Baixe Farmacobotânica e outras Notas de estudo em PDF para Bioquímica, somente na Docsity!
Fotos da capa, da esquerda para a direita:
- Baccharis trimera (Less.) DC.
- Emilia fosbergii Nicolson
- Mikania involucrata Hook. & Arn.
- Mikania cordifolia (L.f.) Willd.
- Vernonia scorpioides (Lam.) Pers.
- Baccharis singularis (Vell.) G.M. Barroso
- Sphagneticola trilobata (L.) Pruski
- Barrosoa betoniciformis (DC.) R.M. King & H. Rob.
- Crepis japonica (L.) Benth.
- Piptocarpha leprosa (Less.) Baker
- Centratherum punctatum Cass.
- Achyrocline satureoides (Lam.) DC.
Fotos: 1, 8-9: R.S. Bianchini; 2, 6, 12: F.O. Souza, 3-5: S.E. Martins, 7: A.L. Santos, 10-11: R.P. Romanini. Layout da capa: Anderson Luiz dos Santos
Ficha Catalográfica elaborada pela Seção de Biblioteca do Instituto de Botânica
Souza, Fátima Otavina de
S729a Asteraceae no Parque Estadual da Ilha do Cardoso, Cananéia , SP/
Fátima Otavina de Souza -- São Paulo, 2007.
147p. il.
Dissertação (Mestrado) -- Instituto de Botânica da Secretaria de
Estado do Meio Ambiente, 2007
Bibliografia.
1. Asteraceae. 2. Mata Atlântica. 3. Florística. I. Título
CDU 582.
Comissão Julgadora:
Lucia Rossi Roberto Lourenço Esteves Prof(a). Dr(a). Prof(a). Dr(a).
Dra. Rosangela Simão Bianchini Orientadora
AGRADECIMENTOS
Quero expressar os meus mais sinceros agradecimentos a todos que contribuíram de alguma forma para que este trabalho tenha sido desenvolvido e meu objetivo alcançado, em especial:
Ao Instituto de Botânica, na pessoa do ex-diretor geral Dr. Luiz Mauro Barbosa pela utilização das instalações durante o desenvolvimento deste trabalho.
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), pela concessão da bolsa de mestrado (processo 04/11499-7 ).
Ao Instituto Florestal, órgão responsável pela unidade do Parque Estadual da Ilha do Cardoso, na pessoa do ex- diretor geral Dr. João Batista Baitello, por permitir a coleta do material botânico e a utilização da infra-estrutura na Ilha.
À minha querida orientadora, Dra. Rosangela Simão Bianchini, que me apresentou e incentivou a trabalhar com as maravilhosas Asteraceae, pela confiança, paciência, amizade e por me deixar caminhar quase que sozinha.
Aos coordenadores do programa de pós-graduação em biodiversidade vegetal e meio ambiente e aos funcionários e estagiários da secretaria de pós-graduação, especialmente a Márcia R. Ângelo e Antonio A. C. Borges, pela atenção e serviços prestados.
Aos curadores dos herbários visitados HRCB, IAC, SP, SPF, SPSF, UEC e do Instituto Plantarum, pela atenção e disponibilidade que me receberam e pelo empréstimo de materiais.
Aos membros da banca do exame de qualificação Fábio de Barros, Maria das Graças Lapa Wanderley e Mara Agelina Galvão Magenta, pelas críticas e valiosas sugestões.
A todos os professores das disciplinas cursadas, com os quais pude ampliar meus conhecimentos, em especial, Carlos E.M. Bicudo, Fábio de Barros, Jefferson Prado, Maria das Graças L. Wanderley, Tarciso Filgueiras, Renato Mello Silva, Rosangela S. Bianchini e Sergio Romaniuc Neto.
Aos funcionários e pesquisadores da seção de curadoria do herbário, Ana Célia Calado, Cintia Kameyama, Eduardo L.M. Catharino, Fábio de Barros, Claudinéia Inácio, Georgete J. Silva, Gerlene L. Esteves, Inês Cordeiro, Jefferson Prado, Letícia R. Lima, Lucia Rossi, Lurdes Indrigo, Maria Cândida H. Mamede, Maria das Graças L. Wanderley, Maria Margarida R.F. Melo, Marie Sugiyama, Mizué Kirizawa, Tereza Yano, Regina T. Shirasuna, Sergio Romaniuc Neto e Sonia Aragaki, por estarem sempre dispostos a nos ajudar e pelos momentos agradáveis compartilhados durante todos esses anos, não apenas durante o mestrado.
