¡Descarga QUITONES y más Apuntes en PDF de Biología solo en Docsity!
Original article
Late Pleistocene and Holocene chitons (Mollusca, Polyplacophora)
from Uruguay: Palaeobiogeography and palaeoenvironmental
reconstruction in mid latitudes of the southwestern Atlantic
Chitons (Mollusca, Polyplacophora) du Pléistocène tardif et de l’Holocène d’Uruguay :
paléobiogéographie et reconstruction paléoenvironnementale aux latitudes moyennes
de l’Atlantique du sud-ouest
Alejandra Rojas
a, *, Diego Urteaga
b a (^) Departamento de Evolución de Cuencas, Facultad de Ciencias, 598-2-5252646, Iguá 4225, CP 11400, Montevideo, Uruguay b (^) Museo Argentino de Ciencias Naturales, Bernardino Rivadavia, 54-11-49826670 ext. 138, Av. Ángel Gallardo 470, C1405DJR, Buenos Aires, Argentina Received 28 December 2009; accepted 27 September 2010 Available online 29 April 2011
Abstract
Here we present a revision of the fossil record of chitons (Mollusca, Polyplacophora) of Late Pleistocene and Holocene marine deposits of Uruguay and discuss their potential for palaeoenvironmental reconstruction. Chitons were recorded as isolated valves in bivalve- and gastropod- rich assemblages. They are represented by the species Chaetopleura angulata (Spengler), C. isabellei (d’Orbigny), C. asperrima (Gould) and Ischnochiton striolatus (Gray). The last two species are recorded for the first time as fossils not only in Uruguay but also in South America. Exclusively recorded for the Late Pleistocene is the warm water species I. striolatus, whose current southern range limit is located in Santa Catarina, Brazil. Higher temperatures than at present are inferred for the Uruguayan coast during the Late Pleistocene, probably driven by the southward influence of the warm Brazilian Current along the eastern margin of South America. Chitons from Holocene deposits show a wider geographic distribution along the Uruguayan coast in comparison to current distributions. They are recorded where an estuarine and eurihaline fauna lives today, which suggests the former existence of marine conditions. This is explained by the westward displacement of the marine front in the Uruguayan coast during the Holocene transgressive events. Chitons have proven to be useful palaeoenvironmental proxies for the reconstruction of salinity and temperature trends in the Uruguayan coast during the climatic oscillations of the Late Pleistocene and Holocene epochs. As such, their patterns of distribution are related to the recent physical evolution of the Southwestern Atlantic Ocean.
2011 Elsevier Masson SAS. All rights reserved.
Keywords: Quaternary; Palaeoenvironments; South America; Sea level oscillations; Chaetopleura; Ischnochiton
Résumé
Nous présentons ici une révision du registre fossile des chitons (Mollusca, Polyplacophora) de dépôts marins du Pléistocène tardif et de l’Holocène d’Uruguay, et discutons leur potentiel en matière de reconstruction paléoenvironnementale. Ces chitons sont enregistrés sous forme de valves isolées, au sein d’assemblages riches en bivalves et gastéropodes. Quatre espèces sont identifiées : Chaetopleura angulata (Spengler), C. isabellei (d’Orbigny), C. asperrima (Gould) et Ischnochiton striolatus (Gray). Les deux dernières espèces sont enregistrées pour la première fois comme fossiles, non seulement en Uruguay, mais également en Amérique du Sud. L’espèce d’eau chaude I. striolatus, dont la limite actuelle d’extension vers le Sud est située à Santa Catarina, Brésil, n’est enregistrée qu’au Pléistocène tardif. Des températures supérieures à l’actuel sont inférées pour la côte uruguayenne à cette époque, probablement dues à l’influence du Courant Brésilien chaud descendant vers le Sud le long de la marge Est de l’Amérique du Sud. Comparée à l’actuel, les chitons des dépôts holocènes montrent une distribution géographique plus large le long de la côte uruguayenne. Ils sont enregistrés là où se trouve aujourd’hui une faune estuarienne et euryhaline, ce qui suggère l’existence préalable de conditions marines liées au
Geobios 44 (2011) 377– 386
§ (^) Corresponding editor: Gilles Escarguel.
0016-6995/$ – see front matter # 2011 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.geobios.2010.09.
déplacement du front marin vers l’Ouest durant les événements transgressifs holocènes. Les chitons montrent leur utilité en tant qu’indicateur paléoenvironnemental pour la reconstruction des profils de salinité et de température de la côte uruguayenne durant les oscillations climatiques du Pléistocène tardif et de l’Holocène. Leur distribution est reliée à l’évolution physique récente de la partie sud-occidentale de l’Océan Atlantique.
