Scarica Microbiologia VIRUS e più Appunti in PDF di Microbiologia solo su Docsity!
MICROBIOLOGIA- MODULO 3
VIRUS
Già dalla fine dell’800’ si riconobbe l’esistenza di queste piccole particelle, più piccole dei batteri, agenti di infezione, definite come ORGANISMI FILTRABILI (date le dimensioni). Dal momento che però a quei tempi non vi erano effettivamente i mezzi per andare a osservare direttamente questi, rimase comunque un’ipotesi. La prima dimostrazione di ciò arrivò nel 1898, da parte di LOEFFER e FROSCH, dove è stata trasferita l’afta epizootica (dermatite animale) attraverso l’uso di un filtrato acellulare. Nel 1953 vi fu, da parte di STANLEY, l’isolamento in forma cristallizzata del virus mosaico del tabacco (quindi le prime immagini). Oggi abbiamo la conferma dell’esistenza dei virus e siamo in grado non solo di visualizzarli ma anche di coltivarli. Il punto interrogativo principale tutt’ora riguarda la loro natura, si tratta di esseri viventi o solo di molecole nucleoproteiche? La caratteristica di filtrabilità si è dimostrata infatti comunque inadeguata al fine di distinguere i virus da batteri più piccoli (come i Rickettsia); per cui la dimensione non è un parametro adeguato per distinguere delle particelle virali. Inoltre il grande dibattito sul fatto che si tratti di essere viventi o meno sta sul fatto che da un lato microorganismo vivente è un organismo in grado di fare sintesi proteica (quindi di farsi le sue stesse proteine e strutture), ma sappiamo benissimo che i virus sono dei parassiti, per cui hanno bisogno di infettare una cellula per potersi replicare (sfruttando appunto i sistemi replicativi della cellula ospite infetta). Di contro però, un’altra corrente di ricercatori ritiene che si tratti di esseri viventi in quanto posseggono un proprio genoma. Le differenze fondamentali comunque, che contraddistinguono i virus sono:
semplicità della loro organizzazione strutturale;
modalità replicative (diverse, poiché comunque nei virus noi troviamo un genoma ad RNA, che invece di solito negli altri organismi ha una funzione diversa, e non costituisce genoma). I VIRUS sono definiti come delle ENTITA’ METABOLICAMENTE INERTI, in quanto INCAPACI di esplicare funzioni metaboliche e respiratorie (autonomamente almeno): infatti si collocano ipoteticamente al limite tra il vivente e il non vivente! I virus, per moltiplicarsi/ replicarsi, devono “reclutare” (entrare/ infettare) una cellula ospite (da sfruttare ai fini della propria replicazione, cioè sfruttano i sistemi replicativi della cellula per la sintesi delle proprie proteine). I Virus sono ELEMENTI GENETICI che replicano indipendentemente dal genoma dell’ospite, ma NON dal suo apparato metabolico e biosintetico (in quanto sfruttano questo ai fini della replicazione del proprio genoma).
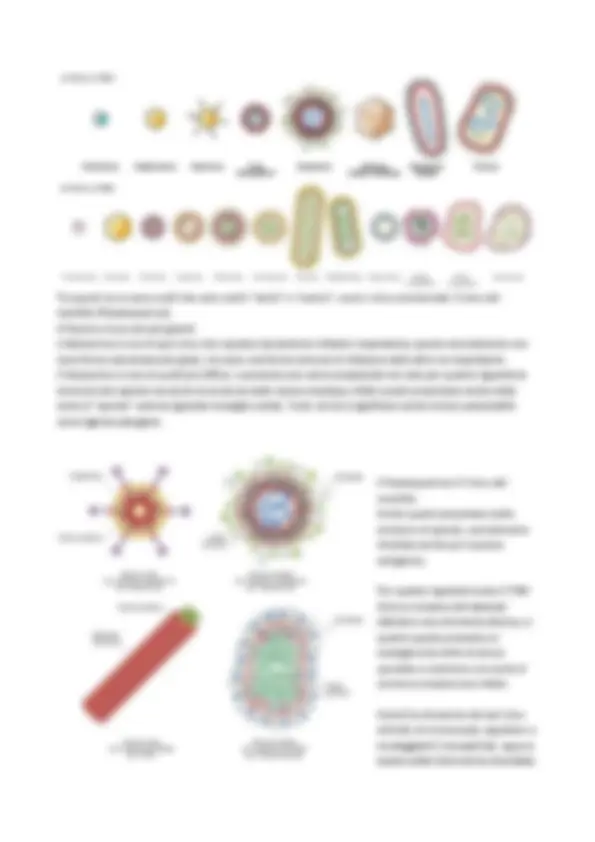
Sempre a tal fine di replicarsi sono capaci infatti di RIPROGRAMMARE tutto il metabolismo della cellula ospite. Pertanto un VIRIONE è una particella relativamente SEMPLICE, poco più di un blocco di materiale genetico racchiuso da un involucro di natura proteica, il CAPSIDE. A differenza degli altri microorganismi, i virus non possiedono un proprio apparato per l’utilizzazione e la trasformazione dell’energia e per la sintesi delle proteine virali (infatti necessitano e sfruttano quello della cellula ospite che vanno ad infettare). Sostanzialmente infatti tutti i virus hanno in comune questo meccanismo replicativo di base, ma differiscono poi notevolmente per altre caratteristiche quali dimensioni e forma, composizione chimica del genoma e tipo di cellula ospite. Infatti quello che andremo a vedere principalmente sono le diverse classi replicative dei virus (cioè le diverse modalità di replicazione dei virus, in quanto possono presentare differenti genomi). Generalmente la DIMENSIONE dei VIRUS