Ao Jefferson Prado e Cintia Kameyama, pelo auxílio e sugestões na citação do cabeçalho das espécies.
Aos Sinanterólogos Mara A.G. Magenta, João Semir e Roberto L. Esteves, pelas dicas, auxílio e sugestões de bibliografia.
Aos funcionários da biblioteca Jéferson A. de Souza, Maria Helena S.C.F. Gallo e Sueli P. de Caldas e a estagiária Tais C. da Silva, pela prestatividade e atenção com a qual sempre me atenderam.
Às funcionárias do Xerox Maria R.P. Oliveira, Sirléia C. Ferreira e Rosalina de Mattos, pelos trabalhos prestados durante esses anos.
Aos motoristas Luiz Zanqueta e Aliomar O. Gomes (Mazinho) por terem nos conduzido até Cananéia, sempre com muito profissionalismo e simpatia.
Às amigas Marie Sugyama e Suzana E. Martins pessoas maravilhosas, pelo apoio que me deram, pelas viagens de coleta, companhia, auxílio, conselhos e sugestões e, principalmente pela amizade.
À Margarida pelo profissionalismo, convivência agradável, amizade, por sempre nos ajudar e especialmente pela disponibilidade em ler a dissertação e pelas sugestões.
RESUMO
Este trabalho é o resultado do levantamento florístico das Asteraceae no Parque Estadual da Ilha do Cardoso, localizado no município de Cananéia, extremo sul do Estado de São Paulo, entre as coordenadas 25°03’05’ – 25°18’18’S e 47°53’48’ – 48°05’42’W. O estudo foi baseado na análise de 218 exsicatas provenientes da Ilha do Cardoso e observações das espécies em campo. Foram amostradas 51 espécies, distribuídas em 29 gêneros, pertencentes às subfamílias Asteroideae e Cichorioideae. A maioria dos gêneros amostrados, ou seja , 79% apresentou apenas uma espécie e os gêneros com mais de um representante foram: Mikania (13 spp.), Baccharis (4 spp.), Vernonia (4 spp.), Piptocarpha (3 spp.), Pterocaulon (2 spp.) e Symphyopappus (2 spp.). Além de contribuir para o conhecimento da flora local, este trabalho auxiliará também no conhecimento das Asteraceae para a Flora Fanerogâmica do Estado de São Paulo. Foram elaboradas chaves de identificações, descrições e ilustrações para os gêneros e espécies, comentários sobre distribuição geográfica, importância econômica, nomes populares, além de outras informações relevantes.
Palavras chave: Astereae, Florística, Mata Atlântica, Ilha do Cardoso.
ABSTRACT
This work presents a floristic survey of the Asteraceae in the “Parque Estadual da Ilha do Cardoso”, located at Cananéia town, in the southernmost of the state of São Paulo, between 25°03’05’ – 25°18’18’S and 47°53’48’ – 48°05’42’W. This study was based on the analysis of 218 herbarium sheets from “Ilha do Cardoso”, and the observations of the species in the field. Fifty one species were sampled, distributed in 29 genera, belonging to subfamilies Asteroideae and Cichorioideae. Most of the sampled genera, corresponding to 79%, is represented only by a single species, and the genera with more than one species are: Mikania (13 spp.), Baccharis (4 spp.), Vernonia (4 spp.), Piptocarpha (3 spp.), Pterocaulon (2 spp.), and Symphyopappus (2 spp.). Besides contributing to the knowledge of the local flora, this research will also improve the knowledge of the Asteraceae for the study of Phanerogamic Flora of the State of São Paulo project. The text includes not only wild plants, but also weeds species. It provides scientific and common names, taxonomic descriptions, identification keys, illustrations, geographic ranges, economic importance, and other relevant biological information.
Key words: Astereae, Floristic, Atlantic Forest, Ilha do Cardoso.
