¡Descarga Transporte vesicular y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 2: TRANPORTE VESICULAR
Es la transferencia que ocurre a partir del retículo, se construyen a partir de un compartimento donador (RE), a partir de él se forma la vesícula, viaja por el citoplasma y funde con otro compartimento hasta el orgánulo receptor.
La maquinaria de transporte está muy conservada evolutivamente.
Hay diferentes pasos para el transporte vesicular:
Evaginación: es la formación de la vesícula. A partir de que una membrana se deforma forma la vesícula, esta deformación puede ser complicada o no, por ejemplo la membrana citoplasmática es rígida, por lo que tiene coste energético. Por el contrario para formar una vesícula del Golgi no lo sería tanto. Sea cual sea el sitio siempre hay unas proteínas de cubierta no se formaría la vesícula, son los que le dan la forma, la invaginan o evaginan. Además de esta función seleccionan el cargo que va dentro de la vesícula, esa selección es distinta para el cargo soluble que para el insoluble. El cargo que es de membrana posee dominios citosólicos que son reconocidos por las proteínas de cubierta. Para las proteínas solubles es distinto ya que hay una membrana que las separa, por lo que estas se seleccionan de manera indirecta a través de receptores de cargo, estas poseen un dominio luminar y citosólico. Por el lumen interaccionan con el cargo y por el dominio citosólico interaccionan con las proteínas de cubierta.
Tipos de proteínas de cubierta:
- Clatrina : Proteína hexamérica que forma un complejo con 3 brazos. En la célula se asocia a la membrana indirectamente mediante adaptadores, proteínas solubles que reconocen a las proteínas integrales de membrana, los seleccionan y agrupan. Por lo que hay un reparto del trabajo. Las proteínas adaptadoras son varias (ver tabla).
- COP : transporte de retículo a Golgi y viceversa. Es un heptámero que posee subunidades que seleccionan el cargo y otras deforman la membrana.
Fosfoinosítidos: es un fosfolípido con una cabeza polar con un grupo fosfato. Posee un anillo inosiltol (azúcar). Hay muchas quinasas que modifican este lípido fosforilandolo y desfosforilándolo, haciendo que el 10% de este lípido que se encuentra en total en la célula se encuentre de manera muy diversa. En la membrana hay mucho PI(3,4,5)P3. Hay sitios de reconocimento para estas moléculas muy específicos ya que los sitios de reconocimiento de anillos fosforilados son muy específicos. Eso determina las proteínas de cubierta y adaptadores que se adhieren a la membrana.
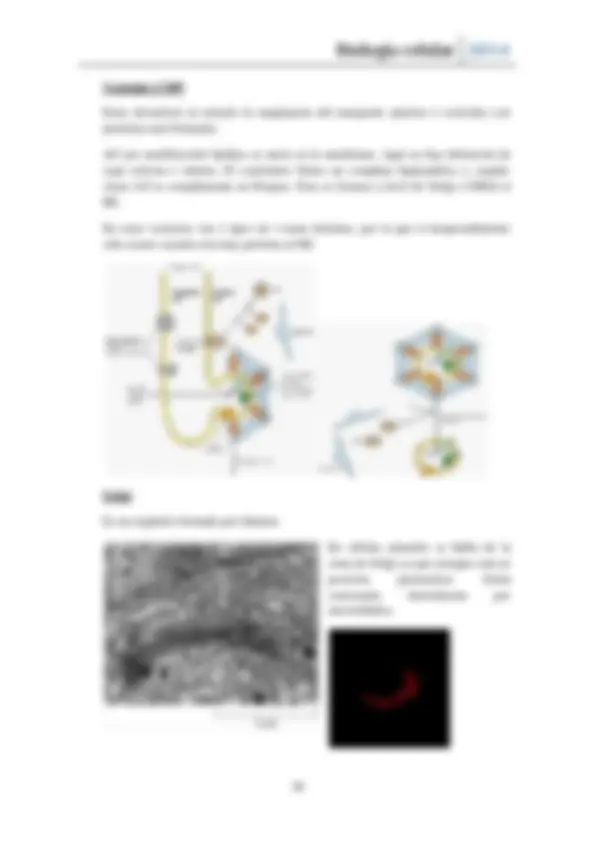
El proceso de ensamblaje de las cubiertas depende de GTPasas , proteínas que están de forma activas cuando tiene GTP e inactivas cuando tienen GDP. Estas controlan la asociación de proteínas a la membrana.
