¡Descarga T5. Impulso nervioso y más Apuntes en PDF de Enzimas y Metabolismo solo en Docsity!
TEMA 5: TRANSMISIÓN DEL IMPULSO NERVIOSO
1. POTENCIALES GRADUADOS
Un cambio cualquiera en el potencial de membrana es un potencial graduado. ¾ Despolarización: cambio de voltaje que eleva el potencial por encima de - 70mV. ¾ Hiperpolarización: cambio de voltaje que disminuye el potencial por debajo de - 70mV ¾ Repolarización: cambio de voltaje restaura el potencial al valor de - 70m Características:
- Intensidad variable
- Distancias cortas
- Excitatorio o inhibitorio
- Si es despolarizante puede desencadenar un potencial de acción
2. POTENCIAL DE ACCIÓN
El potencial de acción se genera en la zona gatillo del axón, donde empiezan los canales dependientes de voltaje (siempre tiene la misma intensidad, el mismo perfil, y la misma duración). Es despolarizante y se propaga largas distancias.
- Potencial de membrana en reposo.
- Estímulo eficaz despolarizante
- Potencial umbral: Los canales de Na+ regulados por voltaje se abren y entra Na+ a la célula. La membrana se despolariza hasta el umbral (- 55 mV).
- Despolarización rápida por entrada de Na+ despolariza la célula hasta +30 mV.
- Amplitud máxima. Los canales de Na+ se inactivan y los canales de K+ dependientes de voltaje más lentos se abren.
- Sale K+ a favor de gradiente y comienza la repolarización.El K+ se mueve desde la célula hacia el líquido extracelular, buscando su potencial de equilibrio.
- Los canales de K+ permanecen abiertos al llegar a - 70 mV (se cierran lentamente), K+ adicional abandona la célula
- Los canales lentos de K+ regulados por voltaje se cierran, sale menos K+ de la célula.
- La célula retorna a la permeabilidad iónica en reposo y vuelve al potencial de membrana en reposo. Las cantidades de Na+ que han entrado y K+ que han salido, son reestablecidas por la actividad de la bomba Na+/K+ 3, 4 y 5 Despolarización
- Repolarización 7 y 8. Hiperpolarización