SUMÁRIO
ÍNDICE DE FIGURAS
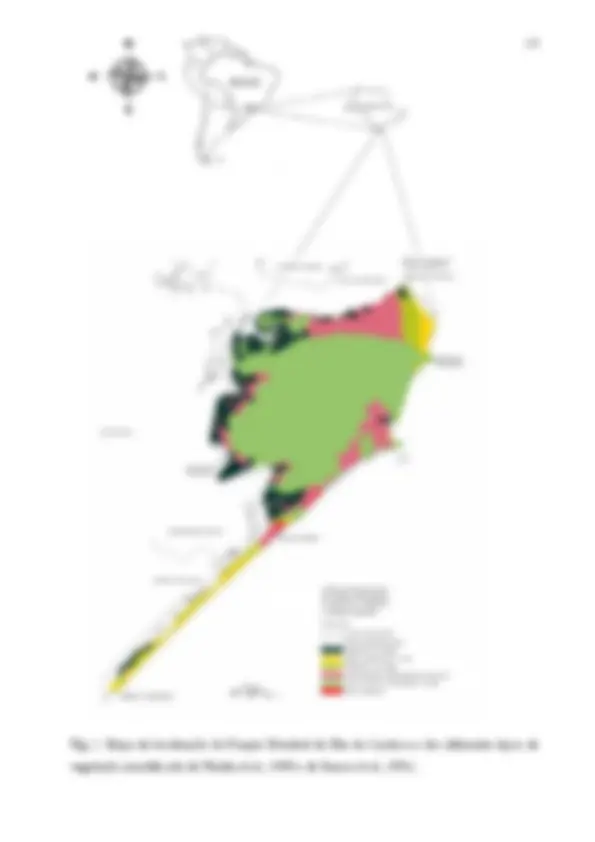
Fig. 1. Mapa de localização do Parque Estadual da Ilha do Cardoso e os diferentes tipos de vegetação ......................................................................................................................................................... 15
Fig. 2. Prancha de Fotos dos diferentes tipos de vegetação .................................................................... 16
Fig. 3. Prancha de Fotos de Achyrocline satureoides (Lam.) DC.; Baccharis singularis (Vell.) G.M. Barroso; Barrosoa betoniciformis (DC.) R.M. King & H. Rob.; Centratherum punctatum Cass.; Conyza bonariensis (L.) Cronquist; Emilia fosbergii Nicolson .............................................................................................................. 43
Fig. 4-8. Ilustrações de Baccharis brachylaenoides DC. ................................................................................... 44
Fig. 9-13. Ilustrações de Baccharis schultzii Baker. ....................................................................................... 44
Fig. 14-18. Ilustrações de Baccharis singularis (Vell.) G.M. Barroso .......................................................... 44
Fig. 19-22: Ilustrações de Baccharis trimera (Less.) DC. ............................................................................. 44
Fig. 23-25. Ilustrações de Mikania biformis DC. .......................................................................................... 87
Fig. 26-29. Ilustrações de Mikania cordifolia (L.f.) Willd. ........................................................................... 87
Fig. 30-32. Ilustrações de Mikania hastato - cordata Malme. ......................................................................... 87
Fig. 33-38. Ilustrações de Mikania hoehnei B.L. Rob. ................................................................................. 87
Fig. 39-42. Ilustrações de Mikania involucrata Hook. & Arn. .................................................................... 87
Fig. 43-45. Ilustrações de Mikania laevigata Sch. Bip. ex Baker ............................................................... 87
Fig.46. Prancha de Fotos de Mikania biformis DC.; Mikania cordifolia (L.f.) Willd.; Mikania hoehnei B.L. Rob.; Mikania involucrata. Hook. & Arn.; Piptocarpha leprosa (Less.) Baker .............................................. 88
Fig. 47-50. Ilustrações de Mikania lundiana DC. ........................................................................................ 89
1. INTRODUÇÃO
Considerada uma das maiores famílias, representando cerca de 1/10 de todas as
Angiospermas, Asteraceae possui aproximadamente 23.000 espécies distribuídas em 1.
gêneros (Bremer, 1994).
Possui distribuição cosmopolita sendo encontrada em todos os continentes, com
exceção do Antártico, e seus representantes normalmente apresentam adaptações às várias
condições ambientais e ecológicas (Barroso et al., 1991; Judd et al., 1999).