2011 Elsevier Masson SAS. Tous droits réservés.
Mots clés : Quaternaire ; Paléoenvironnements ; Amérique du Sud ; Niveau de la mer ; Chaetopleura ; Ischnochiton
- Introduction
Chitons are an ancient group of marine molluscs that live exclusively on rocks or other hard substrates, with the majority of species living in intertidal to shallow subtidal habitats (Kaas and Van Belle, 1985). The fossil record of chitons remains poorly known as their occurrence as fossils is sporadic and occasional, especially when compared with the more diverse molluscan classes Bivalvia and Gastropoda. The oldest remains of this group extend back to the Upper Cambrian (Smith, 1960) and with the exception of some infrequent preservational conditions (e.g., Dell’Angelo et al., 2003; Pojeta et al., 2003; Vendrasco et al., 2004), most palaeontological records are based on disarticulated and isolated plates. The fossil record of chitons is also a poorly investigated topic and it has not been adequately assessed with respect to geographic areas or geologic time (Puchalski et al., 2008). Thus, new data on fossil chitons can be relevant to weigh their importance in past communities, and a good case in point is the scarcely studied but biogeographically interesting Southwestern Atlantic Ocean region. Indeed, the palaeobiogeography of chitons from Pleistocene and Holocene assemblages of the Uruguayan coast could aid in the reconstruction of palaeoenvironmental conditions of this coast in the recent past and provide a snapshot of the evolution of this highly dynamic area. The knowledge of living chitons from Uruguay is far from well-known. The main reason for this is the omission of chitons in faunistic studies, reduced sampling efforts and sampling adequacy, as well as the scarcity of local specialists to account for the diversity, distribution and ecology of living chitons along the Uruguayan coast (Scarabino, 2003). Most authors have recorded living chitons only in the counties of Maldonado and Rocha along the eastern shores of Uruguay (Barattini, 1951; Castellanos, 1951; Barattini and Ureta, 1961; Layerle and Scarabino, 1984) (Fig. 1(2)). The western record of chitons in the Río de la Plata estuary is from Carranza and Rodríguez (2007) who mention the presence of Chaetopleura angulata (Spengler, 1797) in Banco Inglés, a shallow-water sandy bank located 10 km south-east of Montevideo (Fig. 1(2)). Recent chitons living along the Uruguayan coast were summarized by Scarabino (2003) in a taxonomic list that took into account the available literature for C. angulata, C. asperrima (Couthouy MS, Gould, 1852), C. carrua Righi, 1970, C. isabellei (d’Orbigny, 1839), and Connexochiton moreirai (Righi, 1973). With the exception of the finding of an intermediate valve assigned to Chaetopleura sp. in a borehole core of the late Miocene Camacho Formation (Figueiras and Broggi, 1971; Sprechmann, 1978), the Uruguayan fossil record of chitons is restricted to deposits of late Quaternary age. Until now, little
attention has been paid to these late Quaternary fossil chitons as they constitute only a minor proportion of all invertebrates preserved in these assemblages. Instead, most taxonomic compilations (De Mata, 1947; Figueiras, 1962, 1967; Clavijo et al., 2005) as well as palaeoecological and palaeoenvironmental inferences (Teisseire, 1928; Sprechmann, 1978; Martínez, 1990; Martínez et al., 2001, 2006; Rojas, 2007) have come from the much more abundant and diverse bivalve and gastropod taxa. Chitons have only been briefly and superficially mentioned as part of these assemblages when their plates were occasionally found in the marine shell-beds. De Mata (1947) and later Figueiras (1962) recorded the presence of C. angulata (as C. tehuelcha d’Orbigny, 1841) from the Areneras de Carrasco locality in Canelones county. Sprechmann (1978) mentioned the presence of Chaetopleura isabellei and Chaetopleura sp. in a borehole core in Puerto de La Paloma, Rocha county. Finally, Piñeiro et al. (1992) and Rojas (2007) reported as Polyplacophora indet. the chiton plates recorded in Punta Rasa (Maldonado county) and La Coronilla (Rocha county), respectively (Fig. 1(3)). Until then, only the two most conspicuous species of chitons – C. angulata and C. isabellei – had been reported for the Quaternary marine deposits of Uruguay. Thus, we expected that a detailed analysis of previously collected fossil chitons from the Uruguayan Quaternary deposits along with new material collected for this study could provide a valuable untapped source of data on the diversity and palaeobiogeography of chitons along the Uruguayan coast. In the correlated Quaternary marine deposits of the adjacent Atlantic area of Argentina, chitons have also been reported as associated taxa from shell-beds. They come from Holocene sediments along the northeastern coast of Buenos Aires Province (Aguirre, 1990), from Bahía Blanca (Farinati,
- and along the northern coast of the Beagle Channel in Tierra del Fuego (Gordillo et al., 1992; Gordillo and Schwabe, 2009). The presence of chiton remains in Pleistocene deposits (locally referred to as ‘‘Belgranense’’) was reported by Bordas (1957) for the Buenos Aires Province. The aim of this paper is to provide a review and first compilation of the fossil record of chitons from the littoral Quaternary deposits of Uruguay. We describe the species that occur in these deposits, assess their chronological and palaeobiogeographic distribution along the Uruguayan coast, and test their potential utility for palaeoen- vironmental reconstruction in mid latitudes of the Southwestern Atlantic Ocean.