può andare dai 20 ai 400 nm. Quindi quando andiamo a vedere le caratteristiche virali non si tratta sostanzialmente del fatto che siano particelle molto piccole, ma si tratta piuttosto di estrema semplicità nella sua organizzazione, data dalla sua sola struttura proteica e nel relativo contenuto genomico, e inoltre anche nella sua capacità replicativa. I VIRUS vengono suddivisi principalmente in VIRUS A DNA e VIRUS A RNA (immagine sotto). Quindi per la prima volta l’RNA ha una funzione di materiale genico. Andando dalle strutture più piccole e semplici a quelle più complesse e articolate, vengono a loro volta suddivisi in virus NUDI (che non hanno un altro rivestimento, che prende il nome di ENVELOPE) e sono delle strutture molto più complesse, mentre quando abbiamo strutture avvolgenti la struttura del capside si parla di VIRUS RIVESTITI (perché presentano quest’ulteriore struttura di rivestimento del capside) e che generalmente troviamo nei virus animali (i fagi infatti ad esempio non la presentano). Questa ulteriore struttura di rivestimento deriva dal modo in cui questi virus (che hanno infettato una cellula eucariotica) fuoriescono dall’infezione, perché alcuni di questi determinano il processo di GEMMAZIONE (quindi fuoriescono dalla cellula che infettano e con questa si avvolgono in questa struttura).
Vengono definiti VIRIONI quando i virus si trovano in FORMA EXTRACELLULARE, altamente infettiva, nella quale si possono trasmettere da un ospite ad un altro (non possono stare molto tempo senza “parassitare”). I componenti principali delle particelle virali libere sono: il genoma (a DNA o RNA), associato con proteine o poliamine (il genoma, all’interno della struttura del capside è comunque protetto e avvolto da proteine virus-specifiche); poi presentano un rivestimento proteico, detto CAPSIDE, costituito dall’aggregazione di proteine tutte uguali, dette CAPSOMERI, che conferisce la simmetria alla particella (e contenente il genoma), che talvolta può essere a sua volta circondato da un rivestrimento membranoso di natura lipidica, detto ENVELOPE (virus rivestiti). Alcuni virioni posseggono inoltre degli enzimi, necessari nelle fasi iniziali della replicazione del genoma (enzimi che non ritrovano magari nella cellula che vanno ad infettare). Quindi in un VIRIONE distinguiamo la porzione del RIVESTIMENTO, con un CAPSIDE di natura proteica (costituito dall’aggregazione delle proteine CAPSOMERI) e talvolta di un ulteriore rivestimento di natura lipidica, l’ENVELOPE; e la porzione del CORE INTERNO, in cui ritroviamo l’ACIDO NUCLEICO e tutte le proteine virus-specifiche ad esso associate (enzimi virus-specifici), che a loro volta vanno a “riempire” tutto quello spazio tra la struttura del capside e il core nucleico, e in un certo senso proteggendo quest’ultimo. Nel momento in cui abbiamo ACIDO NUCLEICO+ CAPSIDE stiamo parlando del NUCLEO-CAPSIDE. Questi NUCLEO-CAPSIDI sono sempre costruiti in maniera simmetrica (sono organizzati con una simmetria altamente programmata). Infatti l’informazione per una corretta e precisa aggregazione delle subunità strutturali in capsomeri è già contenuta nella natura delle proteine stesse che la compongono (il virus non improvvisa una sua simmetria sul momento, ma l’informazione per la sua formazione è già implicita e presente nelle proteine che la dovrebbero costituire), con un informazione, un messaggio, di AUTO- ASSEMBLAGGIO, dettato già dal genoma. Le forze che legano insieme tutti i capsomeri non sono forze covalenti ma sono interazioni idrofobiche ed elettrostatiche. I virioni quindi sono dati dalle interazioni proteina-proteina, proteina-acido nucleico e proteina-lipidi. La SIMMETRIA quindi (modo in cui le subunità strutturali dei capsomeri si aggregano, si organizzano, nella formazione del capside) è già ben definita (genomicamente). CAPSIDE