¡Descarga Tema 2. la membrana plasmática. y más Apuntes en PDF de Citología e Histología Vegetal y Animal solo en Docsity!
Tema 2. La Membrana Plasmática.
Contenido:
- Membranas celulares, estructura y funciones. ·Bicapa lipídica: base universal de la estructura de la membrana celular. ·Lípidos de membrana. ·Proteínas de membrana. ·Propiedades de las membranas.
- Transporte de moléculas pequeñas a través de la membrana. ·Principios del transporte a través de membranas. ·Proteínas transportadoras. Transporte activo y transporte pasivo. ·Canales iónicos.
- Transporte de macromoléculas y pequeñas a través de la membrana. ·Exocitosis. ·Endocitosis: pinocitosis y endocitosis mediada por receptores. ·Fagocitosis.
Estructura de la membrana.
Las membranas celulares son cruciales para la vida celular. La membrana plasmática envuelve la célula definiendo sus límites y manteniendo las diferencias esenciales entre su contenido y el entorno. Dentro de la célula eucariota, las membranas del retículo endoplasmático, del complejo de Golgi, de las mitocondrias y de otros orgánulos delimitados por membrana mantienen las diferencias características entre el contenido de cada orgánulo y el citosol. Los gradientes iónicos que se establecen a través de las membranas, generados por la actividad de proteínas de membrana especializadas, pueden utilizarse para sintetizar nerviosas y musculares para producir y transmitir señales eléctricas. En todas las células, la membrana plasmática contiene también proteínas que actúan como sensores de señales externas, permitiendo que la célula cambie su comportamiento en respuesta a indicaciones ambientales, incluyendo señales procedentes de otras células; estas proteínas sensores, o receptores, no transfieren moléculas a través de la membrana sino información. A pesar de que tienen diferentes funciones, todas las membranas biológicas comparten una estructura básica como: una finísima capa de moléculas lipídicas y proteicas, que se mantienen unidas fundamentalmente por interacciones no covalentes. Las membranas celulares son estructuras dinámicas, fluidas y la mayoría de sus moléculas pueden desplazarse en el plano de la membrana. Las moléculas lipídicas están dispuestas en forma de una doble capa continua de
aproximadamente 5nm de grosor. Esta bicapa lipídica constituye la estructura básica de la membrana y actúa de de barrera relativamente impermeable al paso de la mayoría de moléculas hidrosolubles. Las moléculas proteicas que atraviesan la bicapa lipídica son las responsables de la mayoría de las demás funciones de la membrana, por ejemplo transportando determinadas moléculas a través de ella o canalizando reacciones asociadas a membrana, como la síntesis de ATP. En la membrana plasmática algunas proteínas transmembrana actúan de eslabones estructurales que conectan el citoesqueleto, a través de la bicapa lipídicas, con la matriz extracelular o con una célula adyacente. Otras proteínas actúan como receptores que reciben y translucen las señales químicas procedentes del entorno celular. Como podría esperarse, la célula precisa de numerosas proteínas de membrana para poder funcionar e interaccionar con su entorno. De hecho, se estima que el 30% de las proteínas codificadas por el genoma de una célula animal son proteínas de membrana.
La bicapa lipídica.
La bicapa lipídica proporciona la estructura básica de todas las membranas celulares. Es fácil de observar con el microscopio electrónico; su estructura se debe exclusivamente a las propiedades especiales de las moléculas lipídicas que la forman que se ensamblan espontáneamente formando bicapas, incluso bajo condiciones artificiales sencillas.
Los fosfogilicéridos, los esfingolípidos y los esteroles son los
principales lípidos de las membranas celulares.