- Geological setting and study area
The studied chitons come from the Quaternary deposits included in the Chuy and/or Villa Soriano formations in
Pleistocene deposits (with minimum radiocarbon ages available of ca. 30,000–35,000 14 C years before present) and more abundant and widespread Holocene deposits that range discontinuously in age between approximately 10,000 and 1800 cal. years before present. These fossiliferous deposits have been correlated with adjacent Atlantic units from Argentina and Brazil (Goñi and Hoffstetter, 1964; Forti Esteves, 1974; Martínez, 1990; Aguirre and Whatley, 1995). The study area comprises the Uruguayan coastal waters currently dominated by the fluvio-marine gradient of the Río de la Plata and Atlantic Ocean (Fig. 1(2)). The Río de la Plata estuary receives from the west a freshwater discharge from the Paraná and Uruguay rivers, and from the east the marine waters of the Atlantic Ocean (Urien, 1972). It is divided by a submerged shoal (Barra del Indio) into an inner fluvial system and an outer mixohaline brackish system (Mianzan et al., 2001). The biogeography of the living biota responds to this gradient with a west to east dominance of freshwater, estuarine and marine taxa, respectively (Masello and Menafra, 1998; Giménez et al., 2005; Brazeiro et al., 2006). The limits and position of the salinity zones vary according to wind balance, river discharge and season (Guerrero et al., 1997). In the marine front, the large-scale dynamics of the Southwestern Atlantic Ocean are dominated by the southward flowing Brazilian Current and the northward flowing Malvinas (Falkland) Current. The former transports tropical and sub- tropical waters along the continental margin of South America, while the latter transports cold subantarctic waters along the Argentinean shelf (Olson et al., 1988; Odebrecht and Castello, 2001). Both currents meet at the Brazil-Malvinas Confluence Zone (between 25 8 S and 45 8 S), resulting in highly variable physicochemical and biological attributes on the shelf and slope. The temperature gradients related to these water masses outline the current malacological provinces of the Southwestern Atlantic, the warm water Brazilian Province, the confluence Argentinean Province and the cold Magallanic Province (i.e. Scarabino, 1977; Briggs, 1995; Martínez and del Río, 2002). Coastal studies in Brazil, Argentina and Uruguay have shown that faunal compositions include a combination of species of warm-temperate and cool-temperate affinities. In this scenario, the Río de la Plata acts as an ecologic barrier and represents a broad ecotone between the southern and northern areas (Masello and Menafra, 1998; Scarabino et al., 2006a, 2006b).
- Material and methods
The chiton remains studied herein are housed in the Colección de Invertebrados Fósiles de la Facultad de Ciencias (FCDPI), in Montevideo, Uruguay. The material was obtained in bulk samples of about 3 dm 3 by previous authors (Piñeiro et al., 1992; Martínez et al., 2001, 2006; Rojas, 2007) and additional material was recovered during further fieldwork. Samples were sieved in water and the residues dried at room temperature; some specimens were further cleaned using an ultrasonic bath. Chiton remains were photographed using a Nikon D100 digital camera equipped with a Nikkor 28–85 mm macro lens and a Philips XL30 scanning electron microscope at the Museo Argentino de Ciencias Naturales (MACN); they were measured by means of a digital caliper. All specimens available were determined (when possible) to species level. The systematics follows Sirenko (2006). The area covered by the prospective studies (previous and this work) represents much of the geographic extension of the Quaternary littoral deposits of Uruguay (see above). The localities yielding chiton plates are shown in Fig. 1(3). Two of them (Punta Espinillo and Areneras de Carrasco) were radiocarbon dated by Acceleration Mass Spectrometry at the NSF Arizona AMS Facility, University of Arizona, United States. Ages are shown in Table 1 and were calibrated using the program CALIB REV5.0.2 (Stuiver and Reimer, 1993) with the Hughen et al. (2004) calibration datasets with a 19 � 13 delta R assumed.
- Systematic palaeontology
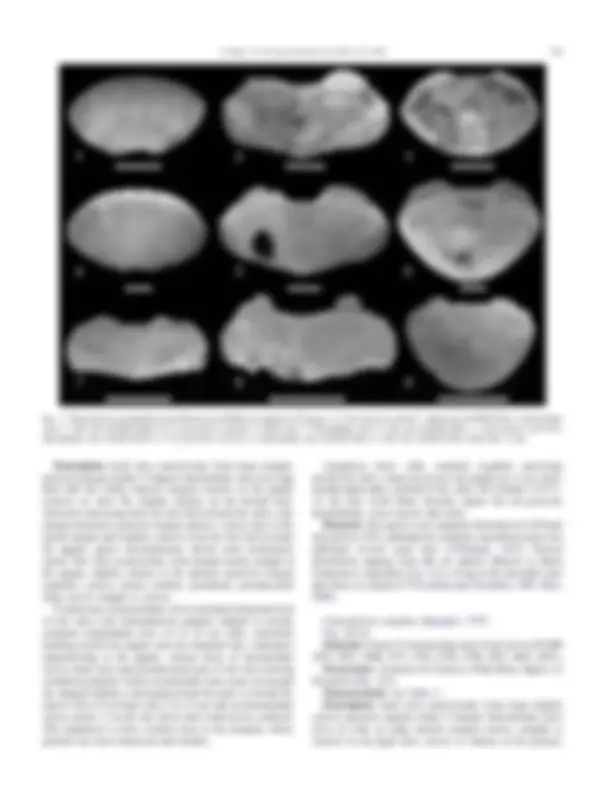
Class POLYPLACOPHORA Gray, 1821 Order CHITONIDA Thiele, 1910 Family CHAETOPLEURIDAE Plate, 1899 Genus Chaetopleura Shuttleworth, 1853 Chaetopleura isabellei (d’Orbigny, 1839) Fig. 2(1–3) Material: 8 head, 66 intermediate and 2 tail valves (FCDPI 2877, 2895, 3152, 4794, 4797, 4850, 4858, 4928, 4994). Occurrence: Punta Espinillo, Areneras de Carrasco, Punta Fría, Punta Rasa, Saglia, La Coronilla (Fig. 1(3)). Measurements: see Table 2.