Sar1-GDP posee un extremo amino terminal que cuando está GTP unido está inactiva, en la membrana del retículo hay una proteína integral que se denomina Sar1-GEF , lo que hace es que activa a Sar1-GTP y une el dominio amino a la membrana, por lo que Sar1 queda unida a la membrana. Una vez unido se une el complejo COP2 que permite el paso del retículo al Golgi. En el caso de COP1 es al revés, viaja de Golgi a retículo y su proteína es ARF.
Rab aparte de reconocer, activa a proteínas efectoras. Un ejemplo son las proteínas motoras que permiten a las vesículas viajar por el citoesqueleto (Rab1). Otras reclutan las proteínas de enganche, solubles, que se asocian a la vesícula cuando hay una Rab activa, haciendo posible que la vesícula sea atraída a su orgánulo específico.
Formación de dominios específicos de membrana
Rab5-GDP junto a Rab-GEF se va reclutando en la membrana del endosoma temprano, además de reclutar a Pi3-kinasa , formándose PI(3)P en ella. Por lo que se forma un dominio de membrana con filamentos de enganche para captar vesículas. Cuando el endosoma temprano pasa a maduro éste se desprende de Rab5 y se carga de Rab 7.
Son los responsables de que cada orgánulo mantenga su identidad.
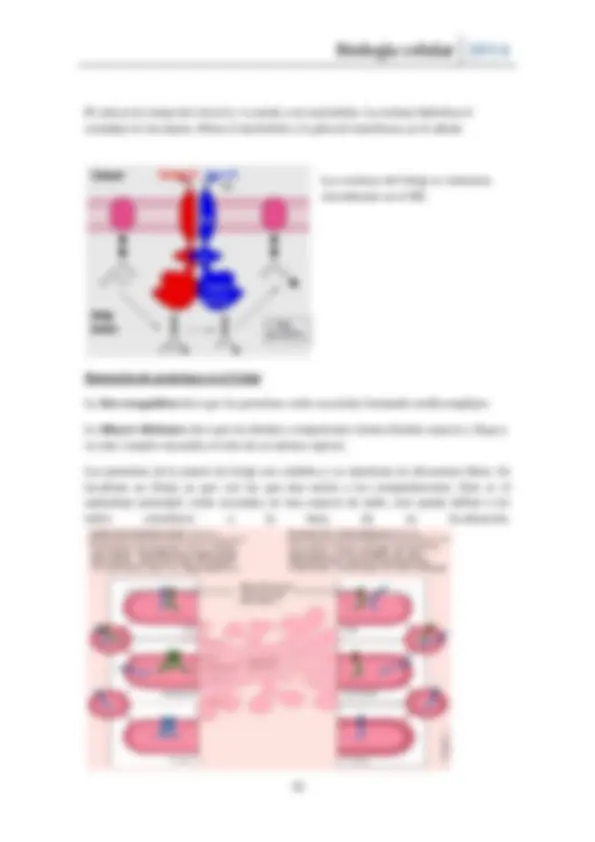
Fusión: Las SNARE son proteínas de tipo cargo específico que se forman en la vesícula ( SNARE-v ) y la membrana del orgánulo diana ( SNARE-t ). La fusión no está favorecida ya que hay agua entremedio de las membranas ya que los lípidos rehúyen del agua, acercarlas no ocurre de manera espontánea. En la formación del complejo SNARE ocurre la unión muy próxima por SNARE muy específico y afín que hace que la fusión sea espontánea.
Son proteínas de membrana, v-SNARE es una proteína única mientras que t-SNARE pueden ser dos o tres. Son proteínas coiled-coi que cuando se unen forman un haz de 4 súper hélices muy enrollado unidos por interacciones débiles. Para desengancharlas a la hora de fusionarlas con la membrana es difícil ya que ni un detergente no lo desnaturaliza debido a lo afín y específico que es. Una vez unidos se juntan tanto las membranas que acaban fusionando.
En neuronas las vesículas sinápticas funden con el terminar pre-sináptico cuando deben hacerlo, tienen su v- SNARE y t-SNARE, sin potencial de acción no se produce fusión pero se encuentran muy próximas. Cuando hay potencial de acción por la sinaptotagmina que posee sitios de unión a Ca permite la fusión de vesícula. La complexina por otro lado inhibe la fusión de membrana mientras no esté la sinaptotagmina activa ya que esta quita a la complexina, produciéndose la fusión.