Las moléculas lipídicas constituyen aproximadamente el 50% de la masa de las membranas de las células animales y casi todo el resto es proteína. En una sección de bicapa lipídica de 1 micrómetro x 1 micrómetro hay 5 x 106 moléculas lipídicas, es decir, unas 109 moléculas lipídicas en la membrana plasmática de una célula animal pequeña. Todas las moléculas lipídicas de las membranas celulares son anfipáticas, es decir, tienen un extremo hidrofílico (“atraído por el agua”) o polar y un extremo hidrofóbico (“que rehuye el agua”) o apolar. Los lípidos de membrana más abundantes son los fosfolípidos. Tienen una cabeza polar y dos colas hidrocarbonadas hidrofóbicas. En las células de los animales, de las plantas y de las bacterias las colas suelen ser ácidos grasos y pueden tener diferente longitud (por lo general contienen de 14 a 24 átomos de carbono). Una de las colas presenta típicamente uno o más dobles enlaces cis (es decir, insaturada) mientras que la otra normalmente no tiene dobles enlaces (es decir, saturada). Cada doble enlace cis genera una suave curvatura en la cadena. Las diferencias de longitud y de grados de saturación entre las colas hidrocarbonadas son importantes
orientan solas en la bicapa con su grupo hidroxilo cerca de los grupos polares de las moléculas adyacentes de los fosfolípidos.
Los fosfolípidos forman bicapas de forma espontánea.
La forma y la naturaleza anfipática de las moléculas de fosfolípido hace que formen espontáneamente bicapas lipídicas en un entorno acuoso. Las moléculas hidrofóbicas se disuelven con facilidad con facilidad en agua, ya que contienen grupos cargados o grupos polares no cargados que les permiten establecer interacciones electrostáticas o enlaces de hidrógeno favorables con las moléculas de agua. Por el contrario, las moléculas hidrofóbicas son insolubles en agua, porque todos o casi todos sus átomos no presentan carga, son no polares, por lo que no pueden formar interacciones favorables energéticamente con las moléculas de agua. Si se dispersan en agua fuerzan a las moléculas de agua adyacentes a reorganizarse alrededor de las moléculas hidrofóbicas formando una especie de entramado similar al hielo. Dado que en estos entramados las moléculas de agua se encuentran más ordenadas que en el agua que las rodea, su formación incrementa la energía libre. Sin embargo, este coste de energía libre se reduce si las moléculas hidrofóbicas se agregan entre sí, reduciendo al mínimo el número de moléculas de agua afectadas. Las regiones hidrofóbicas e hidrofílicas de las moléculas lipídicas se comportan de esta misma forma. Así, las moléculas lipídicas se agregan espontáneamente de manera que sus colas hidrofóbicas hidrocarbonadas se esconden en el interior del agregado y sus cabezas hidrofílicas
queden expuestas al agua. Dependiendo de cual sea su forma, pueden conseguir esta disposición de una de estas dos maneras: pueden formar micelas esféricas, con las colas hacia el interior, o pueden formar láminas de dos capas o bicapas, con las colas hidrofóbicas escondidas entre dos capas de cabezas hidrofílicas. Debido a su forma cilíndrica, en un entorno acuosos la mayoría de los fosfolípidos de membrana forman espontáneamente bicapas. En esta distribución más favorable desde un punto de vista energético, las cabezas hidrofílicas están en contacto con el agua en la superficie de la bicapa, y las colas hidrofóbicas están protegidas del agua en el interior de la estructura. Las mismas fuerzas que impulsan a los fosfolípidos a formar bicapas les confieren propiedades de autosellado. Una pequeña rotura de la bicapa crea un extremo libre en contacto con el agua; los lípidos tienden a reorganizarse de forma espontánea y eliminan este extremo libre energéticamente desfavorable. (En las membranas plasmáticas de las células eucariotas, los grandes “desgarrones” son reparados mediante la fusión de vesículas intracelulares). El hecho de que no puedan existir extremos libres tiene una consecuencia muy importante: la única manera en que una bicapa puede evitar tener extremos libres es cerrándose sobre sí misma y formar un compartimento sellado. Este notable comportamiento, que es fundamental para la creación de una célula viva, es una consecuencia directa de la forma y la naturaleza anfipática de las moléculas de fosfolípido. Además de esta propiedad de autosellado, las bicapas lipídicas tienen otras características que hacen de ellas una estructura ideal para constituir las membranas celulares. Una de las más importantes es su fluidez, que es crucial para funciones de la membrana.