Table 1 Chiton species recorded in Quaternary marine deposits of Uruguay by locality and age.
Chiton species a^ Locality 14 C age 95.4% (2s) cal. age BP Lab. Nr. Taxon dated Age source
1 Punta Espinillo 3501 � 40 3251 – 3465 AA83199 Natica isabelleana This work 1, 2, 3 Areneras de Carrasco 7379 � 44 7710 – 7928 AA83200 Anomalocardia brasiliana This work 1 Punta Fría 2490 � 70 1858 – 2276 LP-817 Tegula patagonica Martínez et al. (2006) b 1, 2 Punta Rasa - - - - - 2 Saglia 1 3530 � 50 3148 – 3468 LP-907 Erodona mactroides Martínez et al. (2006) 2 Saglia 2a 4460 � 70 4312 – 4786 LP-893 Erodona mactroides Martínez et al. (2006) 1, 2 Saglia 2b 4800 � 70 4798 – 5241 LP-901 Erodona mactroides Martínez et al. (2006) 1, 2, 3, 4 La Coronilla 29.500 � 600 Min. age LP-884 Ostreola equestris Martínez et al. (2001) 35.500 � 1900 Min. age LP-824 Mactra isabelleana Martínez et al. (2001) a (^) 1: C. isabellei; 2: C. angulata; 3: C. asperrima; 4: I. striolatus. b (^) Martínez et al. (2006) referred as ‘‘Punta Rasa’’ the age obtained for Punta Fría by mistake.
Description: head valve semicircular, front slope straight, posterior margin widely V-shaped. Intermediate valve less long than half the width; anterior margin concave at the jugum (convex on valve II), slightly sinuose on the pleural area, obviously narrowing from the last third toward the sides; side margin truncated, posterior margin sinuose: convex near to the lateral margin and slightly concave from the first third toward the jugum; apices inconspicuous; lateral areas moderately raised. Tail valve semicircular; front margin nearly straight at the jugum, slightly sinuose at the pleurae; posterior margin regularly convex; mucro median, prominent; postmucronal slope can be straight or convex. Central area of intermediate valves and anteromucronal area of tail valve with hemispherical granules aligned in mostly complete longitudinal rows (14 to 23 per side), somewhat bending toward the jugum near the diagonal line, sometimes anastomosing at the jugum. Lateral areas of intermediate valves, head valve and postmucronal area of tail valve bearing cylindrical pustules which exceptionally bear a pore on top and are aligned radially, converging toward the apex or toward the mucro (30 to 47 on head valve, 5 to 13 per side on intermediate valves, about 17 on the tail valves that could not be counted). This alignment is more evident close to the margins, where pustules are more numerous and smaller.
Apophyses short, wide, rounded, regularly narrowing toward the sides, connected across the jugum by a very short, straight jugal plate, notched at the sides; slit formula 11/1/11- 12, slit rays, teeth blunt, dorsally rugose but not grooved, inequidistant, eaves narrow and solid. Remarks: this species was originally illustrated in 1839 and described in 1841, although the complete expedition report was published several years later (d’Orbigny, 1847). Present distribution ranging from Rio de Janeiro (Brazil) to Punta Camarones (Argentina; Fig. 1(1)); living in the intertidal zone and down to a depth of 70 m (Kaas and Van Belle, 1987; Ríos, 2009).
Chaetopleura angulata (Spengler, 1797) Fig. 2(4–6) Material: 6 head, 53 intermediate and 18 tail valves (FCDPI 2479, 3073, 3098, 3477, 4793, 4795, 4796, 4857, 4891, 4927). Occurrence: Areneras de Carrasco, Punta Rasa, Saglia, La Coronilla (Fig. 1(3)). Measurements: see Table 2. Description: head valve semicircular, front slope slightly convex, posterior margin widely V-shaped. Intermediate valve twice as wide as long; anterior margin convex, straight or concave in the jugal area, convex or sinuose at the pleurae,
[(Fig._ 2 )TD$FIG]
Fig. 2. Chiton species recorded from Late Pleistocene and Holocene deposits of Uruguay. 1–3. Chaetopleura isabellei. 1. Head valve (FCDPI 4794). 2. Intermediate valve. 3. Tail valve (FCDPI 4858). 4–6. Chaetopleura angulata. 4. Head valve. 5. Intermediate valve. 6. Tail valve (FCDPI 4857). 7. Chaetopleura asperrima. Intermediate valve (FCDPI 4859). 8, 9. Ischnochiton striolatus. 8. Intermediate valve (FCDPI 4861). 9. Tail valve (FCDPI 4934). Scale bars: 2 mm.
A. Rojas, D. Urteaga / Geobios 44 (2011) 377– 386 381
among the chiton taxa recorded in the Quaternary deposits of Uruguay, while further studies are ongoing in order to clarify its taxonomic status. Present distribution between Espírito Santo (Brazil) and Maldonado (Uruguay) (Fig. 1(1, 2)); it has been collected in the intertidal zone, down to a depth of almost 50 m (Kaas and Van Belle, 1987; Ríos, 2009).