- Potencial de membrana en reposo 2.1. INACTIVACIÓN DE LOS CANALES DEPENDIENTES DE VOLTAJE Este canal tiene la capacidad de estar abierto, cerrado o inactivo Como podemos ver, en este proceso es fundamental los tres posibles estados del canal de Na+. Es necesario para que el potencial de acción viaje en una sola dirección. Como están inactivos, permite que la despolarización avance a la vez que se repolariza por detrás. Esto se conoce con el nombre de periodo refractario, tiempo que hay desde que una neurona dispara un PA hasta que es capaz de responder a otro estímulo y dispara otro PA, distinguimos:
- Periodo refractario absoluto: los canales de Na están inactivos, la neurona no puede disparar un segundo PA
- Periodo refractario relativo: como el K sigue hiperpolarizando la célula, se podría disparar un segundo potencial de acción pero necesitamos un estímulo más despolarizante que el anterior (los canales de Na no están inactivos)
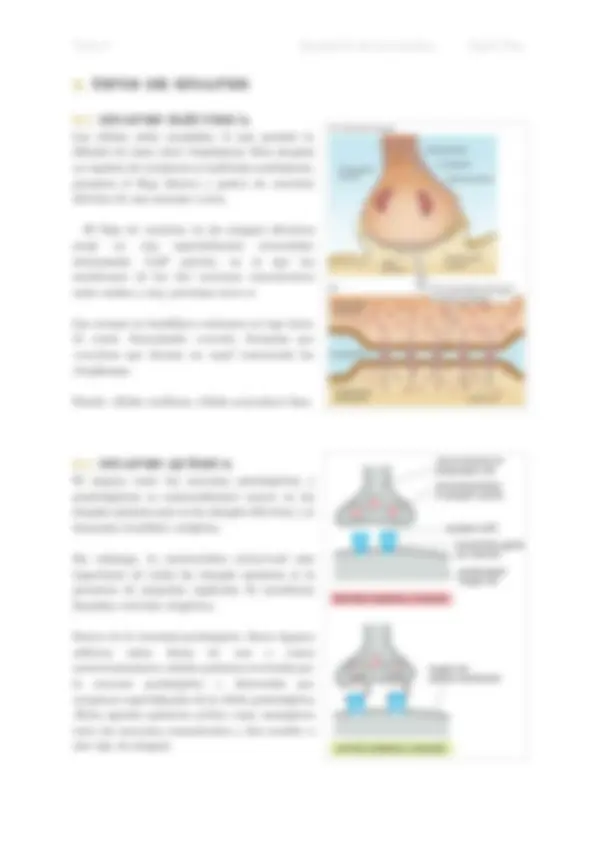
3. TIPOS DE SINAPSIS
3.1. SINAPSIS ELÉCTRICA
Las células están acopladas, lo que permite la difusión de iones entre citoplasmas. Esta sinapsis no requiere de receptores ni moléculas mediadoras, permiten el flujo directo y pasivo de corriente eléctrica de una neurona a otra. El flujo de corriente en las sinapsis eléctricas surge en una especialización intercelular denominada GAP juntion, en la que las membranas de las dos neuronas comunicantes están unidas y muy próximas entre sí. Las uniones en hendidura contienen un tipo único de canal, denominado conexón , formados por conexinas que forman un canal conectando los citoplasmas. Donde: células cardíacas, células musculares lisas.
3.1. SINAPSIS QUÍMICA
El espacio entre las neuronas presinápticas y postsinápticas es sustancialmente mayor en las sinapsis químicas que en las sinapsis eléctricas y se denomina hendidura sináptica. Sin embargo, la característica estructural más importante de todas las sinapsis químicas es la presencia de pequeños orgánulos de membrana llamados vesículas sinápticas. Dentro de la terminal presináptica. Estos órganos esféricos están llenos de uno o varios neurotransmisores, señales químicas secretadas por la neurona presináptica y detectadas por receptores especializados de la célula postsináptica .Estos agentes químicos actúan como mensajeros entre las neuronas comunicantes y dan nombre a este tipo de sinapsis.
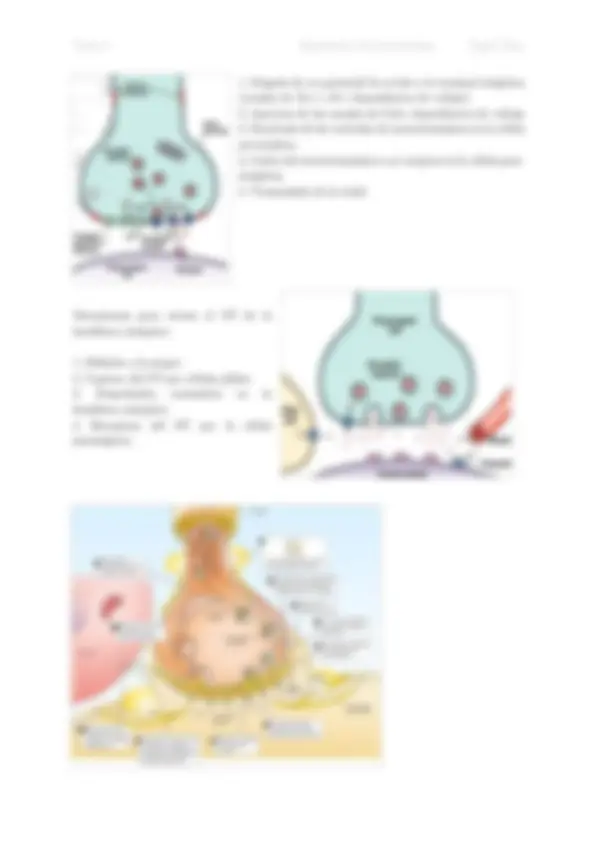
- Llegada de un potencial de acción a la terminal sináptica (canales de Na+ y K+ dependientes de voltaje).