La bicapa lipídica es un fluido bidimensional.
Hacia la década de 1970, los investigadores reconocieron por primera vez que las distintas moléculas lipídicas pueden difundir libremente dentro de las bicapas lipídicas. La demostración
translocasas de fosfolípidos, que catalizan un rápido flip-flop de fosfolípidos desde la monocapa donde han sido sintetizados hasta la monocapa opuesta.
La fluidez de una bicapa lipídica depende de su composición.
La fluidez de las membranas celulares tiene que ser regulada con precisión. Algunos procesos de transporte y algunas actividades enzimáticas, por ejemplo, pueden detenerse cuando la viscosidad de la bicapa se incrementa experimentalmente más allá de un nivel umbral. La fluidez de una bicapa lipídica depende tanto de su composición como de su temperatura, como se ha demostrado por estudios en bicapas sintéticas. Una bicapa sintética, producida a partir de un único tipo de fosfolípido, pasa de un estado líquido a un estado cristalino rígido en un punto de congelación característico. Este cambio de estado recibe el nombre de transición de fase; la temperatura a la que se produce es más baja (es decir, la membrana resulta más difícil de congelar) si las cadenas hidrocarbonadas son cortas o tienen dobles enlaces cis. Una menor longitud de la cadena reduce la tendencia de las colas hidrocarbonadas a interaccionar entre sí, tanto entre fosfolípidos de la misma capa como de la capa opuesta, y los dobles enlaces cis producen pliegues en las cadenas hidrocarbonadas que dificultan su empaquetamiento, de forma que las membranas permanecen fluidas a temperaturas bajas. Bacterias, levaduras y otros organismos cuyas temperaturas fluctúan con la de su entorno, controlan la composición de los ácidos grasos de sus lípidos de membrana manteniendo una fluidez relativamente constante. En caso de que la temperatura disminuya, las células de estos organismos sintetizan ácidos grasos con más dobles enlaces cis, de manera que evitan una perdida de fluidez de la bicapa por efecto de la disminución de temperatura.
El colesterol modula las propiedades de las bicapas lipídicas. Cuando se mezcla con fosfolípidos incrementa las propiedades de barrera permeable de la bicapa lipídica. Se inserta en la bicapa con su grupo hidroxilo próximo a las cabezas polares de las moléculas de fosfolípidos de forma que su anillo esteroideo, plano y rígido, interactúan con -y en parte inmoviliza- las regiones de las cabezas hidrocarbonadas que están más próximas a los grupos polares de la cabeza. El colesterol disminuye la movilidad de los primeros grupos CH2 de las cadenas hidrocarbonadas de las moléculas de fosfolípidos; este hecho provoca que la bicapa lipídica se amenos deformable en esa región y se reduzcan su permeabilidad a pequeñas moléculas solubles en agua. A pesar de que el colesterol condensa el empaquetamineto de los lípidos de una bicapa, no hace que la membrana sea menos fluida. A las elevadas concentraciones a las que se presenta en la mayoría de las membranas plasmáticas de las células eucariotas, el colesterol también impide que las cadenas hidrocarbonadas se junten y cristalicen.
A pesar de su fluidez, las bicapas lipídicas
pueden formar dominios de composición diferente.
Dado que una bicapa lipídica es un fluido bidimensional, podríamos esperar que la mayoría de los diferentes tipos de moléculas lipídicas presentes en ella se encontraban distribuidas al azar, mezcladas unas con otras en cada monocapa. Las fuerzas de atracción de Van der Waals entre las colas hidrocarbonadas adyacentes no son los bastante selectivas para mantener unidos grupos de moléculas de fosfolípidos. Sin embargo, con ciertas mezclas lipídicas, diferentes moléculas lipídicas pueden permanecer juntas transitoriamente generando una mezcla dinámica de parches de diferentes dominio. En bicapas lipídicas sintéticas compuestas de fosfatidilcolina, esfingomielina y colesterol, las fuerzas de van der Waals entre las largas cadenas hidrocarbonadas saturadas de las moléculas de esfingomielina podrían ser bastantes fuertes para mantener unidas de forma temporal a las moléculas adyacentes. Se ha producido un largo debate entre científico sobre si las moléculas lipídicas de la membrana plasmáticas de las células animales pueden ensamblarse transitoriamente formado dominios especializados denominados balsas lipídicas. Ciertas regiones especializadas de la membrana plasmática como es el caso de las caveolas que participan en la endocitosis están enriquecidas en esfingolípidos y colesterol y parece que las proteínas especificas que se ensamblan en ellos pueden ayudar a estabilizar estas balsas. Dado que las cadenas hidrocarbonadas de los esfingolípidos son más largas y más rectas que las de los otros lípidos de membrana, los
La asimetría de la bicapa lipídica es importante para su función.