Le funzioni principali del capside sono: protezione (del materiale genico, nel momento in cui è nella forma di virione), riconoscimento recettori (a livello di questa struttura infatti vi sono tutti i recettori di riconoscimento e interazione con l’ospite), e trasporto del genoma al nucleo. Inoltre il capside svolge un ruolo STRUTTURALE, dal momento che a partire dalle subunità irregolari dei CAPSOMERI si andranno a costituire le strutture regolari, le 2 simmetrie tipiche per l’architettura del virione: la SIMMETRIA ICOSAEDRICA (tipica del gruppo di adenovirus, herpesvirus, picornavirus), costituita
da 20 facce e 12 vertici; oppure la SIMMETRIA ELICOIDALE (tipica di TMV o Rhabdovirus), in cui il genoma si avvolge con queste subunità dei capsomeri a costituire quello che a noi sembra visivamente al microscopio un “bastoncello”. Inoltre vi piò essere una STRUTTURA COMPLESSA, ovvero quella che ritroviamo in tutti i FAGI della serie “T” (come il T4), in cui distinguiamo una testa glicosaedrica, tutta una parte più allungata che costituite la guaina contrattile (struttura tubulare) e poi la piastra basale, all’esterno della quale ci sono tutte le fibre e le spicole. Questa struttura particolare consente di norma un facile riconoscimento dei fagi: normalmente quando il fago fa infezione, attraverso la piastra basale abbiamo il riconoscimento della cellula ospite, la guaina si contrae e questo fa sì che il fago inietta il genoma virale all’interno della cellula ospite. Normalmente nelle infezioni fagiche non entra mai la struttura capsidica ma entra solo il genoma fagico all’interno della cellula ospite, che viene iniettato. Altra simmetria del CAPSIDE ICOSAEDRICO (tipica dell’Adenovirus). Ci sono tutte le proteine del capside organizzate in esoni, eptoni… a seconda di come vengono ad assemblarsi. Inoltre presentano tutte queste FIBRE (o ANTENNE) che contribuiscono al riconoscimento, all’attecchimento e all’infezione. In generale visivamente al microscopio appaiono come “palline”.
- alcuni virus presentano inoltre delle glicoproteine di superficie con attività neuroaminidasica. Le NEURAMINIDASI hanno un ruolo funzionale per la penetrazione del virus (nella cellula) in quanto sono in grado di distruggere l’acido sialico delle membrane dell’ospite, consentendo così l’attacco dei virioni, la penetrazione del genoma, l’emoagglutinazione, la fusione e l’emolisi.
- nella porzione più centrale è possibile ritrovare delle PROTEINE DI MATRICE, le quali rafforzano l’envelope (non sono glicosilate). Tutte queste PROTEINE VIRALI si possono distinguere ulteriormente in:
PROTEINE FUNZIONALI Permettono ad esempio all’acido nucleico virale di replicarsi (DNA/RNA polimerasi).
PROTEINE STRUTTURALI Fanno parte della struttura stessa virus. Infatti sono incorporate nelle nuove particelle virali come costituenti del capside o come strutture particolari (es: emoagglutinine) presenti nell’envelope.
PROTEINE CHE ALTERANO ALCUNE FUNZIONI E STRUTTURE DELLA CELLULA OSPITE
PROTEINE STRETTAMENTE ASSOCIATE AGLI ACIDI NUCLEICI
PROTEINE DI MATRICE (M) Per quanto riguarda la composizione chimica dei virus, vi sono diversi ENZIMI importanti per l’infezione virale (anche se mancano gli enzimi deputati ad esempio alla produzione di energia o coinvolti nelle vie biosintetiche dei precursori):
LISOZIMA (presente in alcuni batteriofagi, fondamentale per degradare la parete della cellula ospite e consentire così l’accesso del proprio genoma virale).
NEURAMINIDASI (rientrano negli Ortho e Paramyxovirus, virus influenzali e del morbillo).
PROTEASI VIRUS-SPECIFICHE
RNA POLIMERASI RNA-DIPENDENTE (per i virus ad RNA, fondamentali per i processi di replicazione).
TRASCRITTASI INVERSA (tipica dei RETROVIRUS ed è una DNApol RNA-dipendente, ovvero un enzima in grado di copiare dal DNA, all’RNA e da questo nuovamente al DNA->cDNA).
