Baixe Mitocôndria I - CEDERJ e outras Notas de aula em PDF para Biotecnologia, somente na Docsity!
Mitocôndria 26
Ao fi nal desta aula, você deverá ser capaz de:
- Conhecer as principais características das membranas
e compartimentos mitocondriais.
- Discutir a origem simbiótica de mitocôndrias e
cloroplastos.
a
u
l^
a
OBJETIVOS Pré-requisitos
Conhecer as características
de fluidez e permeabilidade
das membranas biológicas.
Saber as características de
eucariotos e procariotos.
Biologia Celular I | Mitocôndria
90 CEDERJ
INTRODUÇÃO Muito antes do surgimento dos seres eucariontes, todas as reações metabólicas que resultam na geração de energia já eram executadas em sistemas proca- riontes ou mesmo pré-bióticos. Nos seres vivos atuais, a energia produzida nesses sistemas é armazenada em moléculas como o ATP e o NADH.H+^ e uti- lizada para geração de calor (nos seres homeotérmicos), movimentos (como o batimento ciliar), na síntese de novas moléculas e outros processos essenciais para a manutenção da vida celular. Duas organelas se destacam na produção de ATP para as células: mitocôndrias e cloroplastos. Estes últimos, você sabe, só existem nos vegetais e serão estudados mais adiante neste mesmo módulo. Nesta aula e na próxima, vamos tratar das mitocôndrias. Na disciplina de Bioquímica, você viu que as mitocôndrias produzem ATP, a principal molécula armazenadora de energia, em dois conjuntos de reações: o ciclo de Krebs e a cadeia respiratória. As mitocôndrias estão entre as organelas mais conhecidas, sendo estudadas há muito tempo. A razão do interesse tão antigo por essa organela pode estar baseada em dois fatores:
- a facilidade com que as mitocôndrias podem ser visualizadas, já que estão acima do limite de resolução do microscópio óptico;
- são muito antigos os registros do consumo de O2 e liberação de CO2 por células vivas, assim como a associação dessa atividade com a produção de ATP.
Mitocôndria, ontem e hoje Mais de 100 anos após sua descoberta, sabemos muito sobre a produção de ATP nas mitocôndrias e conhecemos sua enorme importância para a sobrevivência das células. No entanto, essa interessante organela ainda pode nos surpreender: há menos de 10 anos foi descoberto e vem sendo estudado seu papel central na morte celular programada; ainda há muito por descobrir, como você vai saber em Biologia Celular II.
AS MITOCÔNDRIAS EXISTEM EM TODAS AS CÉLULAS EU-
CARIÓTICAS?
As mitocôndrias estão presentes no citoplasma de quase todas as células eucarióticas, incluindo animais, vegetais, fungos, protozoários e algas. As exceções são protozoários que vivem em ambientes anaeróbicos: amebas, Giardia e Trichomonas.
Biologia Celular I | Mitocôndria
92 CEDERJ
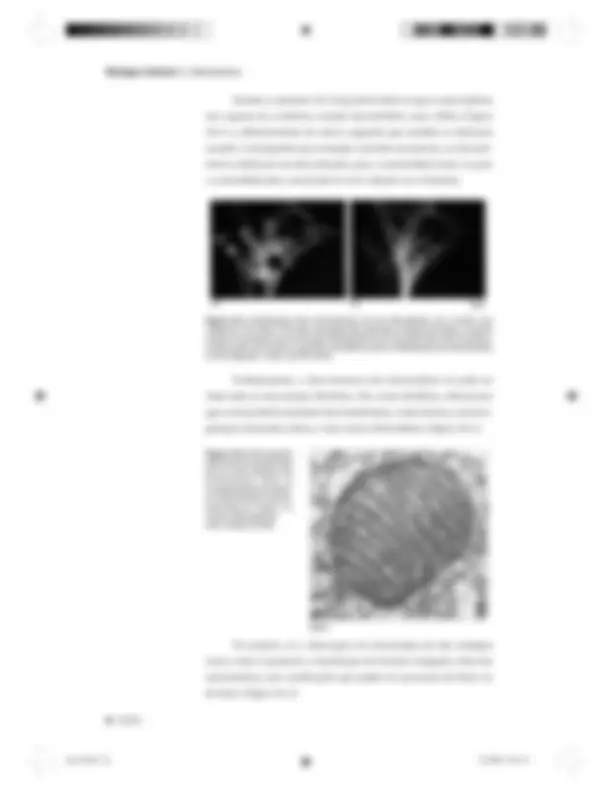
Usando a rodamina 123, foi possível observar que as mitocôndrias são capazes de se deslocar usando microtúbulos como trilhos (Figura 26.1) e, diferentemente de outras organelas que também se deslocam usando o citoesqueleto (por exemplo, vesículas secretórias), as mitocôn- drias se deslocam nas duas direções, para a extremidade minus ou para a extremidade plus , associando-se ora à dineína ora à kinesina.
