¡Descarga TEMA 8. INMUNODEFICIENCIAS y más Apuntes en PDF de Inmunología solo en Docsity!
TEMA 8: INMUNODEFICIENCIAS
Defectos en uno o más componentes del SI. Dos tipos:
- Congénitas (o primarias): asociadas a defectos genéticos
- Adquiridas (o secundarias): asociadas a malnutrición, cáncer, tratamiento con inmunosupresores, infecciones de células del sistema inmunitario) Consecuencia principal: mayor susceptibilidad a infecciones La naturaleza de la infección depende del componente del sistema inmunitario afectado Pacientes con inmunodeficiencias son más susceptibles a ciertos tipos de cáncer (virus oncogénicos, EBV) Ciertas inmunodeficiencias están asociadas con una mayor incidencia de autoinmunidad (mecanismo desconocido) Las inmunodeficiencias se producen por:
- Defectos en la diferenciación de linfocitos T o B
- Defectos en la activación de linfocitos T o B
- Defectos en los mecanismos efectores de la inmunidad innata y adaptativa
INMUNODEFICIENCIAS CONGÉNITAS:
Defectos en la inmunidad innata:
- Defectos en fagocitos: infecciones bacterianas y fúngicas en piel y vías respiratorias
- Defectos en señalización por TLRs (detectan componentes no proteicos de bacterias y virus) e interferones tipo I: infecciones piogénicas recurrentes e infecciones víricas
- Defectos en IL12 y vía de IFNg : infecciones intracelulares, especialmente micobacterias Enfermedad granulomatosa crónica:
- Mutaciones en phox
- Producción defectiva de superóxido
- Deficiente eliminación de microbios fagocitados Es un fallo en células fagocíticas, neutrófilos y macrófagos, que provoca la formación de un granuloma porque cuando el neutrófilo o macrófago fagocita un microorganismo va a ser incapaz de eliminarla y esto va a provocar que sobreviva en su interior dando lugar a una inflamación granulomatosa. El neutrófilo con lisosomas cargados con ROS fagocita una bacteria y se produce una fusión de los lisosomas con el fagosoma, la liberación de ROS y la eliminación de la bacteria. Si no hay presencia de ROS en los lisosomas la bacteria no se elimina y sobrevive en el interior del fagocito originando la formación de granulomas.
Diferencias en adhesión leucocitaria Existen varios tipos dependiendo de cuál sea la etapa del proceso de migración de los leucocitos al tejido infectado. Hay un deficiente reclutamiento de leucocitos (mayoritariamente neutrófilos) a sitios de infección
- LAD-1: deficiente expresión de b2-integrinas. Falta de adhesión de los leucocitos al endotelio. La adhesión está mediada por la interacción entre las integrinas expresadas en la superfície de los leucocitos con las moléculas de adhesión del endotelio tipo ICAM. Mutaciones o expresión deficiente de esta b 2 - integrina va a provocar que este proceso de adhesión sea deficiente y por tanto el leucocito no pueda migrar a través del endotelio.
- LAD-2: deficiencia en sialyl Lewis X debido a mutaciones en el transportador de GDP- fucosa. Afecta a la fase de rolling que se produce por la interacción de unas moléculas de carbohidrato presentes en la superfície de los leucocitos como sialyl Lewis X con las selectinas presentes en el endotelio. Esta primera interacción favorece el rolling y la activación del linfocito y la unión al endotelio. Una deficiencia en sialyl Lewis X que media este rolling debido a mutaciones en el transportador de GDP-fucosa afecta a este proceso.
- LAD-3: mutaciones en KINDLIN-3 y deficiente activación de integrinas requerida para la unión de neutrófilos al endotelio. El proceso afectado es la activación de los leucocitos en el proceso de migración al tejido infectado y está asociado a mutaciones en la proteína KINDLIN-3 que participa en la activación de las integrinas para la unión de los neutrófilos o otros leucocitos al endotelio. Afectaciones en cualqueira de estos procesos tiene la misma consecuéncia: los leucocitos no pueden llegar al tejido infectado y las infecciones no pueden ser resuletas.