A América Latina é considerada um centro de diversidade de Asteraceae, sendo que
na América do Sul, em algumas regiões semi-áridas e na Patagônia, os representantes de
Asteraceae correspondem a 20% da flora (Cabrera, 1978). Para o Brasil são estimados
aproximadamente 180 gêneros e 3.000 espécies (Hind, 1993) com distribuição desde as
regiões mais frias e úmidas, como as serras do Sudeste e Sul, até as áreas secas na região do
semi-árido nordestino (Pereira, 1989), sendo menos freqüentes em formações florestais
(Hind, 1993).
São plantas com hábito variável abrangendo desde ervas até arbustos, mais raramente
árvores; podem ser monóicas, dióicas ou poligamo-dióicas. As folhas são alternas ou
opostas, raramente verticiladas, pecioladas ou sésseis, simples, inteiras, lobadas ou dissectas,
distribuídas ao longo dos ramos, em rosetas basais ou ausentes; estípulas geralmente
ausentes.
O indumento é variável, constituído de tricomas multicelulares unisseriados ou
multisseriados, composto de células uniformes ou células basais e terminais modificadas.
Geralmente apresentam tricomas tectores que podem ser simples, estrelados, malpiguiáceos
ou escamiformes, também são comuns tricomas glandulares depressos na face abaxial das
folhas, e neste caso, aparecem como pontos pretos, translúcidos ou estrias alongadas. Sua
secreção é responsável pela aparência envernizada das folhas de alguns gêneros, em especial
de Baccharis L., Eupatorium L. sensu lato , Gochnatia Kunth e Symphyopappus Turcz. (Metcalfe &
Chalk, 1950). Tricomas unicelulares são raros, sendo exclusivos da subfamília
Barnadesoideae (Bremer, 1994).
Quanto à anatomia, a principal característica é a ocorrência de canais laticíferos e
resiníferos, sendo que sua distribuição nos membros da família tem valor taxonômico, uma
vez que canais ou cavidades de látex são característicos de Cichorioideae, especialmente na
tribo Lactuceae, enquanto que canais ou cavidades de resinas são comuns nos representantes
de Asteroideae (Bremer, 1994).
Uma das principais características das Asteraceae é a diversidade de substâncias
químicas, especialmente os polifrutanos inulinas e as lactonas sesquiterpênicas, além de óleos
voláteis terpenóides e algumas classes de alcalóides, principalmente alcalóides pirrolizidínicos
característicos do gênero Senecio L., estando ausentes os iridóides e taninos (Cronquist, 1981).
A diversidade estrutural dos terpenos, em especial das lactonas sesquiterpênicas e dos
furanosesquiterpênos levou Seaman (1982) a fazer um estudo analisando as variedades destes
terpenos na família e concluiu que estes compostos podem ser aplicados nos diferentes
níveis taxonômicos, podendo auxiliar nas análises cladísticas, salientando, entretanto, que
para delimitar espécies não são muito seguros, uma vez que é comum a ocorrência de
“quimiotipos”, podendo ser úteis no reconhecimento de ecotipos, variedades ou
subespécies.
Asteraceae é facilmente reconhecida pelas flores reunidas em capítulos, formando um
pseudanto, que podem ser isolados ou agrupados nas mais diversificadas inflorescências,
formando capitulescências corimbiformes, paniculiformes, tirsóideo-paniculadas,
racemiformes, espiciformes, fasciculadas, glomeruliformes ou até mesmo formando
capítulos de capítulos envoltos por invólucro secundário.
Os capítulos podem ser classificados quanto ao sexo e à morfologia das flores
(adaptado de Bremer, 1994): Quanto ao sexo, podem ser heterógamos (constituídos de
flores monoclinas e flores diclinas, femininas ou neutras) e homógamos (compostos apenas
por flores monoclinas ou flores diclinas). Com relação à morfologia floral, são classificados
em disciformes (formados por dois tipos de flores, marginais filiformes e centrais tubulosas),
discóides (constituídos por flores tubulosas, ou flores liguladas, ou bilabiadas) e radiados
(com flores do raio liguladas e flores do disco tubulosas).