- Apertura de los canales de Ca2+ dependientes de voltaje
- Exocitosis de las vesículas de neurotransmisor en la célula pre-sinática.
- Unión del neurotransmisor a su receptor en la célula post- sináptica.
- Transmisión de la señal. Mecanismos para retirar el NT de la hendidura sináptica:
- Difusión a la sangre
- Captura del NT por células gliales
- Degradación enzimática en la hendidura sináptica
- Recaptura del NT por la célula presináptica
4.1. ACETILCOLINA
La acetilcolina (ACh) fue la primera sustancia identificada como neurotransmisor. Además de su función como neurotransmisor en las uniones neuromusculares esqueléticas, en la sinapsis neuromuscular entre el nervio vago y las fibras musculares cardiacas, la ACh sirve como transmisor en en los ganglios del sistema motor visceral y en diversos lugares del SNC. A. RECEPTOR NICOTÍNICO DE ACETILCOLINA Muchas de las acciones postsinápticas de la ACh están mediadas por el receptor nicotínico de la ACh (nAChR), llamado así porque porque la nicotina estimulante del SNC también se une a estos receptores. Antagonista (muscular): d-tubocurarina / Antagonista (neuronal): α- bungarotoxina En resumen, el receptor nACh es un canal iónico regulado por ligando. La íntima asociación de los sitios de unión de ACh de este receptor con el poro del canal permite la rápida respuesta a ACh. Cuando une a la acetilcolina se abre el poro: Esta disposición general es característica de todos los receptores ionotrópicos de las sinapsis que emplean glutamato, GABA, serotonina y otros neurotransmisores. Así pues, este receptor nictínico sirve para el estudio de otros receptores ionotrópicos. Receptores Ionotrópicos. Receptores Metabotrópicos.
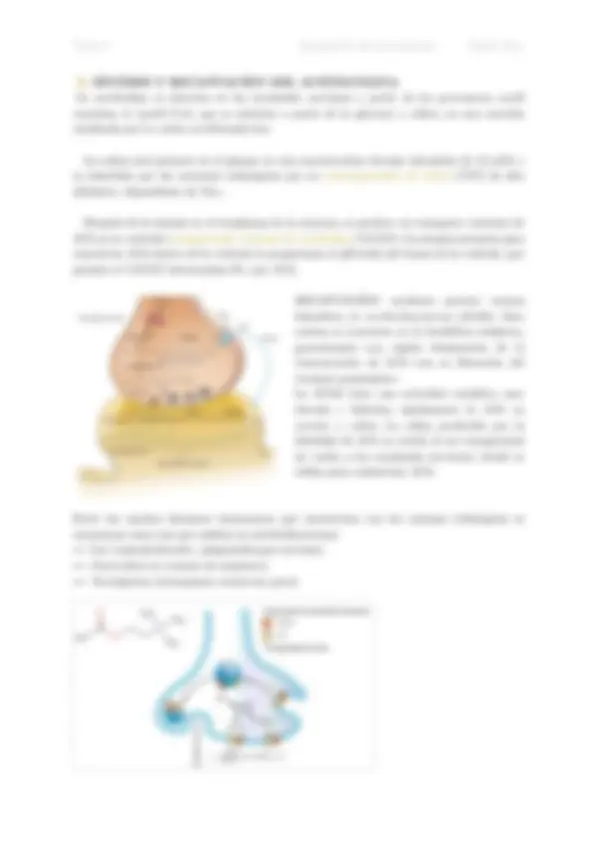
B. SÍNTESIS Y RECAPTACIÓN DEL ACETILCOLINA
La acetilcolina se sintetiza en las terminales nerviosas a partir de los precursores acetil coenzima A (acetil CoA, que se sintetiza a partir de la glucosa) y colina, en una reacción catalizada por la colina acetiltransferasa. La colina está presente en el plasma en una concentración elevada (alrededor de 10 mM) y es absorbida por las neuronas colinérgicas por un cotransportador de colina (ChT) de alta afinidad y dependiente de Na+. Después de la síntesis en el citoplasma de la neurona, se produce un transporte vesicular de ACh en su vesícula (transportador vesicular de acetilcolina VAChT). La energía necesaria para concentrar ACh dentro de la vesícula la proporciona el pH ácido del lumen de la vesícula, que permite al VAChT intercambiar H+ por ACh. RECAPTACIÓN: mediante potente enzima hidrolítica, la acetilcolinesterasa (AChE). Esta enzima se concentra en la hendidura sináptica, garantizando una rápida disminución de la concentración de ACh tras su liberación del terminal presináptico. La AChE tiene una actividad catalítica muy elevada e hidroliza rápidamente la ACh en acetato y colina. La colina producida por la hidrólisis de ACh se recicla al ser transportada de vuelta a las terminales nerviosas, donde se utiliza para resintetizar ACh. Entre los muchos fármacos interesantes que interactúan con las enzimas colinérgicas se encuentran estos tres que inhiben la acetilcolinesterasa: ¾ Los organofosforados. (plaguicidas-gas nervioso). ¾ Fasciculina en (veneno de serpiente). ¾ Neostigmina (tratamiento miastenia gravi)
4.2. GLUTAMATO