Defectos en células NK y otros leucocitos: síndroma de Chédiak-Higashi
- Autosómica recesiva
- Infecciones recurrentes, albinismo parcial, infiltración de linfocitos en varios órganos
- Mutaciones en LYST , fusión defectiva de fagosomas y lisosomas en macrófagos, neutrófilos, células NK, linfocitos CTL, melanocitos y plaquetas Situación normal: tenemos un neutrófilo, plaqueta, NK o linfo CTL cargado con lisosomas que cuando se fusiona al fagosoma de forma eficiente elimina el microbio fagocitado. LYST disfuncional: Cuando el neutrófilo, plaqueta, NK o linfo CTL no puede eliminar el microbio se crean unos gránulos gigantes no funcionales y esto conlleva una reducida actividad bactericida y una quimiotaxis deficiente. Defectos en TLRs y vías de señalización NF-kB e INF tipo I
- Mutaciones en MYD88, IRAK4, TLR3, IKKgamma Los TLRs son receptores de componentes que forman parte de bacterias y virus que pueden estar tanto en la membrana (TLR1, 2, 5, 6, 4) y pueden reconocer diferentes tipos de components. También pueden ser intracelulares como el TLR7, 9, 3. El TLR4 reconoce polisacáridos LPS de bacterias gram-. TLR1,2,5,8 reconocen peptidoglicanos. También pueden ser receptores de ácidos nucleicos intracelulares cuando el virus inyecta su DNA en la célula y este DNA foráneo es captado por los TLR 7,9,3. Esto señaliza a la célula que ha sido infectada de manera que es capaz de responder con mecanismos antivirales o antibacterianos. La señalización de estos TLRs ocurre mediante MYD88, IRAK que inducen la activación del factor de transcripción NF-kB que va a ir al nucleo y va a inducir la expresión de una serie de genes como citocinas, moléculas de adhesión o estimuladoras… que van a favorecer la respuesta inmunitaria.
Deficiencia en factores inhibidores H e I
- Síndrome hemolítico urémico
- Glomerulonefritis mebrano-proliferativa
- Infecciones recurrentes
- Fallo en la formación de MAC Tanto en la vía clásica como en la alternativa de activación del complemento hay un paso crítico que es la conversión de C3 en C3b que va a iniciar toda la cascada de activación y de la formación del complejo de ataque a la membrana, MAC que es el que provoca la lisis celular. La conversión de C3 a C3b y la activación de la cascada esta regulada por un feedback positivo que a su vez está controlado por el factor inhibidor H y el factor inhibidor I de manera que déficits en estos dos factores provocan que estos feedbacks positivos esten sobreactivados de manera que todo el C3 que se convierte en C3b sea siempre segradado por la C3-convertasa a C3bBb, de esta manera nunca llega a activar el complemento y por tanto va a haber un déficit en la lisis de células infectadas.
Inmunodeficiencias severas combinadas
- Inmunodeficiencias combinadas o Inmunodeficiencias congénitas que afectan a la inmunidad humoral y cellular
- Inmunodeficiencias combinadas severas (SCID) o Inmunodeficiencias combinadas en las que los linfocitos T periféricos no están presentes of son defectuosos SCID: mutaciones en genes que regulan el desarrollo de linfocitos T y pueden estar acompañadas o no de fallos en la maduración de linfocitos B. El 50% de SCIDs son autosómicas recesivas, 50% de SCIDs asociadas al cromosoma X. De la célula madre hematopoiética se genera CLP que da lugar a una célula Pro B o Pro T, en ambos casos uno de los primeros eventos que ocurre para su desarrollo es la recombinación VDJ en los receptores B (genes Ig) o T (TCR). Este proceso está regulado por una serie de recombinasas y genes que participan en la reparación de roturas de doble cadena de DNA. Mutaciones en estas proteínas encargadas de la reparación de DNA estan asociadas a inmunodeficiencias impidiendo que las células pasen a Pre B o Pre T. En el caso de que pasen a Pre B hay otro checkpoint después regulado por BTK que regula el paso a células B inmaduras que van a medurar expresando receptores maduros en superfície. Fallos en esta BTK también estan asociados a inmunodeficiencias. En el caso de que pasen a Pre T hay otro checkpoint controlado por CD3 para pasar a células CD4+, CD8+ o Tcells. Mutaciones en CD3 también van a provocar defectos asociados a inmunodeficiencias severas combinadas.
SCID ligada al cromosoma X
- Mutaciones en el gen que codifica la cadena gamma común (gamma c) de los receptores de IL2, IL4, IL7, IL9 e IL15 que forma parte del receptor.