Figura 26.1: Distribuição das mitocôndrias (a) num fibroblasto vivo corado com rodamina 123. Note o formato alongado das organelas. Depois de fixada, a mesma célula foi incubada com anticorpos antitubulina num procedimento de imunofluo- rescência (b), que revelou a grande coincidência entre a distribuição de mitocôndrias e microtúbulos. Fotos: Lan Bo Chen. Evidentemente, a ultra-estrutura das mitocôndrias só pode ser observada ao microscópio eletrônico. Em cortes ultrafinos, observamos que as mitocôndrias possuem duas membranas, a mais interna, com inva- ginações chamadas cristas, e uma matriz eletrondensa (Figura 26.2).
Figura 26.2: Micrografia eletrônica de transmissão de um corte ultrafino de mitocôndria. Note as invaginações da membra- na mitocondrial interna, formando as cristas, e a matriz eletrondensa. Foto: Daniel Friend.
No entanto, só a observação em microscópio de alta voltagem (veja a Aula 2) permitiu a visualização do formato alongado e fino das mitocôndrias, com ramificações que podem ser processos de fi ssão ou de fusão (Figura 26.3).
CEDERJ 93
AULA
^26
Figura 26.3: alongadas em células de Mitocôndrias MÓDULO 5 epitélio de caramujo obser- vadas ao microscópio de alta voltagem. As mitocôndrias são muito mais eletronden- sas do que o citoplasma e se concentram na região apical da célula. É possível observar cristas e ramificações. Foto de Pierre Favard.
O número de MITOCÔNDRIAS presentes no citoplasma de uma célula eucariótica pode variar de algumas dezenas a milhares, dependendo da necessidade de energia daquele tipo celular naquele momento. Se a demanda por ATP se mantiver alta por algum tempo, a quantidade de mitocôndrias aumentará.
PARA ONDE SE MOVEM AS MITOCÔNDRIAS?
Você pode notar, comparando as Figuras 26.1 e 26.3, que o posi- cionamento das mitocôndrias no citoplasma das células varia muito. Na Figura 26.4, esquematizamos a distribuição de mitocôndrias nos dois tipos celulares das fotos citadas.
Figura 26.4: Em células como os fibroblastos (a) as mitocôndrias se distribuem acompanhando os microtúbulos. Já no epitélio de absorção (b), as mitocôndrias se concentram na região apical, onde ocorre a absorção de nutrientes.
Como assim, a quantidade de mitocôndrias aumenta? Quer dizer que elas proliferam? Sim! Elas se dividem independentemente do ciclo celular, ou seja, de modo não coordenado com a divisão da própria célula. Claro que você lembrou que as mitocôndrias têm DNA! Você também já conhece a hipótese de que as mitocôndrias se originaram de procariotos fagocitados pelo eucarioto ancestral (ou célula pré-eucariótica, reveja a Figura 15.3, na Aula 15) que se tornaram simbiontes. Ao longo desta aula, vamos colecionar semelhanças e diferenças entre mitocôndrias e procariotos. Segure só um pouquinho mais a curiosidade que a gente já vai conversar sobre isso, mais adiante.
