Baixe Linfócitos B e imunoglobulinas e outras Notas de estudo em PDF para Imunologia, somente na Docsity!
C A P Í T U L O 8
LINFÓCITOS B
Os linfócitos são as únicas células do sistema imune que apresentam receptores específicos para os antígenos. No caso dos linfócitos B, os receptores de antígeno são moléculas de Imunoglobulina (Ig), que se encontram imersas na membrana da célula em repouso ( naive ). Estas Igs, após a ativação dos Linfócitos B por LT auxiliares, serão secretadas pelos plasmócitos, linfócitos B que se diferenciam em células secretoras.
A idéia de que as células secretoras de anticorpos possuem receptores de membrana surgiu, pela primeira vez, em 1900. O imunologista Paul Ehrlich lançou a hipótese de que uma única célula possui diferentes receptores com capacidade de reconhecer diferentes estruturas estranhas. A entrada de um determinado antígeno selecionaria qual destes inúmeros receptores seria produzido em maior concentração. Segundo esta teoria, se uma célula tiver receptores para os antígenos X, Y e Z, a entrada do antígeno Z aumentaria o número de receptores para este antígeno, reduzindo o número dos receptores para X e Y. Depois de um certo número de receptores para Z terem reconhecido vários destes antígenos, estas células passariam a secretar estes receptores, que seriam o que conhecemos, atualmente, como anticorpos. Esta teoria de Ehrlich ficou conhecida como a Teoria da Cadeia Lateral.
Por volta de 1957, surgiu a Teoria da Seleção Clonal, criada por Burnet. Esta teoria, que é a aceita atualmente, postula que cada linfócito B reconhece apenas um antígeno. Enquanto que a Teoria da Cadeia Lateral postula que uma célula secretora de anticorpos possui vários receptores para diferentes antígenos, a teoria da seleção clonal postula que cada linfócito B possui um único tipo de receptor. Ou seja, existe um linfócito B que reconhece o antígeno Z, um outro que reconhece o X e um terceiro, que reconhece o Y. A entrada do antígeno Z, faria com que somente as células com receptor para este antígeno proliferassem e passassem a secretar anticorpos específicos para o antígeno Z.
88 FILOMENA M. P. BALESTIERI - IMUNOLOGIA BÁSICA E APLICADA
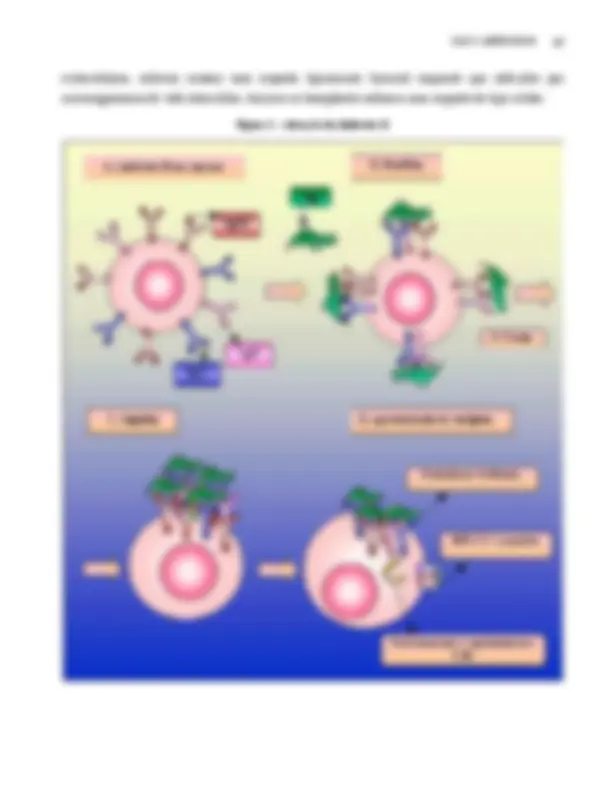
De acordo com a teoria da seleção clonal, cada linfócito B apresenta em sua membrana moléculas de Igs de diferentes classes, que reconhecem o mesmo antígeno (Figura 1A). Todos os linfócitos B possuem IgM e IgD de membrana; no entanto, alguns possuem além destas Igs, ou IgG ou IgA ou IgE. Um LB que expressa IgM, IgD e IgG na sua membrana produzirá, durante uma resposta imune primária (Capítulo 4) grandes concentrações de IgM e baixas concentrações de IgG. A partir do segundo contato com o mesmo antígeno (resposta secundária), este LB produzirá altas concentrações de IgG e baixas concentrações de IgM. Um LB que expressa IgM, IgD e IgA na sua membrana produzirá, durante uma resposta primária, grandes concentrações de IgM e baixas concentrações de IgA. A partir do segundo contato com o mesmo antígeno, este LB produzirá altas concentrações de IgA e baixas concentrações de IgM. Um mecanismo similar ocorre com células apresentando IgM, IgD e IgE de membrana. Esta mudança na expressão das classes de Igs (de IgM para IgG, IgA ou IgE) depende das moléculas (citocinas) produzidas pelos linfócitos T auxiliares que ativaram os linfócitos B (Capítulo 9).