En muchas membranas, la composición lipídica de las dos monocapas de la bicapa lipídica es marcadamente diferente entre sí. Por ejemplo, en la membrana del glóbulo rojo humano en la monocapa exterior de la bicapa lipídica se encuentra la mayoría de las moléculas de fosfolípidos que tienen colina en su grupo cabeza, mientras que en la monocapa interior se encuentran la mayoría que contienen un grupo amino primario terminal. Así, la fosfatidilserina, de carga negativa, está localizada en la monocapa interior, por lo que existe una importante diferencia de carga entre las dos mitades de la bicapa. La asimetría de los lípidos es importante desde el punto de vista funcional, especialmente para transformar las señales extracelulares en señales intracelulares. Muchas proteínas citosólicas se unen a un grupo de cabeza de un lípido determinado en la monocapa citosólica de la bicapa lipídica.
En la superficie de todas las
membranas plasmáticas hay glucolípidos.
Las moléculas lipídicas que contienen azúcares, denominadas glucolípidos, son las que presentan mayor asimetría en cuanto a su distribución en las membranas celulares, ya que solo se encuentran en la monocapa no citosólica de la bicapa lipídica. En las células animales se sintetizan a partir de esfingosina, exactamente igual que la esfingomielina. Estas fascinantes moléculas tienden a autoasociarse en parte mediante la formación de enlaces de hidrógeno entre sus azúcares y en parte mediante fuerzas de van der Waals entre sus largas y rectas cadenas hidrocarbonadas, y preferentemente forman parte de balsas lipídicas. La distribución asimétrica de los glucolípidos en la bicapa en consecuencia de la adición de azúcares a las moléculas lipídicas en el lumen del complejo de Golgi. Así, el compartimento en el que son sintetizadas es equivalente desde un punto de vista topológico al exterior celular. Cuando son descargas en la membrana plasmática, los grupos azúcar quedan expuestos a la superficie celular, donde ejercen funciones importantes en las interacciones de la collas con su entorno. Los glucolípidos probablemente se presentan en las membranas plasmáticas de todas las células animales, constituyendo alrededor de un 5% de las moléculas lipídicas de la monocapa exterior. Además se encuentran en algunas membranas intracelulares. Los glucolípidos más complejos, los gangliósidos, contienen oligosacáridos, con uno o más residuos de ácido siálico, lo que les
proporciona una carga neta negativa. El más abundante de los más de 40 gangliósidos diferentes que se han identificado se encuentra en la membrana plasmática de las células nerviosas, donde constituyen cerca de un 5-10% de la masa lipídica total; también pueden encontrarse en cantidades muchos menores en otros tipos celulares. Los indicios acerca de las funciones de los glucolípidos proceden de su localización. Por ejemplo, en la membrana plasmática de las células epiteliales, los glucolípidos se encuentran confinados en la superficie apical, donde podrían ayudar a proteger a la membrana de las condiciones adversas que con frecuencia existen allí. los glucolípidos con carga, como los gangliósidos, pueden tener importancia debido a sus efectos eléctricos: su presencia alteraría el campo eléctrico a través de la membrana y la concentración de iones -especialmente Ca2+- en la superficie de la membrana. Parece que los glucolípidos participan en los procesos de renacimiento celular, en los que las proteínas de membrana de unión a carbohidratos (lectinas) se unen a los azúcares tanto de glucolípidos como de glucoproteínas, en el proceso de adhesión célula-célula. Sin embargo, sorprendentemente, ratones mutantes deficientes en todos sus gangliósidos complejos no presentan anormalidades evidentes, aunque se ha observado que los machos no pueden transportar de forma correcta la testosterona en los testículos.