A MULTIPLICAÇÃO
DAS MITOCÔNDRIAS
CEDERJ 95
AULA
^26
No coração, as mitocôndrias ficam comprimidas entre as miofibri- MÓDULO 5 las do músculo cardíaco, garantindo que não falte ATP para a contração. O substrato para a produção de ATP também pode estar associado, na forma de gotículas de gordura (Figura 26.6). Você já pensou se para contrair o músculo cardíaco dependesse da chegada de ATP por difusão, a partir de mitocôndrias que estivessem distantes?
Figura 26.6: Esquema (a) e micrografia (b) mostrando mitocôn- drias comprimidas entre as miofibrilas cardíacas, próximas a gotículas de lipídeo.
PRINCIPAIS CARACTERÍSTICAS DAS MEMBRANAS E COM-
PARTIMENTOS MITOCONDRIAIS
As mitocôndrias são delimitadas por duas membranas não coladas, uma externa e uma interna, que assim definem dois compartimentos, o espaço intermembranar e a matriz mitocondrial. As principais caracterís- ticas de cada um desses componentes foram estudadas separadamente, em experimentos de fracionamento celular (Figura 26.7). Em seguida, examinaremos essas características.
(a)
(b)
Biologia Celular I | Mitocôndria
96 CEDERJ
Figura 26.7: Separação dos componentes mito- condriais por fraciona- mento celular.
Membrana mitocondrial externa – é uma membrana cuja bicamada lipídica não possui características especiais, assemelhando- se à bicamada lipídica do retículo endoplasmático. Já as proteínas inseridas nessa membrana são especiais. Podemos citar como típica dessa membrana e essencial para o funcionamento da organela a presença da porina. A porina é uma proteína transmembrana (reveja a Figura 8.8) que tem uma conformação tridimensional tão diferente que forma um poro hidrofílico, permitindo a passagem de moléculas com até 5000 DALTONS pela membrana mitocondrial externa. Ou seja, devido à presença da porina, a membrana mitocondrial externa não forma uma barreira para íons e pequenas moléculas. Curiosamente, a porina da membrana mitocondrial externa é muito similar a proteínas formadoras de poros encontradas na membrana de bactérias Gram negativas.
DALTON
É uma unidade de massa ou raio molecular, que expressa a velocidade com que uma molécula sedimenta por ultracentrifugação. A velocidade de sedimentação varia também com as características do líquido em que ela está dissolvida (densidade, viscosidade), e a velocidade de ultracentrifugação.
Biologia Celular I | Mitocôndria
98 CEDERJ
Membrana mitocondrial interna – essa membrana, sim, é muito especial. É a bicamada lipídica mais fluida e menos permeável de uma célula. Você lembra que as membranas são tanto mais fluidas quanto mais fosfolipídeos com cadeias de ácido graxo curtas e insaturadas e menos colesterol tiverem? A bicamada da membrana mitocondrial interna tem fosfolipídeos com essas características e não possui nenhum colesterol. É importantíssimo que ela seja bastante fluida porque o funcionamento da cadeia transportadora de elétrons depende do choque entre as moléculas. Já a quase impermeabilidade ( quase , porque a membrana mitocondrial interna é permeável à água e a gases, como O 2 , CO 2 e NO) é atribuída a um fosfolipídeo especial: a cardiolipina. A cardiolipina tem esse nome porque foi descrita primeiro em mitocôndrias do músculo cardíaco, mas depois foi encontrada em todas as mitocôndrias. Esse fosfolipídeo é resultado da junção de duas fosfatidilcolinas, formando um fosfolipídeo com quatro cadeias de ácido graxo (é quadrúpede !). Dessa forma, a área ocupada pela cabeça polar corresponde ao dobro do número de cadeias hidrofóbicas na região hidrofóbica da membrana (Figura 26.9).
Figura 26.9: Esquema comparativo dos fosfolipídeos fosfatidilcolina, cardiolipina (verde) e uma bicamada lipídica semelhante à membrana mitocondrial interna.