Procesamiento de proteínas en el RE
Cuando se sintetizan las proteínas de membrana tipo GPI (proteínas de unión a PI). Esta se sintetiza en ribosomas asociados al retículo endoplasmático, en principio es una proteína tipo I, pero enseguida en su sitio de afectación se le une un fosfolípido de fosfatidil inositol junto con una serie de azúcares. Por lo que primero son insertadas y luego modificadas por una proteasa que lo transloca a un PI. Para eliminar esta proteína de la membrana es necesario una fosfolipasa.
Anclaje a GPI (glicosilfosfatidilinositol)
La proteína se une a un fosfolípido con GPI unido a un azúcar, estas proteínas en el retículo aparecen en la luz. Son proteínas importantes ya son señalizadoras. Otras tras ser sintetizadas se le añaden grupos hidrofóbicos, son proteínas aciladas, por lo que se anclan esporádicamente.
Otras se forman en ribosomas libres y se inserta posteriormente en la membrana.
Se produce a nivel de retículo y afecta tanto a proteínas solubles como las insolubles.
Glicosilación en el RE
Las proteínas aciladas se elaboran en ribosomas libres y se anclan a los ácidos grasos.
La glicosilación afecta al plegamiento de las proteínas, es necesario, otra función es la resistencia del ataque proteolítico. La mayor parte de las proteínas glicosiladas intervienen en el reconocimiento de la célula con el ambiente. La glicosilación es la adición de cadenas de carbohidratos unidos covalentemente, en función de esos carbohidratos está la función de esa molécula.
En el RE la glicosilación ocurre a través de un grupo amino del aa Asparagina. Cuando una proteína recibe los oligosacáridos su PM aumenta mucho. Esta ocurre co- traduccionalmente, es decir, cuando la proteína emerge de la proteína de translocación, es reconocido por proteínas de localización y ocurre la glicosilación.
El oligosacárdo se adhiere al dolicol fosfato presente en la membrana del retículo que posee 14 azúcares, sintetiza distintos oligosacáridos a la luz del citoplasma, se transloca y da al lumen del retículo. Este se incorpora de golpe a la proteína que entra en el retículo a través de una oligasacárido transferasa (OT).
En general, el hecho de que sea glicosilada demuestra que la proteína sea madura o no y pueda abandonar o no el retículo.
FORMACIÓN Y REORDENACIÓN DE LOS PUENTES DISULFURO:
Se da en el retículo exclusivamente, la PDI (protein-disulfuro-isomerasa) interacciona con las proteínas que se están formando , las que poseen sulfuros y lo que hace es formar puentes disulfuros.
Los puentes disulfuro se crean por proximidad, esta proteína lo que hace es que interactúen en residuos de azufres lejanos. Esta enzima es un componente de la luz del retículo e interaccionan con las proteínas en formación y ayuda a su conformación.
Su función es la formación de puentes disulfuros y los reordena.
Cuando la PDI está en estado oxidado forma puentes, pero necesita reoxidarse cuando forma puentes disulfuro, a través de Ero1 se reoxida.
dependiente de Calcio para que se unan estas proteínas necesitan que los oligosacáridos terminen en una N-Glucosa, en una sola. Para las que poseen más de una hay glucosilasas que las rompen y permiten que se unan. Sólo se unen a proteínas inmaduras, no plegadas. Cuando una proteína se sintetiza en el RE, las chaperonas las van ayudando a que adquiera su conformación nativa, es un mecanismo de retención donde proteínas inmaduras no abandonan el retículo. Para eso no solo necesitan chaperonas sino una enzima como la glucosiltransferasa que incorpora azúcares a las proteínas mal configuradas, de forma que esa proteína es reconocida por calnexina o calreticulina que está ahí, cuando ya se forma la glicosilasa le quita las glucosas y sale del RE.
Control de calidad en el RE
EN el retículo se monitoriza el estado de las proteínas y su oligomerización. Molecularmente son los componentes endógenos que intervienen en el plegamiento y supervisión de las proteínas.
- Chaperonas: BiP (Hsp70), Calnexina, Calreticulina, etc
- Proteínas Plegadoras: PDI, Petidil prolil isomerasa
Cuando por mucho que se intenten plegar bien no se puede conseguir una proteína madura, por lo que la proteína se degrada, pero primero se retrotransloca por ERAD , por unas chaperonas que la llevan, una retrotranslocasa la retrotransloca y por la vía ubiquitina-proteaoma se degrada.