RNA POLIMERASI DNA-DIPENDENTE (nei Poxvirus). (immagine sopra) esempio di uno dei virus più complessi e studiati, ovvero il virus dell’IMMUNODEFICIENZA ACQUISITA (HIV). Appartiene al gruppo dei RETROVIRUS. Presenta un genoma ad RNA, a cui sono associate delle proteine strutturali che lo aiutano e lo proteggono.
Presenta il rivestimento dell’envelope, e tra questo e la struttura capsidica vi è tutta la parte di proteine che costituiscono la matrice, riempendo così tutto lo spazio interno e conferendo allo stesso tempo maggiore robustezza. Nel genoma di questo virus sono presenti i geni gag, fondamentali geni strutturali, i geni pol che codificano per le polimerasi e i geni env che codificano per l’envelope. Tutta la parte dell’identificazione del virus dell’HIV viene fatta proprio attraverso il sequenziamento e la caratterizzazione dei geni pol, e quindi delle polimerasi, che riescono a darci anche un’indicazione identificativa relativamente al virus. (immagine a sx) altro virus animale piuttosto famoso è il virus dell’influenza, ORTHOMYXOVIRUS. Anche questo rappresenta una struttura un po’ più complessa, in quanto presenta un genoma ad RNA frammentato in 8 segmenti. Presenta la struttura esterna dell’envelope, e in questo envelope vi è tutto il sistema delle emagglutinine e delle neuraminidasi, motivo per cui questo virus ha un potere abbastanza elevato a livello di penetrazione, proprio grazie al sistema delle neuraminidasi. COLTIVAZIONE DEI VIRUS Non è possibile coltivare i virus così come si fa con i batteri o con piccoli organismi eucariotici (quindi con i terreni di coltura etc..), perché i virus sono incapaci di riprodursi indipendentemente dalle cellule (che vanno ad infettare), in quanto parassiti obbligati. Per molti anni (ed ancora oggi in qualche caso) i ricercatori hanno iniettato i virus animali in animali suscettibili oppure in uova embrionate di pollo. Più recentemente la coltivazione dei virus è effettuata in colture cellulari su monostrati di cellule animali (in delle fiasche), infettando queste cellule con i virus praticamente (->DETERMINAZIONE QUANTITATIVA). Questo tipo di coltivazione delle linee cellulari, definito come SAGGIO DI PLACCA, è una tecnica resa possibile dallo sviluppo di terreni di coltura per cellule e soprattutto dall’avvento (e quindi dall’uso) di antibiotici ed antimicotici, capaci in tal modo di prevenire la contaminazione batterica e i funghi. Questo perché per le linee cellulari, quando vengono fatte crescere, si utilizzano dei terreni fortemente nutritivi: si mette il serio bovino, albumina, cioè tutto ciò che fa star bene le linee cellulari, e ciò vuol dire che è un terreno fertilissimo, per i batteri ma anche per i funghi. Per cui ogni volta che si vuole far crescere una linea cellulare si mettono anche degli antibiotici e antimicotici, in modo da garantirne la crescita in un ambiente poco o niente contaminato quindi e oltretutto più pulito possibile (basta una piccolissima contaminazione per distruggere tutte le linee cellulari precedentemente coltivate accuratamente). Quando invece abbiamo a che fare con BATTERIOFAGI, le nostre linee cellulari sono più semplici in quanto rappresentate dagli stessi batteri.
In che cosa consiste il test dell’ELISA (saggio immunoenzimatico)? Esistono 2 tipologie di test: il test DIRETTO con ELISA, ed il test INDIRETTO. Il test DIRETTO è quello che va ad identificare la presenza del virus. (immagine a sinistra) base, come le piastrine di un microtiter (piastrine tipiche con cui si fanno le MIC), pozzetti di plastica piccoli, in cui già si trovano degli anticorpi per uno specifico virus. A questo punto si aggiunge il siero del paziente X, oppure anche il virus stesso (però se si vuole fare un test ad un paziente ad esempio per sapere se ha avuto il morbillo, si lavora sul prelievo di sangue). Se il siero del paziente risulta positivo relativamente alla presenza di questo dato virus, allora ci sarà una reazione specifica anticorpo-antigene (l’antigene sarebbe il virus, oppure comunque strutture della membrana se ha l’envelope, oppure strutture capsidiche, che vengono normalmente identificate come strutture antigeniche). Ora si pone un problema: abbiamo l’anticorpo adeso alla superficie del microtiter, il virus o il suo antigene- virus specifico che si è legato, devono avere un sistema per poterlo evidenziare/vedere. In questo caso allora si aggiungono altri anticorpi. (immagine sopra) anticorpo fisso alla superficie della piastrina, la stellina che sarebbe l’antigene virale, aggiungo un altro anticorpo il quale si andrà a legare al virus però questa volta questo anticorpo sarà marcato (solitamente questo anticorpo è coniugato con la fosfatasi alcalina). Si fanno poi dei lavaggi per eliminare tutto ciò che non è perfettamente legato. Si aggiunge il substrato (come p-nitrofenilfosfato) il quale darà un effetto di fluorescenza, o meglio colorimetrico, cioè dà un colore che va dal blu più intenso al blu meno intenso (più è intenso il blu e più significa che ho una carica virale positiva). In questo modo normalmente è possibile non solo vedere se è positivo o negativo ma diventa possibile anche quantificare. Questo test dell’ELISA è il test più comunemente usato in ospedale (e più facile) per sapere per esempio se si ha avuto un’infezione da morbillo, e può essere DIRETTO (cioè nella piastrina troverò adesi gli anticorpi specifici per quel virus, quindi si sfrutta in questo caso un legame specifico fondamentale che è quello antigene°-anticorpo) o INDIRETTO. °l’antigene è tutto quello che noi non identifichiamo come nostro “self”, tutto ciò che è estraneo.
