actividad de la fosfatasa Cdc25.
ESQUEMA GENERAL DEL CONTROL DEL CICLO CELULAR. Cada complejo Ciclina-Cdk activa al siguiente, asegurándose que el ciclo se verifica de forma ordenada. Además, cada Cdk fosforila, directamente, o a través de cascadas de quinasas, a otros sustratos. Por ejemplo, las S-Cdks, regulan a los factores y enzimas implicados en la replicación del DNA; las M-Cdks, fosforilan a las láminas (fragmentación de la membrana), a ciertas proteínas asociadas a microtúbulos (formación del huso acromático), a las condensinas (condensación de cromosomas) y al factor de promoción de anafase (APC). Los niveles de Cdks permanecen más o menos estables durante el ciclo y nunca son limitantes. Sin embargo, los niveles de ciclinas sí fluctúan, como se ve en la gráfica y sólo cuando alcanzan una concentración apropiada, permiten la activación de sus Cdks. Dicha fluctuación en los niveles depende, no solo de su síntesis, sino también de su degradación: APC es un complejo con actividad ubicuitín ligasa, que señala a las ciclinas para su degradación en el proteasoma. Mientras APC está activo (línea verde discontinua) las ciclinas son degradadas (ausentes desde la anafase hasta el punto de restricción de G1, “Start”). Por último, hay que recordar que la actividad de los complejos ciclina-Cdk, puede bloquearse por los inhibidores de Cdks (CKI). (Nota: La regulación de la actividad de las G1/S-Cdks se explicará más adelante, en relación a la presencia de factores de crecimiento).
COMPLEJO CICLINA-Cdk. La actividad de las quinasas del ciclo celular (Cdk) se regula por:
- Asociación a ciclinas: síntesis y degradación de estas
- Fosforilaciones inhibidoras (no ocurre en todas las Cdks)
- Fosforilaciones activadoras.
- Reclutamiento de inhibidores de Cdks (CKI)
CICLO CELULAR (5)
CICLO DEL COMPLEJO M-Cdk. El esquema ilustra la activación e inactivación de la Cdk mitótica. Ésta permanece inactiva durante G1, S y G2, pues la ciclina mitótica es degradada por APC. La concentración de la ciclina de mitosis aumenta gradualmente, hasta llegar a los niveles apropiados, al final de la fase G2. Entonces se forma el complejo M-Cdk, que se activa como consecuencia de las desfosforilaciones mediadas por Cdc
FUNCIONES DEL COMPLEJO DE PROMOCIÓN DE
ANAFASE. Entre los sustratos de APC se encuentra la proteína segurina , que secuestra a la enzima separasa. La separasa es la responsable de la proteolisis de las cohesinas. Una vez hidrolizadas las cohesinas, se separan las dos cromátidas, iniciándose la anafase. El otro sustrato esencial son las ciclinas.
RESUMEN DE LA TRANSICIÓN G2-M y METAFASE-ANAFASE. Como ya se ha indicado, el aumento gradual de la ciclina mitótica, junto con la activación final de la fosfatasa Cdc25 (activación mediada por las S-Cdks), activa a las M-Cdks. Estas últimas fosforilan y aumentan la actividad de la propia Cdc25 (retroalimentación positiva), así como los sustratos responsables de la entrada en mitosis (véase la figura de arriba a la izquierda). Como entre esos sustratos está APC, que se activa, al final se producirá la degradación de la ciclina mitótica y la inactivación de la M-Cdk, circunstancia necesaria para que el ciclo progrese.
CONTROL DE LA ACTIVIDAD DE APC. Entre las subunidades proteicas que forman el complejo APC se encuentran dos proteínas reguladoras: Cdc20 y Cdh1. Para que APC sea activo, se requiere que esté unido a cualquiera de ellas, circunstancia que no tiene lugar durante las fases S, G2 y parte de M (profase y metafase). Cdc20 se une y activa a APC cuando este último está fosforilado (por las M-Cdks). La consecuencia es que, como APC induce a la inactivación de las M-Cdks, a la larga produciría su propia inactivación. Esto sería catastrófico para el ciclo, pues como ya se apuntó anteriormente, para que no se produzcan errores, las ciclinas deben estar ausentes durante el final de la mitosis (anafase y telofase), la citocinesis, y la mayor parte de G1. Es ahora cuando entra en juego el otro activador, Cdh1 , que a diferencia de Cdc20, se une a APC cuando éste está desfosforilado y así sigue mandando al proteasoma a las ciclinas que pudieran quedar.