Cuando la manosidasa actúa significa que esa proteína ha sido tratada durante mucho tiempo y no ha logrado conformarse nativamente.
Respuesta a proteínas mal plegadas (UPR Unfolded protein response)
Puede darse en situaciones de estrés por ejemplo, choque térmico, si una célula se incuba a una Tª mayor a su temperatura normal (36ºC), muchas proteínas no llegan a plegarse de forma normal. No solo por choque térmico se puede dar, dando un exceso de proteínas mal plegadas.
Para dar salida a eso lo que hace es producir más chaperonas, más proteínas de retrotrasnlocación, etc. En definitiva se activan los genes del control de calidad que tienen que ver con el plegamiento de las proteínas y su maduración.
Para eso tiene que haber una señal que vaya del retículo al núcleo y ocurre en todos los eucariotas. En una célula donde hay estrés en la membrana del RE hacia el lado del retículo donde hay proteínas mal plegadas poseen muchas proteínas unidas a chaperonas que se unen a receptores, por lo que si aumentan el número de proteínas mal plegadas por ejemplo la chaperona Bip , esta no interactúa con el receptor por lo que la célula lo interpreta como una falta y expresa los genes de síntesis de Bip.
Lípidos de membrana
Todos los lípidos vienen del RE
- Fosfolípidos: Fosfatidil-serina, fosfatidil-colina, esfingomielina, fosfatidil- etanolamina. La estructura es una molécula de glicerol, 2 cadenas de ac. Grasos y un grupo fosfato. Los 4 fosfolípidos se sintetizan de manera igual. (diapo 42). Los fosfolípidos se insertan en el lado citosólico de la membrana por lo que tiene que haber un equilibrio entre en lado citosólico y el lumen del retículo, hay una enzima que transloca esos fosfolípidos y es específica, produciéndose una simetría en cada cara de la membrana.
- Colesterol: La maquinaria está en el retículo. La ceramida es un compuesto que se forma de la condensación de un ácido graso con una serina. Con esta cuando viaja al golgi se formará la esfingomielina y los glucolípidos. Las gónadas segregan hormonas lipídicas por lo que poseen un REL muy desarrollado.
- Glicolípidos
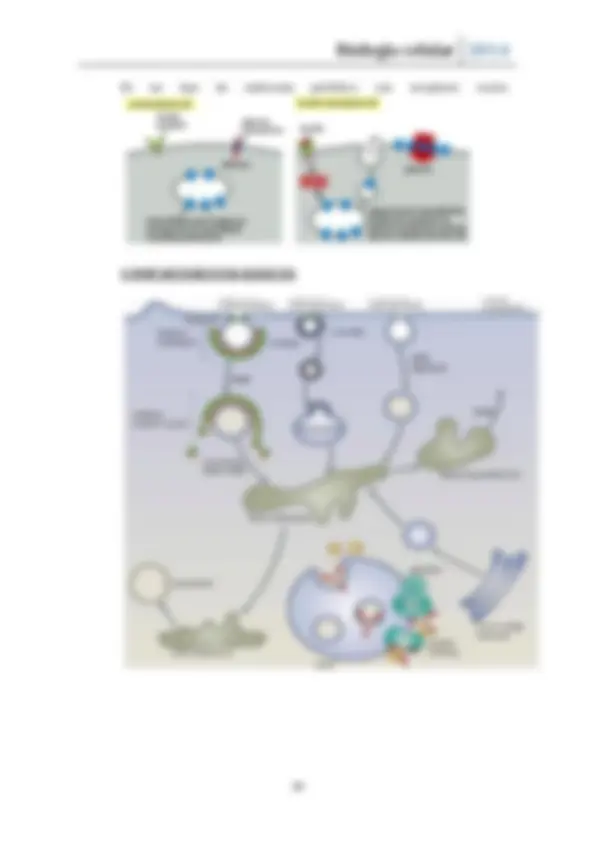
Exportación molecular
El sitio de salida del retículo (exit site – Eres), George Palade lo descubrió y lo denominó retículo de transición. Pero actualmente se denomina retículo de salida. Son los lugares donde se forman las vesículas con destino al Golgi.
En la interfase entre el retículo y Golgi se forma una zona especial que no hay ribosomas y se forman vesículas con cubiertas con COPII permitiendo el trasiego de moléculas entre el RE y Golgi. Este transporte también puede ser al contrario, aquellos componentes que son necesarios en el retículo vuelven el COPI.