Quindi per i virus la forte specificità del legame antigene-anticorpo viene sfruttata a fini identificativi, ma anche quantitativi, della carica infettiva in questo caso. E’ possibile infatti avere una titolazione perché il siero in studio è in diluizione: si allestiscono queste piccolo microtiter dov’è possibile fare queste diluizioni scalari in base 10. Nel modo INDIRETTO invece al posto di mettere nelle piastrine del microtiter l’anticorpo si può mettere invece l’antigene virale, virus-specifico (ad esempio adeso si può mettere, anziché l’anticorpo, una struttura del virus del morbillo con il suo potere antigenico). A questo punto andrò a ricercare gli anticorpi che si legheranno al dato virus, se lo trovano. Si usa però un anti-anticorpo, ovvero un altro anticorpo che va a legare questo complesso ed è sempre coniugato al fine di dare sempre un effetto colorimetrico. Quindi in questo sistema, a seconda dell’obiettivo, sostanzialmente possiamo mettere fissi in questa piasstrina o l’antigene o l’anticorpo, perché comunque il punto principale è la forte specificità del legame antigene-anticorpo. Dopo che è avvenuto questo legame bisogna aggiungere un altro sistema che consente l’evidenziazione di questo legame (effetto colorimetrico): se è una anticorpo avremo l’anti-anticorpo, se il virus avremo il suo rispettivo anticorpo. Si forma una sorta di struttura a sandwich. SAGGIO DELLE PLACCHE Sistema più banale e comune per poter vedere un’infezione. (immagine a sx) abbiamo una sospensione virale ed una sospensione cellulare (generica, in quanto dipende dal tipo di virus che si vuole andare a saggiare-> ammesso che si tratti di un batteriofago si metterà un sospensione cellulare batterica). Il virus penetra e infetta attraverso un recettore specifico. Ad esempio, volendo fare un’infezione con lambda, sappiamo che questo infetta E.coli, quindi avremo una sospensione cellulare di E.coli (forte specificità). Si usa poi un agar molle dove unire queste due sospensioni, e poi si piastra il tutto. Si lascia incubare e poi si può avere l’effetto citopatico, che determinerà la lisi (se lambda fa il ciclo litico, determinerà la lisi e quindi la formazione su piastra di placche) e quindi delle placche di lisi, cioè dei punti in cui si replica e replicandosi determinerà la lisi della cellula.