El glutamato es el transmisor más importante para el funcionamiento normal del cerebro. Casi todas las neuronas excitadoras del SNC son glutamatérgicas y se calcula que más de la mitad de las sinapsis cerebrales liberan este neurotransmisor. El glutamato es un aminoácido no esencial que no atraviesa la barrera hematoencefálica y, por tanto, debe sintetizarse en las neuronas a partir de precursores locales. El precursor más frecuente para la síntesis de glutamato es la glutamina. A. RECEPTORES DE GLUTAMATO Existen varios tipos de receptores de glutamato ionotrópicos. Los receptores AMPA , NMDA y kainato reciben su nombre de los agonistas que los activan. Todos estos receptores son canales de cationes activados por glutamato que permiten el paso de Na+ y K+ (respuestas excitadoras).
- AMPA (α-amino- 3 - hidroxil- 5 - metil- 4 - isoxazol-propionato). Respuestas postsinápticas excitatorias. Despolarización muy rápida, mucho mayores que las producidas por otros tipos de receptores de glutamato ionotrópicos, por lo que estos receptores son los principales mediadores de la transmisión excitatoria en el cerebro.
- NMDA (N-metil-d-aspartato) Respuestas postsinápticas excitatorias. Despolarización es más lenta y dura más. Flujo de Na+, K+ y Ca2+.
- Kainato: ácido kainico. Respuestas postsinápticas excitatorias. Despolarización más lenta que AMPA. Flujo de Na+, y K+. Otro tipo son metabotrópicos. Se encuentran en células pre y post- sinápticos, dando palsticidad neuronal.
B. SÍNTESIS Y RECAPTURA DEL GLUTAMATO
El precursor más frecuente para la síntesis de glutamato es la glutamina, que es absorbida por las terminales presinápticas por el transportador 2 del sistema A (SAT2) y luego metabolizada a glutamato por la enzima mitocondrial glutaminasa. La glucosa metabolizada por las neuronas también puede utilizarse para sintetizar glutamato por transaminación de 2 - oxoglutarato , un intermediario del ciclo Krebs. El glutamato sintetizado en el citoplasma presináptico es empaquetado en vesículas sinápticas por transportadores vesiculares de glutamato (VGLUTs). RECAPTACIÓN: Una vez liberado, el glutamato es eliminado de la hendidura sináptica por los transportadores de aminoácidos excitadores (EAATs) Los EAAT son una familia de cinco cotransportadores de glutamato. El glutamato que se transporta a las células gliales a través de las EAAT se convierte en glutamina mediante la enzima glutamina sintetasa. A continuación, la glutamina es transportada fuera de las células gliales por un transportador diferente, el transportador 1 del sistema N (SN1), y transportada a los terminales nerviosos a través de SAT. Esta secuencia global de acontecimientos se denomina ciclo glutamato-glutamina
D. PAPEL PROTECTOR DEL LACTATO (EXCITOTOXICIDAD)
El ácido láctico protege a la neurona de esa excitotoxicidad. En condiciones de isquemia no hay O2 y no hay glucosa por lo que las neuronas pueden utilizar lactato la obtención de energía. Este lactato proviene, o de la producción de los eritrocitos (ciclo de cori) que va a estar en contacto con el tejido neuronal, o el que han producido los astrocitos mediante la glucólisis. ¾ Ciclo de Cori. Tiene lugar entre el hígado y otros órganos. El hígado capta el lactato y lo transforma en glucosa mediante la gluconeogénesis. Esta glucosa pasa a la sangre y puede ser utilizada por el eritrocito, que vuelve a generar otra vez lactato mediante glucólisis. También puede ser utilizada por el músculo esquelético en periodos de alta actividad. El lactato