Proteínas de membrana.
Aunque la estructura básica de las membranas biológicas está determinada por la bicapa lipídica, la mayoría de sus funciones especificas están desempeñadas por proteínas. Así pues, son las proteínas las que confieren a cada uno de los tipos de membrana celular sus propiedades funcionales características. De acuerdo con este hecho, la cantidad y el tipo de proteínas de una membrana son muy variables. En la vaina de mielina, cuya función principal consiste en aislar los axones nerviosos, la proteína constituye menos de un 25% de la masa de la membrana. Por el contrario, en las membranas implicadas en la producción de ATP aproximadamente un 75% de su masa es proteína. Una membrana plasmática normal está situada entre ambos extremos, siendo las proteínas cerca de la mitad de su masa. Debido a que las moléculas lipídicas son pequeñas en comparación con las proteicas, en todas las membranas celulares hay más moléculas lipídicas que proteicas -una membrana que contenga un 50% de proteína en masa tiene una proporción aproximada de 50 moléculas de lípidos por cada molécula de proteína. Las proteínas de membrana son muy variables en cuanto a estructura y en cuanto a la manera en la que se asocian con la bicapa lipídica, lo cual refleja la variedad de sus funciones.
denomina proteínas periféricas de membrana. Por el contrario, las proteínas transmembrana y muchas proteínas unidas a la bicapa por cadenas de ácidos grasos o por ejemplo polipeptídicas hidrofóbicas que se insertan en el corazón hidrofóbico de la bicapa lipídica no pueden ser liberadas por estos métodos, por lo que se les denomina proteínas integrales de membrana.
Los anclajes lipídicos controlan la localización en la membrana de
algunas proteínas señalizadoras.
Por lo general, la manera en que una prometían de membrana se asocia a la bicapa lipídica indica su función. Así, sólo las proteínas transmembrana pueden actuar en ambos lados de la bicapa o transportar moléculas a través de ella. Por ejemplo, algunos receptores celulares de superficie son proteínas transmembrana que se unen a las moléculas señal en el espacio extracelular y generan diferentes señales intracelulares en el lado opuesto de la membrana plasmática. Para transferir pequeñas moléculas hidrofílicas a través de la membrana, una prometían de transporte puede proporcionar un camino para que la molécula atraviese la barrera de la bicapa lipídica; la arquitectura molecular de las proteínas multipaso a través de la membrana resulta ideal para conseguir esta función. Por el contrario, las proteínas que sólo actúan en un lado de la bicapa lipídica generalmente están asociadas a la monocapa lipídica o al dominio proteico de aquel lado de la membrana. por ejemplo, algunas de las proteínas relacionadas con la señalización intracelular, que participan en la transformación de señales extracelulares en señales intracelulares, están unidas a la cara citosólica de la membrana plasmática por uno o más grupos lipídicos a los que están unidos de forma covalente. Estos grupos pueden ser ácidos ácidos grasos o grupos prenilo. En algunos casos, el ácido mirístico, un ácido graso saturado de 14 carbonos, se añade al grupo amino
terminal de la proteína durante su síntesis en el ribosoma. Todos los miembros de la familia Src de las proteínas tirosina quinasa citoplasmáticas están miristoiladas de esta forma. Sin embargo, la unión a la membrana a través de un solo anclaje lipídico no es muy fuerte y, a menudo se añade un segundo grupo lipídico para anclar las proteínas con más firmeza a la membrana. En el caso de la mayoría de las quinasas Src, la segundo modificación lipídica consiste en la unión de ácido palmítico, un ácido graso saturado de 16 carbonos, a la cadena lateral de una cisteína de la proteína. Esta modificación ocurre en respuesta a una señal extracelular y ayuda a reclutar las quinasas hacia la membrana plasmática. Cuando la vía señalizada se apaga, el ácido palmítico es eliminado de la prometían y la quinasa vuelve al citosol. Otras proteínas señalizadoras intracelulares, como las pequeñas GTPasas de la familia Raf utilizan una combinación de unión a g r u p o p r e n i l o y á c i d o palmítico para reclutar las proteínas en la membrana plasmática.