Family ISCHNOCHITONIDAE Dall, 1889 Genus Ischnochiton Gray, 1847 Ischnochiton striolatus (Gray, 1828) Fig. 2(8, 9) Material: 5 intermediate valves and 1 tail valve (FCDPI 4861, 4934). Occurrence: La Coronilla (Fig. 1(3)). Measurements: see Table 2. Description: intermediate valve broadly rectangular, shorter than half the width; front margin concave in the jugal part, straight to slightly convex at the pleurae; side margins more or less rounded; hind margin almost straight, lateral areas weakly raised. Tail valve less than semicircular; length more than half the width; front margin convex; mucro central; hind slope concave. Weak tegmental sculpture, central areas of intermediate valves and anteromucronal area of tail valve bearing fine granules arranged in lines starting perpendicular to the diagonal line converging toward and across the jugum, becoming slightly stronger near the diagonal ridge, thus forming a quincunx. Lateral areas of intermediate valves and postmucronal area of tail valve roughly granulose, somewhat scaly; concentric grooves barely perceptible. Aphophyses short, inequilaterally triangular; jugal sinus straight about one third of valve’s width; insertion plates short; slit formula ?/1/11, slits inequidistant, slit rays distinct; intermediate valves have several aesthetes openings which are arranged forming a triangle along the jugal area of articulamentum; teeth sharp, smooth; eaves narrow, solid. Remarks: present distribution between Bogue Inlet, North Carolina (34 8 40’N) and Santa Catarina, Brazil (27 8 40’S) (Fig. 1(1)). Intertidal to shallow subtidal (Kaas and Van Belle, 1990; Ríos, 2009).
- Discussion
Four species of Polyplacophora lived on the Uruguayan coast during the late Quaternary. This number suggests a low richness when compared to the southernmost South American fossil chiton assemblage reported by Gordillo and Schwabe (2009), but this richness increases when compared with the fossil record of chitons in adjacent Atlantic areas of Argentina. For example, in northeastern Buenos Aires Province only C. isabellei was recorded in Holocene sediments (Aguirre, 1990), while further south in Bahía Blanca, this species was found together with C. angulata (Farinati, 1995). Until now, no chitons were reported from adjacent Brazilian Quaternary deposits. Considering the living chitons in Uruguayan waters, Scarabino (2003) listed five species. Later, Demicheli and
Scarabino (2006) reconsidered the nominal Chaetopleura species and argued that, apart from C. angulata, the members of this genus in this region were still in need of a detailed revision. In particular, C. carrua and C. asperrima each needs to be confirmed as distinct from C. isabellei, and if so, each of their respective ranges needs to be documented. The presence of C. moreirai in Uruguayan waters cannot be confirmed (F. Scarabino, pers. comm. 2009) because the material found by Layerle and Scarabino (1984) is no longer available for study. Also, the lack of any new specimens of C. moreirai from sampling of extant fauna in the originally prospected area, makes its record at least suspicious. Unlike the neontologic information available, this study provides a reliable geographic and temporal context for the specimens and species represented along the coast of Uruguay during the Late Pleistocene and Holocene. It confirms the presence of C. isabellei in both epochs (Table 1). The fossil record of C. isabellei and C. angulata along the Uruguayan coast increases their geographic range as they were found at localities from which they have not been previously reported (e.g. Punta Espinillo, Punta Fría, Punta Rasa, Saglia and La Coronilla). I. striolatus is reported from the Pleistocene of Uruguay for the first time, and C. asperrima is newly recorded from Holocene mollusc assemblages. The findings presented here also constitute the first fossil record of these two species for South America.
5.1. Palaeobiogeography and palaeoenvironmental reconstruction
The palaeobiogeographic distribution of chitons along the Uruguayan coast during the late Pleistocene and Holocene can be compared to the corresponding extant species, to qualitatively show shifts in salinity gradients related to the Río de la Plata, as well as shifts in temperature due to changing Southwestern Atlantic current regimes.
5.1.1. Pleistocene Pleistocene chitons are only present in La Coronilla (Fig. 1(3)), represented by the four species recorded in this study. For this deposit only minimum radiocarbon ages are available (Martínez et al., 2001; Table 1). The bivalve and gastropod taxa preserved are indicative of warmer conditions than today, thus Martínez et al. (2001) and Rojas (2007) inferred a probable Last Interglacial (OIS 5e) range of dates for the assemblage, between 130,000 and 119,000 years BP (Hearty et al., 2007). The presence of I. striolatus in this assemblage has some paleoenvironmental implications as it lives in warmer water than that off the present Uruguayan coast. This species is found from intertidal to sublittoral depths (Gracia et al., 2005; Rodrigues and Absalão, 2005) with a present southern distribution limit along the Southwestern Atlantic at Santa Catarina, Brazil (Ríos, 2009). Our record of this species as fossil could imply that the coast of Uruguay had warmer temperatures than today during the Late Pleistocene. Then, the current absence of I. striolatus from the living benthic
communities of the Uruguayan coast (Scarabino, 2003; Ríos, 2009), represents a more recent northward range retraction. The presence of 10 co-occurring bivalves and gastropods in the La Coronilla assemblage with a similar biogeographic pattern (Rojas, 2007) further supports this scenario. Similar changing climatic conditions in the Pleistocene had also triggered equivalent biogeographic responses of molluscan faunas in other geographic areas (Valentine and Jablonski, 1993; Roy et al., 1995; Kitamura and Ubukata, 2003). The mechanism proposed to explain this scenario is the displacement of the convergence zone of the Brazil and Malvinas currents toward higher latitudes (Martínez et al., 2001; Rojas, 2007), which would account for the fact that chiton species might have experienced shifts in their range limits, probably due to their adaptation to particular thermal regimes. The changing current regime along the western Atlantic coast of South America could represent widespread southward range extensions along the northern coast of Argentina, reflected in the composition of Pleistocene invertebrate assemblages. However, the still limited sampling effort for Pleistocene deposits along the Buenos Aires coast (Aguirre and Whatley, 1995) prevents any reconstruction of this kind at the moment. The absence of I. striolatus from Holocene assemblages (see below) could indicate a weaker influence of the warm Brazilian Current along the Uruguayan coast in this time interval. Although sampling failure and/or taphonomic agents might account for this, the greater abundance, degree of knowledge and sampling effort in the Holocene deposits of Uruguay (Sprechmann, 1978; Martínez et al., 2006; Rojas, 2007) favors the shifting climatic regime hypothesis.