se transporta mediante los transportadores de lactato a la matriz mitocondrial, y mediante la acción de la lactato deshidrogenasa pasa a piruvato y a partir de ahí, fosforilación oxidativa produciendo ATP. Este ATP saldrá fuera mediante un canal y se va a unir a los receptores P2Y (receptores de nucleótidos) que activan la PI3K. La PI3K nos va a abrir canales de K+, cuando estos se abren, el K+ tiende a salir por lo que se pierden cargas positivas del interior de la neurona, y se queda más negativa (hiperpolarización). Esta hiperpolarización va en distinto sentido que la despolarización que produce el glutamato por lo que le va a costar un poco más excitarse y esto ayuda a que la neurona no muera.
4. 3. NEUROTRANSMISOR GABA
La mayoría de las sinapsis inhibitorias del cerebro y la médula espinal van a utilizar como neurotransmisores al ácido γ-aminobutírico (GABA) o a la glicina. Ahora se sabe que hasta un tercio de las sinapsis cerebrales utilizan el GABA como neurotransmisor inhibidor. El GABA se encuentra abundantemente en interneuronas del circuito local, aunque también en las neuronas espinosas medias del cuerpo estriado y las células de Purkinje del cerebelo. A. RECEPTORES DE GABA Las sinapsis GABAérgicas emplean dos tipos de receptores postsinápticos, denominados GABA a (ionotrópicos) y GABA b ( metabotrópicos). Los receptores ionotrópicos GABAa son canales aniónicos activados por GABA, siendo el Cl- el principal ion permeante en condiciones fisiológicas. La acción del cotransportador K+/Cl-, mantiene baja la concentración de Cl- intracelular. Así, la activación de estos receptores GABA provoca una afluencia de Cl- cargado negativamente que inhibe las células postsinápticas. Curiosidad: En los casos en los que la concentración postsináptica de Cl- es alta (en neuronas en desarrollo, por ejemplo), los receptores GABA pueden excitar sus dianas postsinápticas. Los receptores metabotropicos GABAb también tienen acción inhibitoria. Pero en este caso la inhibición se debe a menudo a la activación de los canales de K+. (GIRK) La segunda acción de los receptores GABAB es bloquear los canales de Ca2+, lo que también inhibe las células postsinápticas.
Transaminación para la síntesis de GABA PARTES DEL CEREBRO USAN GABA: cerebelo (coordinación de movimientos), hipocampo (formación recuerdos). Célula postsináptica: abre canales de potasio (sale), voltaje más negativo (hiperpolarización). Célula presináptica: inhibe los canales de calcio dependientes de voltaje (impide la liberación de neurotransmisores).
4.4. AMINAS BIÓGENAS
Los transmisores de aminas biógenas regulan muchas funciones cerebrales y también son activos en el sistema nervioso periférico. Dado que las aminas biógenas están implicadas en una variedad tan amplia de comportamientos, los defectos en la función de las aminas biógenas estén implicados en la mayoría de los trastornos psiquiátricos. Existen cinco neurotransmisores de aminas biógenas bien establecidos: las tres catecolaminas - dopamina, noradrenalina y adrenalina- , la histamina y la serotonina. Todas las catecolaminas (llamadas así porque comparten la fracción catecol) derivan de un precursor común, el aminoácido tirosina. El primer paso en la síntesis de catecolaminas es catalizado por la tirosina hidroxilasa en una reacción que requiere oxígeno como co- sustrato y tetrahidrobiopterina como cofactor para sintetizar dihidroxifenilalanina (DOPA).