En la mayoría de las
proteínas
transmembrana, las regiones de la cadena polipeptídica que
atraviesan la bicapa lipídica presentan una conformación en hélice
alfa.
Una proteína transmembrana se orienta siempre en un único sentido dentro de las membranas. Este hecho refleja tanto la asimetría del sistema mediante el cual la proteína ha sido insertada en la bicapa lipídica del ER durante su biosíntesis, como la diferencia entre las funciones que desempeñan los dominios citoplasmáticos y no citoplasmáticos de la proteína. Estos dominios están separados por segmentos de la cadena polipeptídica que atraviesan la membrana, se hallan en contacto con el ambiente hidrofóbico de la bicapa lipídica y están formado, en gran parte, por aminoácidos de cadena lateral no polar. Debido a que las uniones peptídicas, son, en sí mismas, polares y dado que el agua no está presente en este ambiente, todas las uniones peptídicas pertenecientes a la porción de la cadena embebida en la bicapa tienden a formar puentes de hidrógeno entre eso. El número de este tipo de uniones se maximiza si la cadena polipeptídica forma una hélice alfa regular al cruza la bicapa; en efecto, así es como la mayoría de los segmentos transmembrana de las cadenas polipeptídicas atraviesan la membrana.
A menudo las hélices alfa transmembrana interactúan entre sí.
Las hélices alfa transmembrana de muchas proteínas de un solo paso no contribuyen al plegamiento de los dominios de la proteína a cada lado de la membrana. Como consecuencia de ello, a menudo es posible modificar de forma artificial a las células para conseguir que produzcan los dominios citosólicos o extracelular de estas proteínas en forma de proteína soluble en agua. Esta proteína no ha servido para estudiar la estructura y la función de estos dominios, especialmente en el caso de las proteínas receptoras transmembrana. Sin embargo, una hélice alfa transmembrana, incluso en una proteína de un solo paso, no hace más que anclar la proteína en la bicapa lipídica. Muchas proteínas de un solo paso, a través de membrana forman homodímeros, que se mantienen unidos mediante fuertes interacciones especificas entre las dos hélices alfa transmembrana; la secuencia de los aminoácidos hidrofóbicos de estas hélices contienen la información que dirige la interacción proteína proteína.
De forma similar, las hélices alfa transmembrana de las proteínas multipaso ocupan posiciones específicas en la estructura plegada de una proteína que está determinada por interacciones entre las hélices vecinas. Estas interacciones son cruciales para la estructura y la función de muchos canales y transportadores que desplazan moléculas a través de la bicapa lipídica. En muchos casos, podemos utilizan proteasas para cortar las asas de cadena polipeptídica que, a cada lado de la membrana, unen las hélices alfa transmembrana; estas hélices permanecen unidas y funcionan con normalidad. En algunos casos, incluso podemos expresar en células vivas genes modificados artificialmente que codifican piezas de proteínas multipaso, y se constata que estas piezas separadas se ensamblan de forma adecuada formando una proteína transmembrana funcional, lo cual pone de manifiesto la exquisita especificidad con la que interactúan entre sí las hélices alfa transmembrana.
Algunos barriles ß forman grande canales transmembrana.