- Maduración defectuosa de células T y NK (pero número normal de células B)
- Deficiencia en respuesta humoral causada por la falta de células Th (no producción de AB) Deficiencias en la cadena gamma de todas estas IL provoca una señalización deficiente por los receptores y estas IL son importantes para la diferenciación y funcionamiento del sistem ainmunitario. Ex: Si no hay receptor de IL-7 porque falla la cadena gamma común provoca que no hay linfocitos T. Si no hay receptor de IL-15 no hay células NK. Las células B se pueden desarrollar normalmente pero hay una deficiencia en la respuesta humoral (sin producción de AC) porque las células B no van a poder ser activadas por las células T. SCID asociada a defects en recombinación VDJ
- Mutaciones en genes implicados en recombinación VDJ
- Falta de linfocitos B y T La recombinación VDJ de los genes de las Ig para formar el TCR o el receptor de la célula B es esencial para el desarrollo de los linfocitos T y B. Fallos en proteínas que regulan este proceso van a provocar deficiencias en linfos B y T. Algunas de estas proteínas son: RAG1, RAG2, ARTEMIS, CERNUNNOS, DNA ligasa 4, DNAPKcs, CD45 y CD3 (T), BTK (B).
Síndrome del linfocito desnudo
- Inmunodeficiencia autsómica recesiva
- Falta de expresión de MHC II (HLA-DR, DQ, DP) en células B, macrófagos y células dendríticas (APCs)
- Mutaciones en factores de transcripción clave en la regulación de la expresión de los genes de MHC II: RFX5 o CIITA
- No activación de células T CD4+ por APCs y, por tanto, selección positiva de células T en el timo deficiente Si estas células no tienen MHC II en su superfície no van a poder activar a las células T y por tanto va a haver una falta de activación linfocitaria y una falta de selección positiva de estas células en el timo. RFX5: regula la expresión de los genes de MHC II porque se une a la caja de promotores de los genes de MHCII, que es la caja X, y regula la expresión constitutiva basal de estos genes. CIITA: master regulator de los genes de MHC II y funciona en la expresión de estos genes inducida por INF-gamma. Mutaciones en estos dos genes van a provocar defectos en la expresión constitutiva basal de estos genes y en su expresión inducida por INF-gamma.
- Menos comunes: deficiencias en subclases específicas de IgG como la ausencia de IgG y IgG3, IgG2, IgG4 o IgG3. Defectos en la diferenciación de células B: Inmunodeficiencia común variable
- Grupo heterogéneo de anomalías caracterizadas por niveles bajos de Ig en el suero, reducida respuesta de Ab frente a infecciones o vacunas y mayor incidencia a infecciones
- Se asocian con infecciones piogénicas, pero también con enfermedades autoinmunitarias (anemia perniciosa, anemia hemolítca, artritis reumatoide, IBD) y linfomas
- Bloqueo en la diferenciación a células plasmáticas y reducción de AC de memoria.
- Asociadas a mutaciones en: ICOS : receptor que se expresa en células T y es el receptor de ICOS que se expresa en células B. La iteracción entre célula B y T mediante este receptor provoca la secreción de citocinas tipo IL10 o IL17 del linfocito T que va a activar a la célula B diferenciándola a pasmática y secretando AC. También participa en la generación de memoria en las células B. Las deficiencias en ICOS provocan un déficit en la producción de células B de memoria, menos linfos B en la periféria, déficit del cambio de isotipo de Ig mediado por la interacción con celulas T. En este caso la mutación está en la célula T que no es capaz de activar la célula B. TAC 1 : receptor de BAFF y APRIL que estan implicadas en la diferenciación de células B a plasmáticas y a células B de memoria. Con una mutación vamos a ver euna reducción en las células B de memoria con infecciones recurrentes, déficit en el cambio de isotipo mediado por las células dendríticas, autoinmunidades... CD19: receptor que se expresa en la superfície de células B importante en la activación de células B en respuesta al AG y una falta de activación lleva unos defectos en la diferenciación a células de memoria y plasmáticas impidiendo la generación de AC.