Quando se faz uma extração bioquímica que separa os lipídeos e as proteínas da membrana mitocondrial interna e depois se seca e pesa o conteúdo, constata-se que essa membrana tem muito mais proteínas do que lipídeos. É a maior relação proteína/lipídeo da célula (70/30). Não é difícil imaginar por que a membrana mitocondrial interna tem tantas proteínas:
- é nela que ocorre a cadeia transportadora de elétrons , e seus componentes são proteínas da membrana interna;
- é na membrana interna que está a enzima que sintetiza ATP, a ATP sintase , em grande número de cópias;
CEDERJ 99
AULA
^26
- se a bicamada lipídica da membrana interna é bastante imper- MÓDULO 5 meável e, no entanto, é preciso que muitas moléculas cheguem à matriz e de lá saiam atravessando essa membrana, ela tem de ter muitos trans- portadores , carreadores etc. Matriz mitocondrial – o conteúdo da matriz mitocondrial é tão concentrado que não se espalha logo que a membrana mitocondrial interna é rompida, como representado na Figura 26.7. A matriz é como um colóide que vai se dispersando devagar. Como está isolada por uma membrana bastante impermeável, a matriz mitocondrial tem composição iônica muito particular. Seu pH, por exemplo, é bem maior que o do citoplasma, cerca de 8, porque a cadeia respiratória bombeia prótons para fora da mitocôndria, alcalinizando a organela (mas sem acidificar o citoplasma, já que o volume deste é muito maior). O estado físico coloidal da matriz é resultado de uma grande concentração de macromoléculas, em que se destacam: a) proteínas solúveis :
- as enzimas do ciclo de Krebs ;
- as enzimas que fazem a β -oxidação dos ácidos graxos;
- a piruvato desidrogenase , complexo enzimático muito grande e abundante que é essencial para o funcionamento do ciclo de Krebs a partir de glicose (e que vamos ver na próxima aula); b) ácidos nucléicos :
- DNA;
- RNAs: ribossomais (rRNA), transportador (tRNA), e o equiva- lente ao mensageiro (mRNA).
Na próxima aula, vamos abordar o metabolismo mitocondrial, com as proteínas e enzimas que dele participam, do ponto de vista celular. Por ora, vamos nos deter nos ácidos nucléicos encontrados na matriz mitocondrial.
O GENOMA DAS MITOCÔNDRIAS E CLOROPLASTOS
A descoberta de que mitocôndrias e cloroplastos (Aulas 28 e 29) possuem ácidos nucléicos (DNA e RNA) despertou enorme curiosidade entre os pesquisadores. Procuramos incluir nesta aula algumas das con- clusões dessas pesquisas, que tiveram enorme influência na elaboração das teorias sobre a evolução da vida em nosso planeta.
CEDERJ 101
AULA
^26
para subunidades de enzimas mitocondriais que dependem da chegada, MÓDULO 5 vindas do citoplasma, de outras subunidades da mesma proteína para poder montar a enzima funcional.
Nem tudo é igual!
Além das muitas semelhanças com os ácidos nucléicos e as enzimas de procariotos, existem diferenças marcantes também. Talvez a maior delas esteja no código genético. No código nuclear de todas as espécies, o significado de alguns CÓDONS é bastante conservado. No genoma mitocondrial de animais e fungos, o código univer- sal para fim de leitura ( stop codon ), por exemplo, não tem esse signi- ficado, codificando para o aminoácido triptofano. Já no genoma das mitocôndrias de plantas, o significado também é parada de leitura. O RNA das mitocôndrias e cloroplastos é mais permissivo no pareamento com os códons. Enquanto nos eucariotos e procariotos cada códon tem um significado, nas organelas a terceira base pode variar, sem mudar o significado. Isso permite usar menos tRNA diferentes (são 22 nas mitocôndrias e 30 no citoplasma). Por isso, os biólogos molecula- res dizem que o código genético das mitocôndrias é mais relaxado , no sentido de menos exigente. O genoma de mitocôndrias e cloroplastos pode ter tamanhos muito diferentes, sendo o dos cloroplastos geralmente maior. Numa mesma planta, o genoma da mitocôndria pode ter cerca de 200 mil pares de bases, enquanto o do cloroplasto tem 250 milhões de pares de bases. Os menores genomas de organelas são os das mitocôndrias de animais, que têm entre 16 e 19 mil pares de bases. O genoma da mito- côndria humana, por exemplo, é bem pequeno e já foi completamente seqüenciado.