Algunas proteínas transmembranas de paso múltiple cuyos segmentos transmembrana están plegados como barriles ß son considerablemente rígidos y tienden a cristalizar con facilidad. Así, algunas de ellas fueron las primeras proteínas transmembrana de paso múltiple que se determinaron mediante cristalografía de rayos X. El número de hebras ß de un barril ß es muy variable, y oscila desde 9 hasta 22. Las proteínas en barril ß son abundantes en la membrana mitocondrial externa, en cloroplastos y en muchas especies de bacterias. Algunas son proteínas formadoras de poros, las cuales generan canales llenos de agua que permiten que algunas pequeñas moléculas hidrofílicas atraviesen de forma especifica la bicapa lipídica de la membrana externa bacteriana. Las porinas son ejemplos bien estudiados. El barril de las porinas está formado por una lamina ß de 16 cadenas antiparalelas y es suficientemente grande como para organizarse en forma de una estructura cilíndrica. Los aminoácidos cuyas cadenas laterales son polares se alinean en el interior del canal acuoso, mientras que las cadenas laterales polares se sitúan en el exterior, Interactuando con el núcleo hidrofóbico de la bicapa lipídica. Con frecuencia algunos bucles de la cadena polipeptídica se proyectan hacia el lumen del canal, estrechándolo de forma que sólo algunos solutos pueden atravesarlo. Así pues, algunas porinas son muy selectivas: por ejemplo, la maltoporina permite el paso preferentemente de maltosa y de sus oligómeros a través de la membrana de E.Coli. La prometían FepA es un ejemplo más complejo de una prometían de transporte de este tipo. Transporta iones hierro a través de la membrana bacteriana externa. Su gran barril está formado por 22 hebras ß y un gran dominio globular que llena por completo el interior del barril. Los iones hierro se unen a este dominio, que al parece realiza un gran cambio conformacional transportando el ion hierro a través de la membrana.
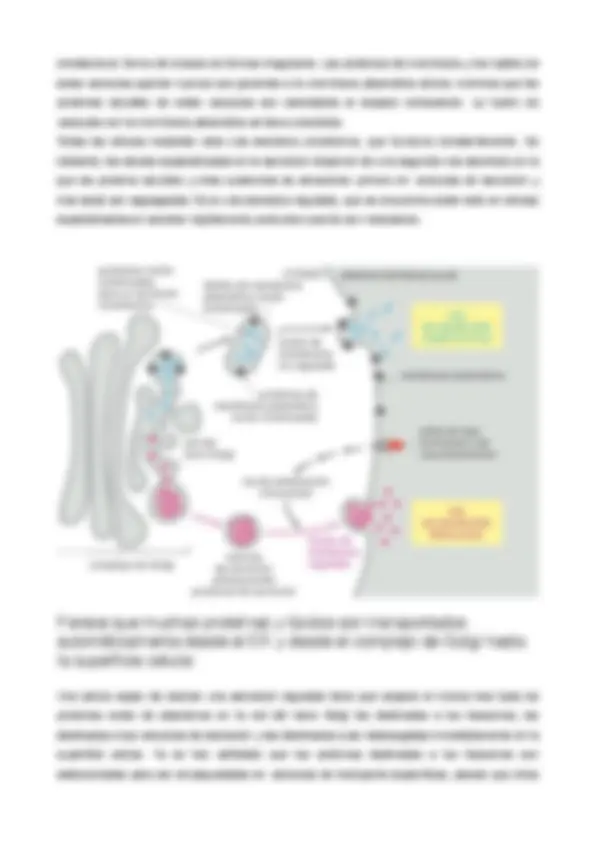
algunos proteoglucanos, el núcleo proteico se extiende a través de la bicapa lipídica o está anclado a la bicapa mediante un anclaje glucosilfosfatidilnositol (GPI). El término cubierta celular o glucocálix se utiliza a menudo para describir la zona de la superficie celular rica en carbohidratos. Esta capa de carbohidratos puede ser visualizada por medio de diversos colorantes, como el rojo de rutenio, o también gracias a su afinidad a viertas proteínas, las llamadas lectinas, que se unen a carbohidratos y que pueden ser marcadas con una tinción fluorescente u otro marcador visible. A pesar de que la mayoría de los carbohidratos están unidas a moléculas intrínsecas de la membrana plasmática, la capa de carbohidratos