Em torno do receptáculo estão dispostas as brácteas involucrais, que apresentam
morfologia e número variados, sendo constante em alguns gêneros, como em Mikania Willd.,
que apresenta sempre quatro. As brácteas involucrais normalmente são livres entre si,
podendo ser parcial ou totalmente conatas, formando um invólucro gamofilo como em
Xanthium L. O receptáculo pode ser plano, cônico, côncavo, convexo ou cilíndrico;
epaleáceo ou paleáceo.
O cálice em Asteraceae é altamente modificado em pápus, normalmente cerdosos,
constituído de cerdas escabras, barbeladas ou plumosas; quando as cerdas são largas e
geralmente aplainadas, são denominadas cerdas aristadas; quando se apresentam mais largas
do que longas são escamas; quando as cerdas são curtas e conatas o pápus é coroniforme, ou
ainda o pápus pode estar ausente. Os elementos dos pápus podem estar dispostos em uma
ou mais séries e serem persistentes ou caducos na maturação do fruto (Bremer, 1994). O
pápus é essencial na dispersão do diásporo e sua morfologia está diretamente relacionada à
dispersão. Desta forma, pápus cerdosos e plumosos estão relacionados à anemocoria,
enquanto os aristados à epizoocoria (Magenta, 1998).
Provavelmente a maior contribuição das Asteraceae para o homem é a ecológica, uma
vez que são responsáveis pela grande diversidade de espécies e, portanto, favorecem a
estabilidade e sustentabilidade produtiva das vegetações em áreas secas do mundo, tais como
os cerrados, capoeiras, campos e semi-áridos, especialmente nas regiões tropicais e
subtropicais (Heywood, 1979). Economicamente são pouco utilizadas na alimentação; têm
potencial medicinal, toxicológico e alergogênico, destacando-se sem dúvida como planta
ornamental ou daninha.
Os caracteres ecológicos que distinguem as Asteraceae das demais famílias são os
pseudantos, o mecanismo especializado de apresentação do pólen e a diversidade de armas
químicas presentes em todos os membros da família (Cronquist, 1981). Por isso, alguns
autores (Cronquist, 1981; Bremer, 1994) sugeriram que o grande sucesso de Asteraceae deva-
se a associação dos compostos químicos com a morfologia especializada, pois apenas a
morfologia especializada, como as flores reunidas em capítulos e o mecanismo secundário de
apresentação do pólen também são encontrados em outras famílias, porém estas não
apresentam a grande diversidade de Asteraceae.
Mesmo sendo um táxon com numerosas espécies, Asteraceae sempre foi considerada
como um grupo natural bem sustentado por caracteres sinapomórficos, tais como: lactonas
sesquiterpênicas, cálice altamente modificado em pápus, anteras conatas com deiscência
introrsa formando um tubo ao redor do estilete; mecanismo secundário de apresentação de
pólen com estilete modificado funcionando como uma escova; ovário bicarpelar com apenas
um óvulo de placentação basal e fruto com endosperma constituído de apenas uma camada
inconspícua (APG II, 2003; Barkley et al., 2006).
Estudos sugerem que as Asteraceae tiveram origem na América do Sul, no Eoceno
Superior com posterior migração para a América do Norte (DeVore & Stuessy, 1995).
A família está posicionada na Ordem Asterales, normalmente sozinha ou junto com
outras pequenas famílias. Cronquist (1988) considerou Asteraceae como única família da
Ordem Asterales, relacionado-a com as Ordens Gentianales, Rubiales, Dipsacales e
Calycerales. Atualmente, estudos filogenéticos baseados em dados morfológicos e
moleculares, como os de APG II (2003), corroboram o posicionamento de Asteraceae em
Asterales juntamente com outras dez famílias. Os caracteres sinapomórficos que sustentam a
posicionamento dessas famílias na ordem são os botões florais com deiscência valvar,
presença de parênquima paratraqueal, substâncias químicas como oligo ou polifrutanos,
ácido elágico e mecanismo especializado de apresentação do grão de pólen. Dentre essas
famílias, Asteraceae emerge juntamente com Menyanthaceae, Goodeniaceae e Calyceraceae,
formando um grupo monofilético sustentado pela presença de ácido caféico, elementos de
vasos com perfurações simples e corola conata em tubo com fusão marginal e, dentro desse
clado, Calyceraceae aparece como grupo irmão de Asteraceae, ambas compartilhando de
cálice modificado e persistente, tipo de nervação da corola, ovário bicarpelar com apenas um
óvulo, pólen com depressão intercolpar e fruto do tipo cipsela (APG II, 2003; Barkley et al.,
O posicionamento inframiliar também não sofreu grandes modificações nos últimos
100 anos, uma vez que Cassini (1819) foi um dos primeiros autores a considerar subdivisões
em Asteraceae, utilizando como caracteres diagnósticos para o reconhecimento das tribos a
morfologia do estilete e das anteras. Esse autor, baseando-se nessas características, descreveu
19 tribos, das quais 13, mesmo após 176 anos e com os diversos estudos, não sofreram
grandes modificações, sendo aceitas no trabalho de Bremer (1994), ainda um dos mais atuais.