CÓDON
Conjunto de três bases nitrogenadas, codifi cando um aminoácido, o início ou o fim da leitura.
Biologia Celular I | Mitocôndria
102 CEDERJ
O genoma mitocondrial humano O seqüenciamento completo do genoma mitocondrial humano mostrou que ele é formado por dois genes que codificam para RNA ribossomal, 22 genes que codificam para RNA transportador e 13 genes que codificam para proteínas. Sua característica mais marcante (comum a vários genomas mitocondriais) é que todos os 16.569 nucleotídeos presentes fazem parte de algum gene, ou seja, não há regiões não codificantes nem espaço para regiões reguladoras. Ao mesmo tempo, apresenta alta taxa de substituição de nucleotídeos, cerca de 10 vezes maior que a taxa nuclear. Por isso, a comparação de seqüências mitocondriais humanas tem sido muito útil no estudo da migração de populações. O seqüenciamento do genoma mitocondrial teve grande impulso depois da guerra do Vietnã porque, pressionado pelas famílias dos soldados mortos que não aceitavam os túmulos do “soldado desconhecido”, o exército americano financiou e executou um grande trabalho de análise do DNA mitocondrial dos soldados mortos. A razão do in- teresse é que o DNA mitocondrial é o mais adequado para a identificação de cadáveres muito destruídos. Por ser menor, está menos sujeito a quebras; por ser circular, é mais fácil saber se a molécula está completa; por estar presente em mais de mil cópias por célula, é mais fácil de ser encontrado; por ser herdado apenas da mãe, as comparações para identificação são mais fáceis. Depois da metodologia estabelecida, o DNA mitocon- drial já foi usado para identificação de cadáveres muito antigos ou destruídos (como os restos mortais da família do último czar russo) e vem sendo freqüentemente usado em genética forense (como, por exemplo, na busca dos netos por avós argentinos, cujos filhos “desapareceram” durante a ditadura).
Não apenas os ácidos nucléicos de mitocôndrias e cloroplastos se parecem com os de procariotos, mas também as próprias enzimas que trabalham na transcrição e na tradução dentro da organela têm caracte- rísticas em comum com as enzimas dos procariotos, sendo inibidas pelas mesmas drogas (Figura 26.11). Surpreendentemente, essas enzimas são elas próprias completamente codificadas por genes do núcleo e tradu- zidas no citoplasma.
Figura 26.11: As drogas que inibem a transcrição no núcleo (α-amanitina) e a tradução e citoplasma (ciclo-heximida) de uma célula são diferentes das que ini- bem transcrição (acridinas) e tradução (cloranfenicol, tetraciclina e eritromicina) em mitocôndrias ou cloroplastos na mesma célula.
Biologia Celular I | Mitocôndria
104 CEDERJ
Figura 26.12: Os complexos de importação mitocondriais t r a n s l o c a s e da membrana externa (TOM) e translocases de membrana interna (TIM).
Vamos imaginar uma proteína de matriz mitocondrial. Ela é sinte- tizada no citoplasma, mantida desenovelada por chaperonas até que sua seqüência de sinal seja reconhecida por uma TOM (Figura 26.13).