contiene además glucoproteínas y proteoglucanos que han sido secretos al espacio extracelular y que luego son absorbidos en la superficie celular. Muchas de estas macromoléculas absorbidas son componentes de la matriz extracelular es sólo una cuestión semántica. Una de las funciones probables de la cubierta celular es proteger las células ante agresiones químicas y mecánicas, y mantener a distancia tanto objetos extraños como otras células para evitar interacciones proteína- proteína indeseables. Las cadenas laterales de oligosacáridos de las glucoproteínas y de glucolípidos son muy diversas en cuanto a la organización de sus azúcares. A pesar de que habitualmente contienen menos de 15 residuos glucídicos, éstos suelen estar ramificados y los azúcares pueden estar unidos entre sí mediante diversos enlaces covalente -a diferencia de los residuos de aminoácidos de una cadena polipeptídica, que están todos unidos por uniones peptídicas idénticas. Tres residuos glucídicos pueden formar cientos de trisacáridos diferentes. En principio, la diversidad y la posición expuesta de los oligosacáridos en la superficie celular les hace en especial indicados para participar en procesos de renacimiento celular. Durante muchos años han existido pocas evidencias de estas sospechada función. Sin embargo, recientemente, se ha determinado que las lectinas unidas a la membrana plasmática reconocen de forma especifica algunos oligosacáridos de glucolípidos de la superficie celular y glucoproteínas. -' "“
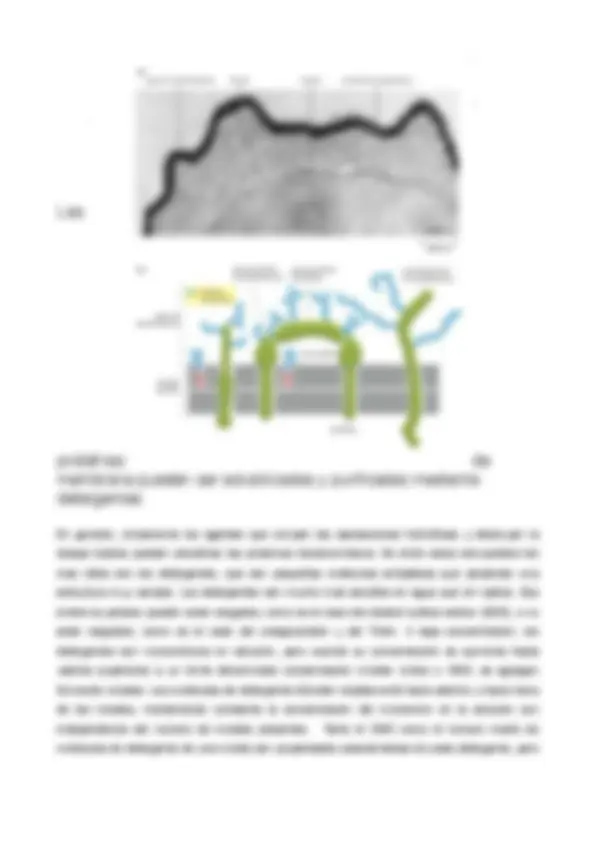
Las
proteínas de
membrana pueden ser solubilizadas y purificadas mediante
detergentes.
En general, únicamente los agentes que rompen las asociaciones hidrofílicas y destruyen la bicapa lipídica pueden solubilizar las proteínas transmembrana. De entre estos compuestos los más útiles son los detergentes, que son pequeñas moléculas anfipáticas que presentan una estructura muy variada. Los detergentes son mucho más solubles en agua que en lípidos. Sus extremos polares pueden estar cargados, como es el caso del dodecil sulfato sódico (SDS), o no estar cargados, como es el caso del octoglucósido y del Tritón. A baja concentración, los detergentes son monoméricos en solución, pero cuando su concentración se aumenta hasta valores superiores a un límite denominado concentración micelar crítica o CMC, se agregan formando micelas. Las moléculas de detergente difunden rápidamente hacia adentro y hacia fuera de las micelas, manteniendo constante la concentración del monómero en la solución con independencia del número de micelas presentes. Tanto el CMC como el número medio de moléculas de detergente de una micela son propiedades características de cada detergente, pero