Nos linfócitos B em repouso, estas Igs estão distribuídas de forma homogênea na superfície celular (Figura 1A). Quando estas Igs reconhecem um antígeno, estas moléculas aglutinam em regiões diferentes da célula, fenômeno ao qual se dá o nome de patching (Figura 1B). Após 5 a 15 minutos, estas Igs associadas ao antígeno aglomeram-se num dos pólos da célula (Figura 1C). A polarização das Igs na membrana é visualizada na imunofluorescência (Capítulo 20), para detectar as Igs de membrana, como um boné ( cap ), e por isso dá-se a este fenômeno o nome de capping. Após um período, que pode variar entre 5 a 60 minutos, ocorre a endocitose das Igs e do antígeno associado (Figura 1D). Este antígeno é processado no interior do linfócito B, num período de 1 a 6 horas, e peptídeos de 10 a 20 aminoácidos são associados a proteínas de classe II, presentes na membrana da célula (Capítulo 6). Desta forma, os linfócitos B tornam-se células apresentadoras de antígenos (APCs), da mesma forma que os macrófagos e as células dendríticas que após endocitarem antígenos apresentam peptídeos associados a proteínas de classe II (Capítulo 6). A apresentação de antígenos por moléculas do MHC-II confere aos linfócitos B, macrófagos e células dendríticas a capacidade de ativarem e serem ativadas por linfócitos T auxiliares (Capítulo 9). A ativação dos linfócitos B por LT auxiliares induz a proliferação e diferenciação destas células em plasmócitos ou linfócitos B de memória.
Quando ocorre um predomínio da ativação de linfócitos B durante uma resposta imune, diz-se que esta é uma resposta do tipo humoral. Por outro lado, quando a ativação dos linfócitos T é predominante chama-se esta resposta de celular. Normalmente, infecções por bactérias ou antígenos
90 FILOMENA M. P. BALESTIERI - IMUNOLOGIA BÁSICA E APLICADA
IMUNOGLOBULINAS Os linfócitos B possuem receptores de membrana que são moléculas de Igs de diferentes classes. Todas as Igs são formadas por 4 cadeias polipeptídicas, sendo que duas destas cadeias apresentam um PM mais alto, devido a sua estrutura que é composta de 450 resíduos de aminoácidos e são chamadas de cadeias pesadas e as outras duas, denominadas de cadeias leves, apresentam 212 resíduos de aminoácidos e por isso um PM mais baixo (Figura 2). Estas cadeias estão associadas entre si por pontes de dissulfeto que ocorrem quando duas cisteínas estão próximas e formam um tipo de estrutura globular que é característico das Igs. A estrutura das Igs, apresentada na figura 2, foi evidenciada por métodos de clivagem enzimática utilizando-se a pepsina e a papaína.
Figura 2 – Estrutura Básica de uma Imunoglobulina (IgG1)
CAP. 8 - LINFÓCITOS B (^91)
A papaína quebra a molécula de Ig em regiões acima da ponte de dissulfeto que associam as duas cadeias pesadas, originando três fragmentos: dois que se associam ao antígeno e são os Fab e um que se cristaliza que é o Fc (Figura 7.2). A pepsina quebra a molécula de Ig abaixo da ponte de dissulfeto que associa as duas cadeias pesadas originando dois fragmentos: os dois Fab, que continuam associados e por isso denominado Fab’ e o Fc (Figura 2).
Quando foram iniciados os estudos das seqüências de aminoácidos das Igs foram evidenciadas regiões em que estas seqüências mantinham um padrão constante e outras em que este padrão era variável. Destas análises surgiu também o conhecimento das diferentes classes ou isótipos de Igs. O que distingue cada uma destas classes é a seqüência de aminoácidos da região constante da cadeia pesada. Estas diferenças definem as 5 classes de imunoglobulinas: IgG, IgM, IgA, IgD e IgE cujas cadeias pesadas são chamadas, respectivamente, cadeia γ (gama), μ (mü), α (alfa), δ (delta) e ε (épsilon). Em relação à região constante das cadeias leves só existem dois tipos de seqüências que foram denominadas κ (kappa) ou λ (lambda). Uma molécula de IgG, por exemplo, sempre possuirá duas cadeias gama e duas kappa ou duas cadeias gama e duas lambda. Não existem moléculas de Ig expressando uma cadeia leve kappa e outra lambda.