5.1.2. Holocene Chitons from the Holocene assemblages of Punta Espinillo, Areneras de Carrasco, Punta Fría, Punta Rasa and Saglia (Fig. 1(3)) are represented by three species of the genus Chaetopleura: C. angulata, C. isabellei and C. asperrima. Radiocarbon ages are available for all localities with the exception of Punta Rasa (Table 1). The ages available for Saglia suggest that C. isabellei and C. angulata lived at the current western border of the Merín Lagoon between approximately 5200 to 3150 cal. years BP, and that marine conditions prevailed where a freshwater lagoon presently occurs. The current western limit of distribution of chitons along the Uruguayan coast (C. angulata, cited as Stereochiton felipponei by Figueiras and Sicardi, 1968) is located at Punta Negra in Maldonado county (Fig. 1(2)). Very close to this locality occurs the youngest record of fossil chitons, at Punta Rasa and Punta Fría approximately at 2200–1800 cal. years BP (Fig. 1(3)). However, the past western boundary of distribution of chitons is located at Punta Espinillo at ca. 3200–3500 cal. years BP, stretching their coastal distribution limit along the Uruguayan coast by approximately 120 km into the Río de la Plata (Fig. 1(2, 3)). A similar pattern occurs at Areneras de Carrasco, the oldest Holocene chiton-bearing deposit (ca. 7700–7900 cal. years BP).
These findings significantly expand the geographic range occupied by chitons along the Uruguayan coast during at least part of the Holocene. As described above, the main physical variable that accounts for the current biogeographic distribution along the Uruguayan coast is salinity (Masello and Menafra, 1998; Giménez et al., 2005; Brazeiro et al., 2006). Along this gradient, Punta Espinillo and Areneras de Carrasco are located within the current estuarine range of the Río de la Plata (Calliari et al., 2003; Giménez et al., 2005). As stated earlier, chitons have only been reported from the shores of the eastern Maldonado and Rocha counties, where the coast receives a strong marine influx from the Atlantic Ocean (Fig. 1(2)). The only exception to this is the record by Carranza and Rodríguez (2007) of C. angulata at Banco Inglés, 10 km south-east off the coast of Montevideo, whose presence can be explained by the highly variable salinity of the area through the occurrence of a quasi-permanent salt wedge regime that generates a bottom salinity front (Guerrero et al., 1997; Mianzan et al., 2001). Since polyplacophorans are a strictly marine group (stenopolyhaline sensu Kaas and Van Belle, 1985), their palaeobiogeography points to the existence of conditions with higher salinity during part of the Holocene that allowed the colonization and establishment of taxa west of their known current coastal distribution. This evidence supports a westward advance of the marine front on the Uruguayan coast inferred also by the palaeoecological analysis of bivalve and gastropod assemblages from these and other Holocene littoral deposits (Martínez et al., 2001, 2006; Rojas, 2007). The low number of assemblages containing chiton remains and the absence of a continuum of absolute ages precludes a more accurate analysis of salinity changes throughout the Holocene. The data currently available are also not sufficient to carry out a species level biogeographic characterization of the Uruguayan coast for the recent past. However, this study shows qualitatively that the palaeoecology and palaeobiogeography of chitons has a potential to provide insights for the reconstruction of the palaeoenvironmental changes occurring in the South- western Atlantic Ocean.
- Conclusions
C. angulata, C. isabellei and C. asperrima lived along the Uruguayan coast during transgressive Holocene events, which together with I. striolatus occupied the coast in the Late Pleistocene. The two latter species are reported for the first time for the South American fossil record. For the Late Pleistocene the warm water I. striolatus indicates higher temperatures than the present for the Uruguayan coast, probably driven by the southward influence of the warm Brazilian Current along the western margin of South America. The Holocene geographic distribution of chitons along the Uruguayan coast was wider than at present. The presence of chitons implies the existence of marine conditions where today only estuarine and euryhaline species live, and this likely reflects the westward displacement of the marine front during Holocene transgressive events.