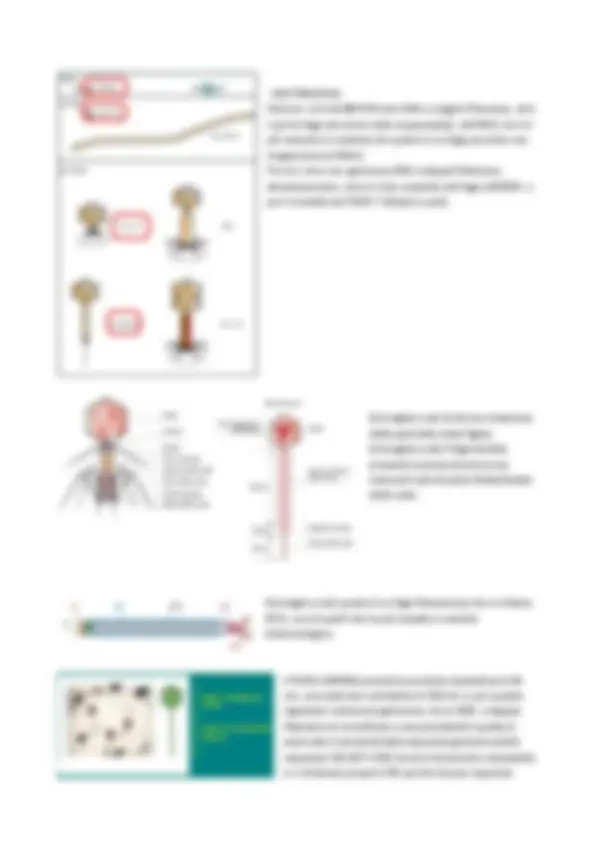
Esattamente come per le colonie batteriche, quando non si può fare una conta ed è necessario effettuare diluizioni, la stessa cosa si deve fare anche nel caso delle infezioni virali. Se si dovesse fare una prima sospensione e non si fa alcuna diluizione, si potrebbe avere una carica così elevata da non riuscire a contare tutte queste placche. Per cui è necessario fare diluizioni seriali (esattamente come per le colture batteriche): 10^-1, 10^-2, 10^-3…. (immagine sopra) piastrine a pozzetti in serie delle MIC, in cui si fanno diluizioni seriali, al fine di andare a vedere la titolazione degli anticorpi (nel caso in cui si tratta di un test specifico antigene-anticorpo) o la titolazione virale (come quello di prima) utilizzando un sistema colorimetrico, così da contare effettivamente la quantità scalare. In questo caso solitamente, questo tipo di analisi con queste piastrine così non sono con i batteriofagi (perché per questi si usano delle piastre normali) ma adesi vi sono delle linee cellulari animali per esempio. FASI CICLO REPLICATIVO VIRALE 1 - Attacco del virione alla cellula Questo avviene per mezzo di un legame fortemente specifico, in quanto ogni virus, ogni fago ha una sua tipologia di ospite specifica). Nel caso specifico dei fagi avremo la fase dell’ Absorbiment o, cioè il momento in cui si stabilisce il contatto e il legame tra virus e cellula da infettare (=riconoscimento specifico tra recettore virale e recettore cellula ospite-> recettori più comuni della cellula batterica cui questi virus si legano possono essere l’acido lipoteicoico nel caso dei Gram+, oppure tutte le strutture proteiche esterne nei Gram-, la struttura stessa del Pilus è uno dei target principali per questo riconoscimento ). 2 - Penetrazione (-> Uncoating / Perdita dei rivestimenti )
I fagi non entrano mai con tutta la struttura capsidica, ma questa resta fuori, in quanto in seguito al riconoscimento e all’adesione, l’unico a penetrare è il genoma virale (per cui si perde il rivestimento capsidico che non penetra). 3 - Sintesi dei componenti virali (proteine virali e replicazione dell’acido nucleico virale) Nel momento in cui questo genoma virale è penetrato all’interno della cellula, si attuano quelli che sono i 2 obiettivi principali, ovvero la sintesi di tutti i componenti virali fondamentali quali proteine virali e soprattutto la replicazione del genoma virale, e contemporaneamente andare a bloccare la sintesi della cellula ospite. Quindi il virus inizia a replicare se stesso e allo stesso tempo agisce in modo negativo nei confronti della cellula ospite, bloccandone i propri processi di sintesi (in modo da consentire sintesi e replicazione solo di materiale virale). 4 - Assemblaggio delle proteine virali con una copia di acido nucleico neosintetizzato Dopo la sintesi di tutte queste proteine virali, queste devono essere assemblate al fine di costituire nuovamente la struttura capsidica, contenente a sua volta al suo interno una copia di genoma virale neo sintetizzato. Questo assemblaggio non avviene a caso in quanto questo processo è intrinseco dell’informazione genica del virus (la capacità dell’autoassemblaggio). Quindi prima verranno raccolte all’interno della cellula tutte le subunità, i CAPSOMERI e poi, nel momento in cui tutto è pronto si procedere con l’AUTO-ASSEMBLAGGIO (=formazione della struttura CAPSIDICA) e poi abbiamo la fase importante dell’IMPACCAMENTO (entrata della copia di genoma virale all’interno della struttura capsidica). 4 - Rilascio della progenie matura Con la replicazione virale (dopo impaccamento ovviamente), vi è la lisi della cellula ospite e quindi il consequenziale rilascio di tutta questa progenie matura. (immagine sopra) Immagine tipica di un fago con struttura complessa (come fagi T), che hanno la struttura organizzata con la testa capsidica (contenente il genoma virale), tutta la coda di guaina contrattile, la piastra basilare