Em relação à região variável das Igs, existem regiões hiper-variáveis que são complementares à seqüência de aminoácidos dos determinantes antigênicos presentes nos antígenos (Capítulo 5). As regiões hiper-variáveis da cadeia pesada estão presentes entre os resíduos 30-35, 50-62 e 94-102, formando o que se chama de CDR1 (Região determinante de complementariedade 1), CDR2 e CDR3, respectivamente. Nas cadeias leves o CDR1, CDR2 e CDR3 estão presentes, respectivamente, entre os seguintes resíduos de aminoácidos: 25-34, 50-60 e 89-97.
As principais interações químicas que ocorrem entre as regiões hiper-variáveis das moléculas das Igs e os determinantes antigênicos são do tipo ligação de van der Waals, forças eletrostáticas e hidrofóbicas e pontes de hidrogênio.
CAP. 8 - LINFÓCITOS B (^93)
Figura 3 – Estrutura das subclasses de IgG humana
IgM A IgM é uma imunoglobulina presente nos líquidos internos do corpo e nas secreções externas, correspondendo a 10% do total das Igs séricas. É encontrada em duas formas estruturais: monômeros e pentâmeros.
A concentração média da IgM no soro é de 1,5 mg/ml, em seres humanos adultos. A vida média da IgM sérica é de 10 dias. O Peso Molecular da IgM pentamérica é de 970 kDa. A IgM possui cerca de 12% de carboidratos em sua estrutura. Faz parte também da sua estrutura uma cadeia J (junção) que serve para manter unidas as moléculas que formam o pentâmero.
A IgM é produzida em altas concentrações na resposta imune primária e em baixas, na resposta imune secundária. A sua principal atividade biológica é a ativação do sistema complemento pela via clássica, levando o agente agressor à lise. A IgM é produzida principalmente na resposta a antígenos T - independentes, que são geralmente polissacarídeos complexos. A sua afinidade é baixa para estes
94 FILOMENA M. P. BALESTIERI - IMUNOLOGIA BÁSICA E APLICADA
antígenos e durante a resposta imune primária, a produção de IgM é trocada pela produção de IgG, IgA ou IgE, que apresentam maior afinidade para antígenos proteícos.
IgA
A IgA é uma imunoglobulina presente principalmente nas secreções externas (saliva, colostro, lágrimas, secreções genitourinárias e secreções traqueobronquiais) correspondendo a 15-20% do total das Igs séricas.
É encontrada em duas formas estruturais: monômeros e dímeros e em duas subclasses IgA1 e IgA2, na espécie humana. A concentração média da IgA1 no soro é de 3,0 mg/ml e a da IgA2 é de 0, mg/ml, em seres humanos adultos. A vida média da IgA sérica é de 6 dias. O Peso Molecular da IgA monomérica é de 160 kDa. A IgA possui cerca de 7-11% de carboidratos em sua estrutura. Faz parte também da sua estrutura uma cadeia J (junção), que mantém os dímeros unidos, e um componente secretório (CS). O CS é produzido por células da mucosa próximas aos centros germinativos onde os linfócitos B estão proliferando e sintetizando IgA (Figura 4).
A IgA é produzida em altas concentrações na resposta imune secundária e em baixas, na resposta imune primária. A sua principal atividade biológica é a neutralização de antígenos na superfície das mucosas. A IgA pode ativar o sistema complemento pela via alternativa (Capítulo 10).
96 FILOMENA M. P. BALESTIERI - IMUNOLOGIA BÁSICA E APLICADA
A sua principal atividade biológica é a de ser receptor para antígenos na membrana de linfócitos B. Sabe-se que no processo de maturação, o linfócito B expressa inicialmente a IgM de membrana, posteriormente a IgD e uma outra Ig. Se um antígeno entrar em contato com o linfócito B antes da expressão da IgD, esta célula torna-se tolerante ao antígeno para o qual é específica.
IgE A IgE também é uma imunoglobulina presente nos líquidos internos do corpo em concentrações inferiores a 1% do total das Igs séricas. A concentração média da IgE no soro é de 0,00005 mg/ml, em seres humanos adultos. A vida média da IgE sérica é de 2 dias. O Peso Molecular da IgE é de 188 kDa. A IgE possui cerca de 12% de carboidratos em sua estrutura.
A sua principal atividade é na ADCC, mediada por eosinófilos, frente a vermes helmintos pela ocorrência nestas células de receptores de alta afinidade para a região Fc de IgE.
Outra atividade da IgE é a indução da liberação de histamina, por mastócitos e basófilos, pela associação a receptores para Fc de IgE. Esta função é importante no aumento da permeabilidade vascular durante o processo inflamatório, aumentando a passagem de células e moléculas séricas para os tecidos.
Em estados patológicos, tais como na asma, rinite, choque anafilático a produção de IgE pode aumentar drasticamente, chegando a concentrações próximas a da IgG, causando uma intensa liberação de histamina e choque circulatório.