Hearty, P.J., Hollin, J.T., Neumann, A.C., O’leary, M.J., McCulloch, M., 2007. Global sea-level fluctuations during the Last Interglaciation (MIS 5e). Quaternary Science Reviews 26, 2090–2112. Hughen, K.A., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand, C., Blackwell, P.G., Buck, C.E., Burr, G., Cutler, K.B., Damon, P.E., Edwards, R.L., Fairbanks, R.G., Friedrich, M., Guilderson, T.P., Kromer, B., McCor- mac, F.G., Manning, S., Bronk Ramsey, C., Reimer, P.J., Reimer, R.W., Remmele, S., Southon, J.R., Stuiver, M., Talamo, S., Taylor, F.W., van der Plicht, J., Weyhenmeyer, C.E., 2004. Marine04 marine radiocarbon age calibration, 0–26 cal kyr BP. Radiocarbon 46, 1059–1086. Kaas, P., 1954. Notes on Loricata 2. On the occurence of Chaetopleura fulva (Wood) on the eastern coast of Latin America. Basteria 18, 14–19. Kaas, P., Van Belle, R.A., 1985. Monograph of living chitons (Mollusca, Polyplacophora). 1 Order Neoloricata: Lepidopleurina. E.J. Brill, Leiden. Kaas, P., Van Belle, R.A., 1987. Monograph of Living Chitons (Mollusca: Polyplacophora). 3 Suborder Ischnochitonina, Ischnochitonidae: Chaeto- pleurinae, and Ischnochitoninae (pars). Additions to vols 1 and 2. E.J. Brill, Leiden. Kaas, P., Van Belle, R.A., 1990. Monograph of Living Chitons (Mollusca: Polyplacophora). 4 Suborder Ischnochitonina: Ischnochitonidae: Ischno- chitoninae (continued). Additions to vols 1, 2 and 3. E.J. Brill, Leiden. Kitamura, A., Ubukata, T., 2003. The sequence of local recolonization of warm- water marine molluscan species during a deglacial warming climate phase: a case study from the early Pleistocene of the Sea of Japan. Palaeogeo- graphy, Palaeoclimtology, Palaeoecology 199, 83–94. Layerle, C., Scarabino, V., 1984. Moluscos del frente marítimo uruguayo entre 9 y 78 m de profundidad: análisis biocenológico. Contribuciones (Departa- mento de Oceanografía, Facultad de Humanidades y Ciencias) 1 (9), 1–17. Martínez, S., 1990. Taphonomy and Palaeoecology of Holocene mollusks from the western margin of the Merin Lagoon (Villa Soriano Fm., Uruguay). Quaternary of South America and Antarctic Peninsula 7 (121), 135. Martínez, S., del Río, C.J., 2002. Late Miocene molluscs from the Southwestern Atlantic Ocean (Argentina and Uruguay): a palaeobiogeographic analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 188, 167–187. Martínez, S., Rojas, A., Ubilla, M., Verde, M., Perea, D., Piñeiro, G., 2006. Molluscan assemblages from the marine Holocene of Uruguay: composition, geochronology, and palaeoenvironmental signals. Ameghiniana 43, 385–397. Martínez, S., Ubilla, M., 2004. El Cuaternario de Uruguay. In: Veroslavsky, G., Ubilla, M., Martínez, S. (Eds.), Cuencas Sedimentarias de Uruguay: Geología, Palaeontología y Recursos Naturales, Cenozoico. DIRAC, Montevideo, pp. 195–228. Martínez, S., Ubilla, M., Verde, M., Perea, D., Rojas, A., Gueréquiz, R., Piñeiro, G., 2001. Palaeoecology and geochronology of Uruguayan coastal marine pleistocene deposits. Quaternary Research 55, 246–254. Masello, A., Menafra, R., 1998. Comunidades macrobentónicas de la zona costera uruguaya y áreas adyacentes. In: Wells, P.G., Daborn, G.R. (Eds.), El Río de la Plata – Una revisión ambiental. Un informe del Proyecto ECOPLATA. Dalhousie University, Halifax, Nova Scotia, pp. 117–167. Mianzan, H., Lasta, C., Acha, M., Guerrero, R., Macchi, G., Bremen, C., 2001. The Río de la Plata Estuary, Argentina-Uruguay. In: Seeliger, U., Kjerfve, B. (Eds.), Coastal Marine Ecosystems of Latin America. Springer-Verlag, Berlin, Heidelberg, pp. 185–204. Odebrecht, C., Castello, J.P., 2001. The Convergence Ecosystem in the South- west Atlantic. In: Seeliger, U., Kjerfve, B. (Eds.), Coastal Marine Ecosys- tems of Latin America. Springer-Verlag, Berlin, Heidelberg, pp. 147–165. Olson, D.B., Podestá, G.P., Evans, R.H., Brown, O.B., 1988. Temporal varia- tions in the separation of Brazil and Malvinas Currents. Deep-Sea Research 35, 1971–1990. Piñeiro, G., Scarabino, F., Verde, M., 1992. Una nueva localidad fosilífera del Holoceno marino de Uruguay (Punta Rasa, Departamento de Maldonado). Boletín de la Sociedad Zoológica del Uruguay 7, 61–62. Plate, L.H., 1898–1901. Die anatomie und phylogenie der chitonen. Fauna chilensis. Zoologische Jahrbücher Supplement 5, 156–216. Pojeta, J., Eernisse, D.J., Hoare, R.D., Henderson, M.D., 2003. Echinochiton dufoei: a new spiny Ordovician chiton. Journal of Paleontology 77, 646–654. Puchalski, S.S., Eernisse, D.J., Johnson, C.C., 2008. The effect of sampling bias on the fossil record of chitons (Mollusca, Polyplacophora). American Malacological Bulletin 25, 87–95.