Figura 26.13: Importação de uma proteína da matriz mitocondrial. Uma vez reconhecidas, as chaperonas se soltam e a hidrólise de ATP por essas chaperonas contribui para que a TOM transporte a proteína para dentro. Logo que sua seqüência-sinal aparecer no espaço intermembranar, uma TIM vai reconhecê-la e translocá-la para a matriz com a ajuda da energia do gradiente de prótons da cadeia respiratória (Figura 26.13). Ao chegar à matriz, sua seqüência-sinal é cortada, e a conformação funcional é adquirida com a ajuda das chaperonas mito- condriais. Com certeza, esse mecanismo seria mais eficiente se a passagem pelas duas membranas fosse consecutiva. Essa idéia é confirmada pela existência de sítios de contato entre a duas membranas da mitocôndria, observada em algumas preparações especiais de microscopia eletrônica (Figura 26.14), e pela descoberta de que uma das TIM (a TIM 23) é uma proteína das duas membranas.
CEDERJ 105
AULA
^26
MÓDULO 5
Figura 26.14: Observando ao microscópio eletrônico de alta voltagem um corte espesso de mitocôndrias do fungo Neurospora crassa preservadas por congelamento, é possível observar que as membranas externa e interna mantêm distância aproxi- madamente constante, exceto nos pontos de contato (no detalhe), que se acredita sejam os locais de importação de proteínas para a matriz mitocondrial. Foto de Daniela Nicastro (J. Struct. Biol. 129:48, 2000).
ORIGEM SIMBIÓTICA DE MITOCÔNDRIAS E CLOROPLASTOS
Agora que você já conhece melhor as características de mitocôn- drias e cloroplastos, podemos voltar à discussão sobre a origem dessas organelas. As mitocôndrias teriam se originado a partir da fagocitose de procariotos aeróbicos por eucariotos que não os destruíram e passaram a usufruir de grandes vantagens numa época em que o teor de oxigê- nio da atmosfera terrestre estava aumentando. Os novos eucariotos desenvolveram, então, uma relação de simbiose com os procariotos que fagocitaram e passaram a predominar porque podiam obter muito mais energia a partir dos substratos disponíveis. A aquisição das mitocôndrias também liberou a membrana plasmática das tarefas de produção de energia, possibilitando outras especializações que contribuíram muito para o aumento da complexidade dos eucariotos. Mas será que essa hipótese é verdadeira? Ainda não foi possível testá-la diretamente, já que ainda não se conseguiu “cultivar” mitocôn- drias e depois oferecê-las a um eucarioto sem mitocôndrias, como uma ameba, na tentativa de refazer o evento simbiótico. O estudo de outras
CEDERJ 107
AULA
^26
De que procarioto as mitocôndrias e os cloroplastos descedem? MÓDULO 5
Certamente, a relação simbiótica entre eucariotos e mitocôndrias é mais antiga e data de um período anterior à separação entre animais e plantas. Acredita-se que o evento endocítico que incorporou as mitocôn- drias aos eucariotos data de cerca de 1,5 x 10^9 anos, quando os níveis de oxigênio na atmosfera aumentaram. Comparando as seqüências gênicas, as mitocôndrias parecem ter se originado de bactérias púrpura; e os cloroplastos, muitos anos depois, com animais e vegetais já separados evolutivamente, de bactérias fotossintéticas (Figura 26.15).
Figura 26.15: A árvore filogenética da provável evolução de mitocôndrias e cloroplastos e seus ances- trais procariotos.
Biologia Celular I | Mitocôndria
108 CEDERJ
EXERCÍCIOS
- As mitocôndrias de todos os tipos celulares são morfologicamente iguais?
- Como se organiza estruturalmente a mitocôndria?
- Quais as principais características da membrana mitocondrial externa?
- Quais as principais características da membrana mitocondrial interna?
- Quais as principais características do espaço intermembranas?
- Quais as principais características da matriz mitocondrial?
- Por que as mitocôndrias representaram um grande salto evolutivo para os seres euca- riontes?
- Como se distribuem as mitocôndrias em uma célula?
- Como é feita a importação de proteínas mitocondriais cujos genes se encontram no núcleo?
- Por que se acredita que as mitocôndrias resultam de uma relação simbiótica entre uma bactéria e uma célula eucarionte primitiva que a fagocitou?