In effetti, durante questa fase di latenza viene anche definito, in maniera più adeguata, come PERIODO DI ECLISSI, ovvero un periodo in cui fenotipicamente non succede nulla ma da un punto di vista molecolare dell’infezione ci si sta preparando per effettuare il successivo burst, ovvero il momento in cui non ci sono virioni in circolo, perché il virus già sta facendo in questa fase i primi step del suo ciclo replicativo. Quindi questa fase di apparente “silenzio” è la fase che include i primi enzimi, fase dell’absorbimento, replicazione dell’acido nucleico, proteine capsidiche e inizia l’assemblaggio, con successivo rilascio, quando tutto è pronto, quando si è formata progenie matura (struttura capsidica pronta, entra la copia di materiale genico e poi sarà pronto per la lisi, ovvero per il burst). Il GENOMA VIRALE, esattamente come per quello batterico, può essere estremamente variabile in quanto a dimensioni, dalle dimensioni più piccolead esempio del virus MS2 di circa 3500 pb (più semplice e piccolo) fino ad altri molto più complessi. Anche qui, andando a vedere le dimensioni e la struttura del fago ci rendiamo conto della sua complessità e del suo relativo potere patogeno. Più è grande il genoma virale più è in grado di avere strategie alternative. Con questa variabilità possiamo avere virus con anche soli 4 geni (come il virus MS2), in grado comunque di replicarsi e anche di prendere il sopravvento su una cellula contenente anche 40.000 geni e distruggerla. Quindi il GENOMA VIRALE può essere con il massimo della VARIABILITA’. Infatti i virus possono avere genoma a DNA o a RNA, a SINGOLA o a DOPPIA ELICA, LINEARE o CIRCOLARE , UNICO o FRAMMENTATO. Hanno anche un’altra caratteristica, che gli deriva dal fatto che tutto il loro materiale genetico deve riuscire a rientrare all’interno della struttura capsidica (limitata quindi come capienza), di avere i GENI SOVRAPPOSTI, e quindi un potenziale di informazione altamente compresso (in questo modo hanno un’economia delle dimensioni ma riescono comunque a fare tante funzioni). Inoltre il GENOMA ad RNA a SINGOLA ELICA viene indicato a sua volta a POLARITA’ + (positiva) o a POLARITA – (negativa), rispetto al mRNA. Per convenzione infatti RNA+ (polarità positiva) ha lo stesso orientamento dell’mRNA per cui può fungere esso stesso da messaggero e iniziare subito la traduzione ; mentre RNA- (polarità negativa) è complementare all’mRNA. Ci sono dei virus che possono anche avere dei GENOMI AMBISENSO, cioè un misto di + e -. Le varie caratteristiche del genoma virale definiscono le diverse strategie di replicazione, e quindi le varie CLASSI REPLICATIVE VIRALI.
Infatti nel 1975 Baltimora elaborò la classificazione dei virus in 7 CLASSI REPLICATIVE VIRALI, basate proprio sul tipo di acido nucleico (genoma virale) che determina infatti la strategia replicativa: àCLASSE I: deossiribovirus (virus a DNA) con DNA a DOPPIA ELICA (Adenovirus, Herpesvirus, Poxvirus). àCLASSE II: deossiribovirus (virus a DNA) con DNA a SINGOLA ELICA (Parvovirus). àCLASSE III: ribovirus (virus a RNA) con RNA a DOPPIA ELICA (Reovirus*). àCLASSE IV: ribovirus (virusa a RNA) con RNA a POLARITA’ + (Picorravirus; Togavirus). Questa singola elica di RNA+ può fungere nella cellula direttamente da mRNA (in quanto ha lo stesso orientamento dell’mRNA). àCLASSE V: ribovirus (virus a RNA) con RNA a POLARITA’ – * (Orthomyxovirus, Rhobdovirus). Questa singola elica di RNA- funge da stampo per la sintesi dell’mRNA (dato che è complementare all’mRNA). Questi virus infatti hanno l’enzima RNApol RNA-dipendente associato al virione già. àCLASSE VI: ribovirus (virus a RNA) con RETROSCRIZIONE (->RETROVIRUS). àCLASSE VII: deossiribovirus (virus a DNA) con RETROSCRIZIONE. *I REOVIRUS e alcune famiglie di VIRUS a RNA- hanno un GENOMA con SEQUENZE INDIPENDENTI di RNA, frammentato (ad esempio il virus influenzale con 8 frammenti; bunyavirus con 3 frammenti; Reovirus con 12 frammenti). Per quanto riguarda i VIRUS con genoma a DNA, sono detti DEOSSIRIBOVIRUS e normalmente presentano un DNA a doppia elica con un filamento unico e lineare. Alcuni virus (come Papovavirus e Hepadnavirus) contengono un DNA circolare invece, e altri (come il Parvovirus) hanno un DNA monocatenario. La grandezza del genoma dà a sua volta una misura della complessità del virus e della sua replicazione, molto più evidente nei virus a DNA oltrettutto rispetto ai virus ad RNA. Nonostante tutte le differenze nella struttura del loro genoma, anche i virus obbediscono al DOGMA CENTRALE della biologia molecolare, per cui l’informazione genetica fluisce dall’acido nucleico (a DNA/RNA) alla proteina (anche perché i virus sfruttando i macchinari delle cellule eucariotiche o procariotiche , per cui loro avranno sempre uno step diverso per riportarsi a livello dell’mRNA, che poi procederà allo stesso modo: infatti ad esempio se partiamo da un RNA-, sappiamo che deve avere poi un mRNA che poi darà le proteine). E inoltre, a prescindere dalla natura del genoma, la traduzione in proteina di questo deve sempre passare per la forma ad mRNA.
