REV NEUROL 2004; 38 (10): 944-948944
INTRODUCCIÓN
Uno de los principales objetivos en el campo de las Neurocien-
cias es entender los mecanismos neuronales involucrados en la
codificación de la memoria. La mayoría de los neurofisiólogos
aceptan que el aprendizaje es el proceso por medio del cual
adquirimos nuevos conocimientos acerca de los eventos del
mundo, y la memoria se refiere a los procesos mediante los cua-
les retenemos y evocamos dichos conocimientos [1]. Indepen-
dientemente del tipo de memoria o de su contenido, se ha suge-
rido que por su duración existen al menos dos tipos: memoria
de corto y de largo plazo. Esta clasificación dependiente del
tiempo la propuso Hermann Ebbinhaus en 1885 y la formalizó
posteriormente William James, quién distinguió entre una me-
moria que duraba unas cuantas horas de una memoria que dura-
ba días, semanas o incluso años [2]. En 1900, Müller y Pilzec-
ker adicionaron el término ‘consolidación’ a esta clasificación
de memoria. De acuerdo con esta idea, la información reciente-
mente adquirida es frágil y susceptible de borrarse; sin embar-
go, la información se puede consolidar y convertirse en una
memoria estable de largo plazo [3].
Se ha propuesto que la información puede almacenarse
mediante cambios en la comunicación sináptica [4]. En la últi-
ma década del siglo XIX, Santiago Ramón y Cajal propuso una
teoría del almacenamiento de la información, que postula que la
información se almacena en el cerebro mediante cambios anató-
micos entre las conexiones de las neuronas [5]. Asimismo, She-
rrington propuso el término plasticidad sináptica para describir
los cambios en las propiedades funcionales de una sinapsis
como resultado de su actividad [5]; este término se refiere a la
posibilidad de inducir cambios que pueden o no ser reversibles.
En 1949, Donald Hebb formalizó esta idea, conocida como el
postulado de la sinapsis hebbiana: ‘Cuando el axón de la neuro-
na A está lo suficientemente cerca para excitar a la neurona B, y
repetida y persistentemente se da esta excitación, algunos pro-
cesos de crecimiento o cambios metabólicos tienen lugar en una
o en ambas células nerviosas, de tal manera que la eficacia de la
célula A para excitar a B se incrementa’ [6]. Asimismo, Hebb
postuló que las conexiones sinápticas pueden fortalecerse cuan-
do la neurona presináptica y postsináptica tienen una activación
temporal coincidente. Este fortalecimiento sináptico se ha
determinado como asociativo, porque asocia el disparo de una
neurona presináptica con el de una neurona postsináptica [5].
En este postulado Hebb manifiesta la necesidad de que ocurran
cambios estructurales para el mantenimiento de la memoria for-
mada como consecuencia del aprendizaje.
CAMBIOS EN LA EFICIENCIA
DE LA COMUNICACIÓN SINÁPTICA
Como ya se mencionó, se ha propuesto que el almacenamiento de
la información ocurre a través de cambios en la eficiencia de la
comunicación sináptica [4]. La potenciación a largo plazo (LTP,
por sus siglas en inglés) es un incremento sostenido en la fuerza
de la comunicación sináptica debido a la activación repetida de
una vía. La LTP se ha aceptado ampliamente como modelo de la
plasticidad sináptica que subyace el almacenamiento de la infor-
mación [7]. Por ejemplo, para inducir LTP se aplica a una vía afe-
rente excitatoria, breves episodios de estimulación de alta fre-
cuencia (100 Hz); esta estimulación, llamada tetánica de la vía,
trae como consecuencia un incremento duradero y estable en la
magnitud de la respuesta sináptica [8], que puede durar horas o
hasta días en animales intactos [4,9]. Como vemos, la LTP es una
forma de plasticidad sináptica dependiente de actividad que se ha
investigado principalmente en el hipocampo y la neocorteza, don-
de probablemente residen las modificaciones que subyacen a
ciertas formas de aprendizaje y memoria [5].
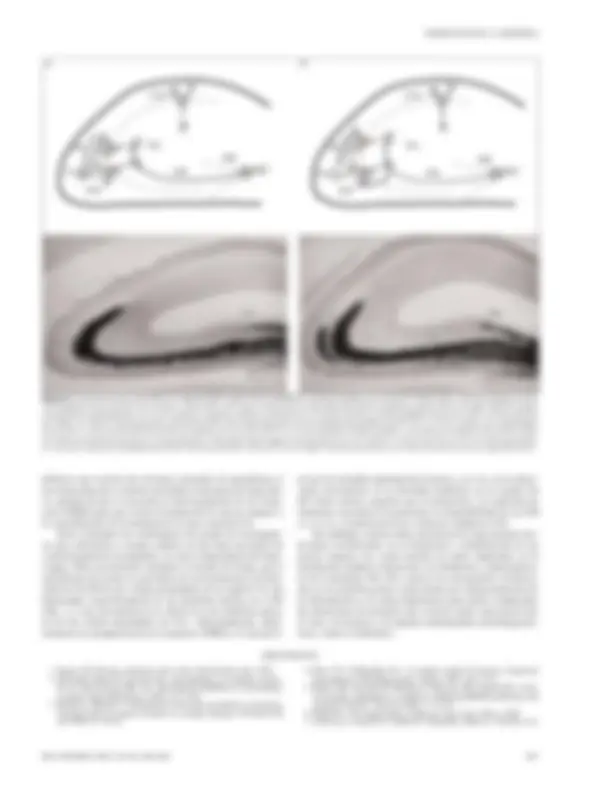
CAMBIOS ESTRUCTURALES
RELACIONADOS CON LA EXPERIENCIA
Los eventos plásticos también pueden incluir cambios en la es-
tructura, la distribución y el número de las sinapsis, y se ha su-
MEMORY-LINKED MORPHOLOGICAL CHANGES
Summary. Introduction. It has been suggested that storing information in the brain takes place by means of changes in synaptic
communication efficiency, which is known as neuronal plasticity. Plastic events include changes in the function, structure,
distribution and number of synapses, and it has been suggested that these plastic events could be related to learning and
memory. Development. In this work we will review some studies that report structural changes in which experience and
learning intervene. In particular, structural changes have been observed in a region of the brain called the hippocampus, which
plays a crucial role in the learning and memory of spatial tasks. It has been claimed that the appearance of new synapses after
learning a spatial task is linked to the formation of long-term memory and that the functioning of NMDA-type glutamate
receptors is needed for both learning and the formation of new synapses to take place. Conclusions. Understanding the cellular
mechanisms involved in the formation of memory is of utmost importance to be able to check the memory deficiencies that arise
from injuries or as a consequence of old age and neurodegenerative diseases. [REV NEUROL 2004; 38: 944-8]
Key words. Hippocampus. Memory. Morfological changes. NMDA receptors. Spatial learning. Synaptic plasticity. Synapto-
genesis.
Recibido: 06.04.04. Aceptado tras revisión externa sin modificaciones:14.04.04.
Departamento de Neurociencias. Instituto de Fisiología Celular. Universi-
dad Nacional Autónoma de México. México DF, México.
Correspondencia: Dr. Federico Bermúdez-Rattoni. Instituto de Fisiología
Celular. Universidad Nacional Autónoma de México. AP 70-253. 04510 Mé-
2004, REVISTA DE NEUROLOGÍA
Cambios morfológicos asociados a la memoria
I. Balderas, V. Ramírez-Amaya, F. Bermúdez-Rattoni
REVISIONES EN NEUROCIENCIA. EDITOR: J.V. SÁNCHEZ-ANDRES