A. CATECOLAMINAS - DOPAMINA
La dopamina es el neurotransmisor catecolaminérgico más importante del Sistema Nervioso Central (SNC) de los mamíferos y participa en la regulación de diversas funciones como la conducta motora, la emotividad y la afectividad así como en la comunicación neuroendócrina. A.1. CRCUITOS DOPAMINÉRGICOS ¾ Nigroestriatal (coordinación de movimientos) ¾ Mesolimbico-cortical (memoria, emociones y motivación) ¾ Hipotalámico (Secreción hormonal). La dopamina está presente en varias regiones cerebrales, aunque la principal zona del cerebro que contiene dopamina es el cuerpo estriado, que recibe la mayor parte de la sustancia negra y desempeña un papel esencial en la coordinación de los movimientos corporales. También se cree que la dopamina está implicada en la motivación, la recompensa y el refuerzo. Además de estas funciones en el SNC, la dopamina también desempeña un papel poco conocido en algunos ganglios simpáticos.
B. SEROTONINA
Es una amina que se sintetiza a partir de triptófano (aminoácido esencial), en dos pasos nos forma serotonina. Está implicada en sueño-vigilia, emociones, ritmos cardíacos, estado de conciencia, saciedad-ingesta, etc. B.1. RECEPTORES Y PATOLOGÍAS Las alteraciones en la función de estos receptores se han implicado en numerosos trastornos psiquiátricos, como la depresión, los trastornos de ansiedad y la esquizofrenia y los fármacos que actúan sobre los receptores de serotonina son tratamientos eficaces para varias de estas afecciones.. Agonistas: Aumentan la duración de la transmisión serotoninérgica a través de distintos mecanismos. ¾ Inhibidores de la degradación de la Serotonina: hay más serotonina disponible. Ejemplo: Fármacos Antidepresivos IMAO (Inhibidores de MAO). ¾ Inhibidores Selectivos de la Recaptación de Serotonina (ISRS): la Serotonina está más tiempo disponible en la hendidura sináptica. Ejemplo: Fármacos antidepresivos (Prozac). Segundo paso de la degradación por la MAO (5-IHH) es soluble y excretado en la orina, marcador tumoral (producción más alta de serotonina que la que debería).
4. 5. NEUROTRANSMISORES NO CLÁSICOS
A. PURINAS - ATP/ADENOSINA
Todas las vesículas sinápticas contienen ATP, que se libera junto con uno o varios neurotransmisores "clásicos". Esta observación plantea la posibilidad de que el ATP actúe como cotransmisor. El ATP actúa como neurotransmisor excitador en las motoneuronas de la médula espinal, así como en los ganglios sensoriales y autónomos. También se han demostrado acciones postsinápticas del ATP en el SNC. Las enzimas extracelulares degradan el ATP liberado en adenosina, que tiene su propio conjunto de acciones de señalización. Así pues, la adenosina no puede considerarse un neurotransmisor clásico porque no se almacena en vesículas sinápticas ni se libera de forma dependiente del Ca2+. Varias enzimas, como la apirasa , la ecto- 5 ′ nucleotidasa y los transportadores de nucleósidos, participan en el catabolismo rápido y la eliminación de purinas de localizaciones extracelulares. B. OXIDO NITRICO El óxido nítrico (NO) es una señal química inusual. Es un gas producido por la acción de la óxido nítrico sintasa , una enzima que convierte el aminoácido arginina en un metabolito (citrulina) y genera simultáneamente NO. En las neuronas, la NO sintasa está regulada por la unión del Ca2+ a la proteína calmodulina, que es un sensor del Ca2+. Una vez producido, el NO puede permeabilizar la membrana plasmática, lo que significa que el NO generado dentro de una célula puede viajar a través del medio extracelular y actuar dentro de las células cercanas. Así, esta señal gaseosa tiene un rango de influencia que se extiende mucho más allá de la célula de origen, difundiéndose unas decenas de micrómetros desde su lugar de producción antes de ser degradado.