Righi, G., 1967. Sobre Polyplacophora do litoral brasileiro. Papéis Avulsos de Zoologia 20, 85–98. Righi, G., 1970. Mollusques Polyplacophores. Résultats scientifiques des campagnes de la ‘‘Calypso’’ (9). Campagne de La Calypso au large des côtes atlantiques de l’Amérique du Sud (1961–1962). Annales de l’Institut Océanographique de Monaco 47, 107–114. Righi, G., 1973. Adições aos Polyplacophora brasileiros (Mollusca). Papéis avulsos de Zoologia (São Paulo) 26, 259–273. Ríos, E., 2009. Compendium of Brazilian Sea Shells. Evangraf, Rio Grande do Sul. Rochebrune de, A.T., 1889. Polyplacophores. In: Rochebrune, A.T., Mabille, J. (Eds.), Mission Scientifique du Cap Horn 1882–1883, 6 (Zoologie). Gau- thiers-Villars et fils, Paris, pp. 131–143. Rodrigues, L.R.G., Absalão, R.S., 2005. Shell colour polymorphism in the chiton Ischnochiton striolatus (Gray, 1828) (Mollusca: Polyplacophora) and habitat heterogeneity. Biological Journal of the Linnean Society 85, 543–548. Rojas, A., 2007. Moluscos de aguas cálidas del Cuaternario Marino del Uruguay. M.Sc. thesis, PEDECIBA, Universidad de la República (unpub- lished). Roy, K., Jablonski, D., Valentine, J.W., 1995. Thermally anomalous assem- blages revisited: patterns in the extraprovincial latitudinal range shifts of Pleistocene marine mollusks. Geology 23, 1071–1074. Scarabino, F., 2003. Lista sistemática de los Aplacophora, Polyplacophora y Scaphopoda de Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay 8 (78–79), 191–196. Scarabino, F., Zafaroni, J.C., Carranza, A., Clavijo, C., Nin, M., 2006a. Gasterópodos marinos y estuarinos de la costa uruguaya: faunística, dis- tribución, taxonomía y conservación. In: Menafra, R., Rodríguez, L., S- carabino, F., Conde, D. (Eds.), Bases Para la Conservacion y el Manejo de la Costa Uruguaya. Vida Silvestre, Montevideo, pp. 143–155. Scarabino, F., Zaffaroni, J.C., Clavijo, C., Carranza, A., Nin, M., 2006b. Bivalvos marinos y estuarinos de la costa uruguaya: faunística, distribu- ción, taxonomía y conservación. In: Menafra, R., Rodríguez, L., Scar- abino, F., Conde, D. (Eds.), Bases Para la Conservacion y el Manejo de la Costa Uruguaya. Vida Silvestre, Montevideo, pp. 157–169. Scarabino, V., 1977. Moluscos del Golfo San Matías (Provincia de Río Negro, República Argentina). Inventario y claves para su identificación. Comuni- caciones de la Sociedad Malacológica del Uruguay 4, 177–285. Shuttleworth, R.J., 1853. Diagnosen neuer Mollusken. 4, Ueber den Bau der Chitoniden, mit Aufzählung der die Antillen und die Canarischen Inseln bewohnenden Arten. Mitteilungen der naturforschenden Gesellshaft in Bern 286/291, 169–207. Sirenko, B.I., 2006. New Outlook on the System of Chitons (Mollusca: Polyplacophora). Venus 65, 27–49. Smith, A.G., 1960. Amphineura. In: Moore, R.C. (Ed.), Treatise on Invertebrate Palaeontology, Part 1: Mollusca 1. Geological Society of America, New York, pp. 141–176. Spengler, L., 1797. Udførlig Beskrivelse over det mangeskallede Konchylie– Slægt, af Linneus kaldet Chiton; med endeel nye Arter og Barieteter. Skrivter af Naturhistorie-Selskabet 4, 62–103. Sprechmann, P., 1978. The palaeoecology and palaeogeography of the Uru- guayan coastal area during the Neogene and Quaternary. Zitteliana 4, 3–72. Stuiver, M., Reimer, P.J., 1993. Extended 14 C database and revised CALIB radiocarbon calibration program. Radiocarbon 35, 215–230. Teisseire, A., 1928. Contribución al estudio de la geología y la palaeontología de la República Oriental del Uruguay. Región de Colonia. Anales de la Universidad 122 (series 37), 319–469. Thiele, J., 1909–1910. Revision des Systems der Chitonen. Zoologica Stuttgart 22, 1–70. Urien, C.M., 1972. Río de la Plata estuary environments. Geological Society of America Memoir 133, 213–234. Valentine, J.W., Jablonski, D., 1993. Fossil communities: Compositional varia- tion at many time scales. In: Ricklefs, R.E., Schluter, D. (Eds.), Species Diversity in Ecological Communities: Historical and Geographical Per- spectives. University of Chicago Press, Chicago, pp. 341–349. Vendrasco, M.J., Wood, T.E., Runnegar, T.N., 2004. Articulated Palaeozoic fossil with 17 plates greatly expands disparity of early chitons. Nature 429, 288 – 291.