Bentham (1873) tendo como base os estudos de Cassini (1819), Lessing (1829, 1830,
1831) e De Candolle (1836-1838), separou as Asteraceae em duas subfamílias e 13 tribos.
Essa classificação foi bem aceita e considerada a base da sistemática sobre a qual a família
tradicionalmente é colocada. Hoffmann (1894) realizou algumas modificações nessa
classificação, mas reconheceu as subdivisões de Bentham, sendo esse tratamento adotado
por muito tempo nos trabalhos com Asteraceae. Em 1977, Heywood et al. fizeram um
estudo geral com as Asteraceae, propuseram algumas modificações e o reconhecimento de
17 tribos.
Bremer & Jansen (1992) publicaram um estudo filogenético baseado em dados
morfológicos e moleculares com DNA do cloroplasto. Neste estudo, excluíram a subtribo
Barnadesiine de Mutisieae, elevando-a a categoria de subfamília. Dois anos depois, a primeira
Tribo, King & Robinson publicaram numerosos trabalhos, a grande maioria sobre gêneros
ou espécies novas, utilizando como características diagnósticas especialmente os
microcaracteres. Esses trabalhos foram posteriormente sintetizados por King & Robinson
(1987) no livro “The genera of the Eupatorieae”. Com a criação dos novos gêneros e o
restabelecimento de outros, a tribo Eupatorieae, anteriormente constituída de 32 gêneros,
passou a abrigar 180 gêneros, dos quais 99 eram novos. Eupatorium L. sensu lato foi o gênero
que sofreu mais divisões, sendo segregado em 80 novos gêneros, muitos dos quais
monoespecíficos. Desta forma, Eupatorium , um gênero tradicionalmente representado por
1.200 espécies, passou a ser composto de apenas 45 espécies praticamente restritas à
América do Norte, com algumas exceções na América Central, Antilhas e Ásia (King &
Robinson, 1987; Esteves, 2001).
Outro táxon que também passou por modificações durante esses anos foi
Vernonieae. Em 1999, Robinson publicou um trabalho propondo uma nova classificação
para as Vernonieae americanas, esse estudo foi baseado nos vários trabalhos realizados por
ele e outros autores durante os anos de 1973 a 1999. A principal modificação ocorreu no
gênero Vernonia que foi segregado em outros, como por exemplo, Lessingianthus H. Rob. (
spp.), Vernonanthura H. Rob. (65 spp.) e Cyrtocymura H. Rob. (6 spp.). O gênero Vernonia
passou a abrigar 500 espécies, ficando representado no Novo Mundo apenas por espécies da
América do Norte e Vernonieae, tradicionalmente constituída de 72 gêneros, passou a
abrigar 98, dos quais 17 eram novos.
Apesar dos inúmeros trabalhos taxonômicos realizados no mundo todo com
Asteraceae, quando levado em consideração o tamanho da família, estes ainda são
insuficientes. Dentre esses trabalhos, destacam-se algumas revisões de tribos, como por
exemplo: Senecioneae (Nordenstam, 1978), Heliantheae (Robinson, 1981), Liabeae
(Robinson, 1983), Eupatorieae (King & Robinson, 1987), Inuleae (Anderberg, 1989, 1991),
Mutisieae (Hansen, 1991) e Astereae (Nesom, 1994). Em relação aos gêneros, citam-se as
revisões de Bidens L. (Sherff, 1937), Pentacalia Cass. (Robinson & Cuatrecasas, 1978),
Centratherum Cass. e Phyllocephallum Blume (Kirkman, 1981), Piptocarpha R. Br. (Smith, 1984),
Lychnophora Mart. (Semir, 1991) e Pluchea Cass. (King-Jones, 2001).