La CLASSSE VI invece comprende un gruppo particolare di virus, ovvero i RETROVIRUS i quali hanno un genoma come RNA+ che non utilizzano direttamente come mRNA in quanto seguono una stretegia diversa; infatti necessitano di un intermedio a DNA (detto cDNA) per poter fare poi il loro mRNA. Quindi dall’RNA+ viene trascritto una copia in DNA, detta per questo cDNA (grazie alla trascrittasi inversa che si portano loro), che porteranno da singolo a doppio filamento e da questo intermedio a doppio filamento otterranno l’mRNA. Questo comunque serve anche perché quando loro sono a doppio filamento è più facile potersi integrare e interferire sul genoma della cellula ospite. La CLASSE VII infine comprende virus sempre con genoma a DNA a DOPPIO FILAMENTO, che però in alcuni punti diventa a SINGOLO FILAMENTO, cioè non ha un genoma interamente uguale, omogeneo, ma presenta tratti a doppio filamento e tratti a singolo filamento. Allora dal DNA viene rapportato in RNA+ che, può servire anche da messaggero ma per duplicare il suo DNA ed essere di nuovo impacchettato, da questo RNA farà un DNA e poi impacchetta quello, cioè fa un intermedio di DNA, per cui anche questo sfrutta una trascrittasi inversa. Quindi DNA-> RNA+à RNA->DNA àDNA doppio filamento. Curiosità per quanto riguarda il VIRUS DELLA STOMATITE VESCICOLARE (VSV). E’ un virus che utilizza presenta un genoma ad RNA-, e questo già ci fa capire un po’ che infezione fanno normalmente: abbiamo un RNA-, un’RNApol (che il virus si porta dietro) che copia l’RNA- in RNA+, il quale servirà da una parte per tradurre e dare proteine virus-specifiche, dall’altra parte fungerà da stampo per la formazione di tanti altri filamenti a polarità negativa, quindi a fini replicativi. Quindi all’inizio abbiamo un RNA- il quale viene copiato, per mezzo dell’RNApol RNA-dipendente, in un’RNA+ che viene in un certo senso segmentato, frammentato in quanto serve come sistema per la traduzione, vengono tradotti tutti i diversi enzimi etc.. ma contemporaneamente una parte di questo RNA+ servirà da stampo per fare l’mRNA complementare che poi verrà impacchettato. Altra strategia è quella fatta nel gruppo dei POLIOVIRUS, i quali presentano genoma con RNA+ , il quale viene tradotto direttamente come unico filamento (essendo RNA+ funge direttamente da mRNA), ovvero l’mRNA viene tradotto in un’unica lunga poliproteina, utilizzando come punto di riconoscimento a livello ribosomiale una proteina PBG. Una volta iniziato questo processo di traduzione e che si è ottenuto un’unica poliproteina questa viene segmentata nelle diverse strutture, cioè in tanti prodotti diversi funzionali (come enzimi fondamentali per il taglio della stessa proteina, strutture capsidiche, tutti gli enzimi dell’RNApol). Un’altra strategia per infettare le cellule eucariotiche.
I BATTERIOFAGI
Faremo i cicli del Φ X174 (con DNA a singolo filamento, ed è il primo fago ad essere stato sequenziato), dell’MS2 che è il più semplice in assoluto (in quanto è un fago piccolino con singola elica di RNA+). Poi tra i virus con genoma a DNA a doppio filamento attenzioneremo tutto il ciclo completo del fago LAMBDA e poi il modello del FAGO T (dispari e pari). (immagine a sx) struttura complessa della particella virale fagica. (immagine a dx) il fago lambda presenta la stessa struttura ma manca di tutta la parte finale/basale della coda. (immagine a sx) questo è un fago filamentoso che si chiama M13, uno di quelli che ha più impatto in ambito biotecnologico. Il FAGO LAMBDA presenta una testa icosaedrica di 64 nm, una coda non contrattile di 150 nm e, per quanto riguarda il contenuto genomico, ha un DNA a doppio filamento di circa 48 pb e una peculiarità è quella di avere alle 2 estremità della sequenza genomica delle sequenze COS (SITI COS) fra loro fortemente compatibili, e si chiamano proprio COS perché sta per sequenze