COPII es una proteína que forma un complejo que envuelve a las vesículas, se forma cuando Sar1 se une a la membrana, luego se forma un dímero de Sec23 (activa) y Sec24 que seleccionan el cargo y luego se une Sec13 y Sec31 que deforma la membrana y la cubierta. Luego cuando Sar hidroliza GTP la cubierta se desprende, pero es necesario que forme la vesícula y lleve el cargo.
Hay proteínas solubles que dadas la gran cantidad que hay no hay receptores de señal para su transporte al golgi. (ejemplo de la albúmina)
Retención de proteínas incompletas
Fusión homotipica, cuando se forma una estructura irregular que va por microtúbulos y se acerca al golgi.
Las moléculas bien ensambladas tienen la capacidad de salir del RE, si están mal ensamblada se retrotransportan, se ubiquitinizan y se destruyen.
Si se ensambla de manera correcta, se forma el complejo Sec13 y Sec31, es el que hace la función mecánica de deformación de la membrana. No se sabe bien que proteína hace que se estrangule la vesícula y se libere.
Esa cubierta sale ya que Sar hidroliza GTP.
La fusión homotípica es la fusión de vesículas iguales, el resultado de ello es la formación de una estructura pleomórfica (compartimento intermedio entre el retículo y
el Golgi – ERGIC ). Resulta de la fusión de vesículas COPII y su sitio final será el golgi guiadas por proteínas motoras. Se le considera un orgánulo independiente con composición distinta al golgi (no se sabe muy bien).
De ella surgen vesículas COPI o de reciclaje.
Recuperación de las proteínas resistentes
Las proteínas de membranas llevan en su extremo citosólico o carboxi terminal un motivo dibásico (KKXX-COOH o RRXX-COOH) por el que son reconocidos y envueltos en vesículas.
Las proteínas solubles también poseen una secuencia KDEL
El receptor de KDEL reconoce las proteínas endógenas del RE. Ese receptor está concentrado en ERGI o cis-Golgi, este une los ligando que salen del retículo accidentalmente, evitando así que salgan vesículas que no deben. De esta forma las vesículas vuelven al RE en vesículas de reciclaje. Este receptor debe reconocer con mucha afinidad al ligando y cuando llega al retículo su afinidad tiene que bajar ya que lo libera en un sitio que hay mucha concentración de ese ligando, por lo que sufre un cambio conformacional dependiendo del pH y la concentración iónica.
En células vegetales no es así, aquí cada uno de los dictiosomas o stack está una sobre otra dispuesta de manera repartida por la célula.
Tienen en común que la cara de entrada es la cis y la de salida es la trans, por lo que un orgánulo polarizado.
En principio las cisternas del lado cis y trans parecen iguales, pero se ha demostrado que eso no es así ya que por tinciones citoquímicas se obtenían tinciones en distintas partes del golgi que no aparecían en otras partes del mismo. Por lo que una enzima no está repartida por todo el golgi, es un orgánulo compartimentalizado.
Se habla de orgánulo compartimentalizado pero no cada cisterna es un compartimento.
El primer compartimento o red cis del Golgi es un entramado membranoso. (ver disposición de las enzimas en la imagen de la izquierda)
El polo distal o trans es un entramado de microtúbulos que se podría considerar un orgánulo independiente.
Glicosilación en el complejo de Golgi
Es la más común.
No hay molde para la síntesis de carbohidratos, sin embargo cuando se produce una glicoproteína en un orgánulo se produce siempre igual y de la misma manera. Pero si cambiamos de tipo celular ya cambia.
Hay dos tipos de uniones de azúcares a proteínas, la N-glicosilación (aaasparagina). Y otra es la unión de aa con grupos hidroxilos (serina o treonina), formándose un enlace éster por lo que es una O-glicosilación.
Los N-oligosacáridos llegan al golgi en parte ya procesados, aquí hay manosidasas y se le añaden otros azúcares y al final lo que sale no tiene nada que ver con los oligosacáridos que venían del RE.
El oligosacárido que se le añade en el retículo siempre es el mismo por eso siempre se le denomina oligosacárido patrón, y éste nunca es igual al que acaba saliendo del Golgi.
Los O-unidos están unidos a aa tipo serina o treonina, asociado a células mucosas como son las células oxínticas. Estos se unen en golgi de manera secuencial y su secuencia