Para o Brasil, dentre os trabalhos de cunho geral, destacam-se os de Baker (1873,
1876, 1882, 1884) para a Flora Brasiliensis, além de levantamentos florísticos para
determinadas áreas, como a Flora Paulista (Loefgren, 1897), Flora do Itatiaia e do Rio de
Janeiro-RJ (Barroso, 1957; 1959), de Mucugê-BA (Harley & Simmons, 1986), Flora da Serra
do Cipó-MG (Leitão Filho & Semir, 1987), do Pico das Almas-Ba (Hind, 1995), do Parque
Estadual da Serra da Canastra-MG (Nakajima, 2000), do Parque Estadual das Fontes do
Ipiranga-SP (Nakajima et al., 2001), de Grão-Mogol-MG (Hind, 2003) e de Picinguaba-SP
(Moraes & Monteiro, 2006).
Além desses levantamentos, outros trabalhos de importância e que têm contribuído
para o conhecimento das Asteraceae no Brasil são os tratamentos taxonômicos para tribos e
gêneros, como: Aspilia Thou. (Santos, 2001), Baccharidinae (Barroso, 1976; Barroso & Bueno,
2002), Eupatorium L. (Barroso, 1950), Heliantheae (Pereira, 1989), Ichthyothere Mart. (Pereira,
2001), Lychnophora Mart. (Semir, 1991), Mikania Willd. (Barroso, 1958; Ritter, & Miotto,
2005.), Mutisieae (Cabrera & Klein, 1973; Mondin, 1996; Roque & Pirani, 1997), Senecio L.
(Cabrera, 1957), Senecioneae (Cabrera & Klein, 1975; Matzenbacher, 1998), Stevia Cav.
(Monteiro, 1982), Stilpnopappus Mart. ex DC. (Esteves, 1993) e Trichogonia Gardn. (Barroso,
A família está representada no Estado de São Paulo, de acordo com levantamento no
banco de dados da “Flora Fanerogâmica do Estado de São Paulo”, por cerca de 160 gêneros
e 635 espécies e, apesar da grande representatividade, são poucos os trabalhos taxonômicos
ou florísticos, destacando-se além de alguns já citados, os de Leitão Filho (1972) com as
Vernonieae, de Magenta (1998) com Ambrosiinae, Galinsoginae e Coreopsidnae e de
Esteves (2001) com Eupatorium sensu lato ; bem como as listagens de Asteraceae para o Parque
Estadual da Ilha do Cardoso (Leitão Filho, 1991), para a Estação Ecológica Juréia-Itatins
(Semir, 2004) e para a Reserva Biológica do Alto da Serra de Paranapiacaba (Kirizawa, 2004).
O Parque Estadual da Ilha do Cardoso, local onde foi efetuado o levantamento das
Asteraceae, está localizado no extremo sul do Estado de São Paulo, no município de
Cananéia, divisa com o Estado do Paraná, entre as coordenadas 25°03’05’ – 25°18’18’S e
47°53’48’ – 48°05’42’W. Estudos científicos referentes à botânica na Ilha do Cardoso estão
sendo publicados desde 1979, sendo um dos pioneiros o de De Grande & Lopes (1981). O
projeto Flora Fanerogâmica da Ilha do Cardoso iniciou-se oficialmente em dezembro de
1982, entretanto as coletas estavam sendo realizadas muito antes; vários anos de coletas
sistemáticas resultaram em uma coleção com mais de 4.500 exsicatas, amostradas em
diversas áreas da Ilha (Barros et al., 1991). Em 1991 foi publicado o primeiro volume da
série “Flora Fanerogâmica da Ilha do Cardoso” e, desde então, 11 volumes foram
publicados, com 112 famílias já concluídas, restando apenas as grandes famílias como
Orchidaceae, Fabaceae, Myrtaceae e Asteraceae. A família Orchidaceae foi concluída por