Defectos en la activación de células B dependientes de células T: síndroma de hiper-IgM
- Ligada al cromosoma X en Xq26 donde reside el ligando de CD
- Mutaciones en CD40L (CD154)
- Deficiente cambio de isotipo a IgG e IgA
- Incremento en suero de IgM y inmunidad celular reducida (activación de macrófagos)
- Infecciones recurrentes
- Otros síndromes de hiper-IgM son autosómicos recesivos (mutaciones en CD40, AID, UNG, NEMO) El ligando de CD40 está expresado en la células T y se une a su receptor que está en la célula B. La interacción de las dos células mediante CD40 y el TCR- MHC provoca el correcto cambio de isotipo de IgM o IgD a IgG o IgA. Si hay mutaciones en el receptor de CD40 no puede haber una buena interacción entre las dos células y no hay un cambio de isotipo de Ig y un cúmulo de IgM que conlleva una inmunidad celular reducida por ejemplo una menor activación de macrófagos. Habrá una menor inflamación porque este CD40L interacciona también con el CD40 de los macrófagos de forma que la célula T puede activar al macrófago y que secrete una serie de mediadores proinflamatorios impidiendo una inflamación cuando hay una mutación.
Síndrome linfoproliferativo ligado al cromosoma X
- Se caracteriza por una incapacidad de eliminar EBV, infecciones de mononucleosis y aparición de tumores de células B asociados con hipogammaglobulinemia (no se produce gammaglobulinas).
- Mutaciones en SAP, SLAM associated protein (80% de casos) y XIAP, X-linked inhibitor of apoptosis (20% de casos) Se dan mutaciones en dos tipos proteínas, en ambos casos son proteínas que se encuentran en el cromosoma X pero es mucho mas fracuente en SAP (SLAM associated protein) con un 80% de los casos. En un porcentaje menor esta asociado a XIAP que es un inhibidor de la apoptosis. SAP: proteína que se une a SLAM que es un receptor que se expresa en la superficie de linfocitos y una vez unidos participan en la activación de quinasas implicadas en la señalización intracelular de linfocitos como FYN o LCK. Esto tiene como consecuencia: FYN-dependientes: que por ejemplo los linfocitos T secreten citocinas Th2, este implicado en la citoxicidad de estos linfocitos y en el desarrollo de NKT. FYN-independientes: participan en la interacción de linfocitos T y B y en la formación de centros germinales. XIAP: En un paciente normal, la célula infectada con el virus de EBV que suele ser linfocito B, interacciona un linfocito T via SLAM receptor con su ligando CD48 produciendose la casacada que da lugar a la lisis de la célula infectada. En un paciente enfermo la casacada de señalización no tiene lugar porque SAP esta mutado y por tanto no se produce la lisis de la célula infectada con el virus. Síndrome de linfohistocitosis hemofagocítica familiar
- Desgranulación defectiva en células NK y CTL
- Resolución defectuosa de infecciones víricas y activación de macrófagos incontrolada, que llegan a fagocitas eritrocitos
- Mutaciones en PERFORIN, RAB27A, MUNC13- 4 que son genes importantes en el proceso de citotoxicidad, en la formación de gránulos y su secreción. Problemas asociados a los efectos citolíticos de los linfocitos T o células NK. En estos pacientes sis células citolíticas tienen un déficit en la desgranulación de los gránulos que contienen las sustáncias para provocar la lisis de la célula diana y por tanto van a tener problemas en resolver infecciones víricas y van a tener una activación descontrolada de macrófagos que pueden llegar a fagocitar eritrocitos. Las proteínas Rab27a juegan un papel clave en la formación de gránulos citolíticos. La célula citolítica va a hacer contacto con la célula diana, se van a fusionar las membranas y van a inyectar vía perforina componentes como la granzima B que va a provocar la apoptosis de la célula diana.
Estos gránulos citolíticos se tienen que fusionar con la membrana de la célula efectora y secrete su contenido, por ejemplo en el proceso de anclaje participa MUNC13- 4. Después del anclaje se fusionan las membranas y esto provoca la secreción a través de la perforina de granazima B. Déficits en la formación de los gránulos citolíticos donde participa RAB27A, como en el anclaje de los gránulos a la membrana de la célula efectora donde participa MUNC13-4 y en la PERFORINA que facilita la inyección de las granazima B tiene como consecuencia final que no se elimine la célula infectada por el virus. Defectos en la activación de linfocitos T y B Las inmunodeficiencias congénitas pueden ser:
- Defectos en la señalización por TCR
- Defectos en la diferenciación a células plasmáticas ya sea por fallos en la maquinaria que provoca el cambio de isotipo o en la producción de AC y diferenciación a células plasmáticas.
- Defectos en la activación de las células T y en sus mecanismos efectores
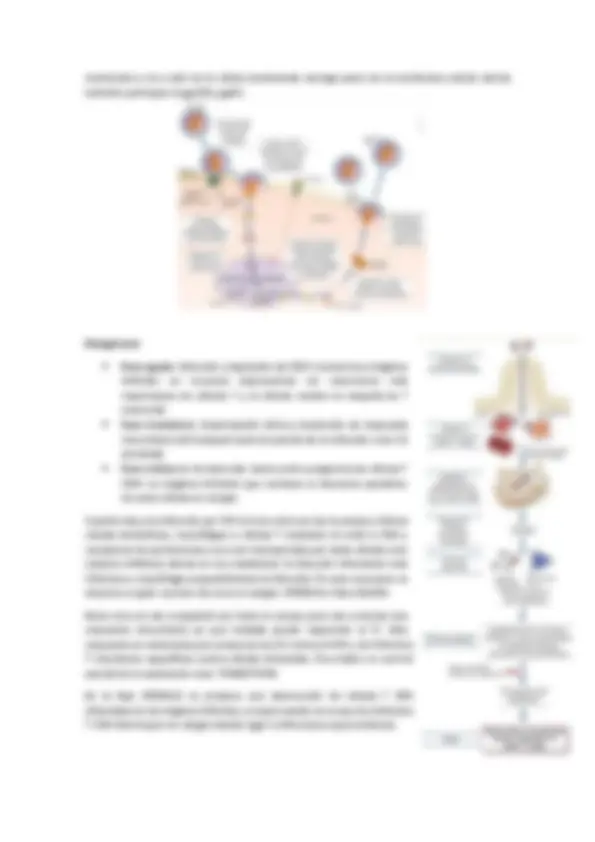
membrana y va a salir de la célula arrastrando consigo parte de la membrana celular donde también participan la gp120 y gp41. Patogénesis
- Fase aguda: infección y depleción de CD4+ memoria en órganos linfoides en mucosas (representan los reservorios más importantes de células T y es dónde residen la mayoría de T memoria)
- Fase transitoria: diseminación vírica y desarrollo de respuesta inmunitaria del huésped (control parcial de la infección a las 12 semanas)
- Fase crónica (o de latencia): destrucción progresiva de células T CD4+ en órganos linfoides que conlleva un descenso paulatino de estas células en sangre. Cuando hay una infección por VIH el virus entra en las mucosas y infecta células dendríticas, macrófagos o células T mediante la unión a CD4 y receptores de quimiocinas y va a ser transportado por estas células a los nódulos linfáticos donde se va a establecer la infección infectando más linfocitos y macrófagos expandiéndose la infección. En este momento se observa un gran numero de virus en sangre. VIREMIA o fase AGUDA Estos virus se van a expandir por todo el cuerpo pero van a activar una respuesta inmunitaria ya que todavía puede responder el SI. Esta respuesta se caracteriza por presencia de AC contra el VIH y de linfocitos T citocóxicos específicos contra células infectadas. Va a haber un control parcial de la replicación viral. TRANSITORIA En la fase CRÓNICA se produce una destrucción de células T CD infectadas en los órganos linfoides, es aquí cuando se ve que los linfocitos T CD4 disminuyen en sangre dando lugar a infecciones oportunísticas.
Durante la infección primaria los linfocitos T CD4 aun no se han eliminado. A medida que incrementa el num de partículas virales disminuyen los linfocitos T CD4. Esto es la fase aguda en la que se produce está diseminación de partículas virales por todo el organismo y la colonización de los órganos linfoides. En un punto el SI empieza a controlar la infección de modo que desciende en num de partículas virales en sangre debido a la eliminación por parte de nuestro SI de células infectadas, y vemos un repunte del num de linfos T CD4 pero poco a poco el virus va a ir infectando células T CD disminuyendo otra vez su número y subiendo el de virus. Fase de latencia. Se llega a un punto crítico en el que es favorable para infecciones oportunistas y aquí empieza la fase crónica reactivando la transcripción de partículas virales de VIH y producción de nuevos virus incrementando el número de virus en sangre. La producción de AC contra la cápside del virus y contra la p- 24 va augmentando a partir de la 8 ª semana. La pesencia de partícula virales en plasma sube muy rápido. Se produce la laténcia y el posterior repunte.