


















Estude fácil! Tem muito documento disponível na Docsity

Ganhe pontos ajudando outros esrudantes ou compre um plano Premium

Prepare-se para as provas
Estude fácil! Tem muito documento disponível na Docsity
Prepare-se para as provas com trabalhos de outros alunos como você, aqui na Docsity
Encontra documentos específicos para os exames da tua universidade
Prepare-se com as videoaulas e exercícios resolvidos criados a partir da grade da sua Universidade
Responda perguntas de provas passadas e avalie sua preparação.
Ganhe pontos para baixar
Ganhe pontos ajudando outros esrudantes ou compre um plano Premium
Este documento aborda detalhadamente a fisiologia da glândula tireoide, incluindo a formação e a regulação dos hormônios tireoidianos, bem como o metabolismo do cálcio e sua regulação pelos hormônios paratireoidianos e calcitonina. O texto descreve o processo de captação e oxidação do iodo pelas células da tireoide, a síntese dos hormônios t3 e t4, e seus efeitos no metabolismo de carboidratos, proteínas e no sistema nervoso. Além disso, o documento explora a homeostase do cálcio, a interação entre o paratormônio (pth), a vitamina d e a calcitonina, bem como os mecanismos de regulação da concentração de cálcio iônico no líquido extracelular. Com uma abordagem detalhada e abrangente, este texto é uma fonte valiosa de informações para estudantes de áreas como medicina, farmácia, biologia e fisiologia, fornecendo uma compreensão profunda dos processos fisiológicos envolvidos na regulação da tireoide e do metabolismo do cálcio.
Tipologia: Exercícios
1 / 25

Esta página não é visível na pré-visualização
Não perca as partes importantes!
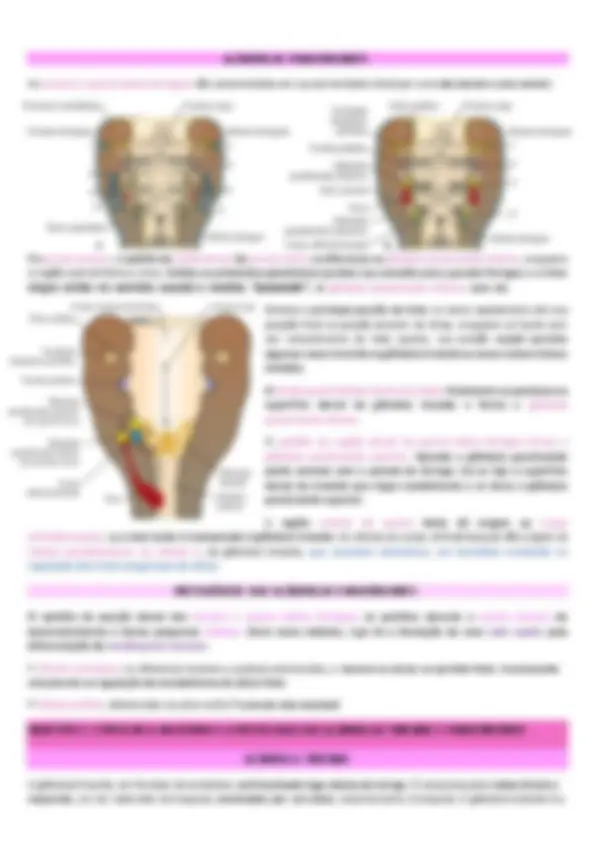
- Contemplar o ducto tireoglosso GLÂNDULA TIREOIDE A glândula tireoide aparece como proliferação epitelial no assoalho da faringe entre o tubérculo ímpar e a cópula, no ponto indicado mais tarde pelo forame cego (abertura proximal do ducto tireoglosso que persiste como uma pequena fosseta no dorso da língua). Subsequentemente, a tireoide desce na frente do intestino faríngeo como um divertículo bilobado. Durante essa migração, a tireoide permanece conectada à língua por um canal estreito, o ducto tireoglosso. Mais tarde, esse ducto desaparece, geralmente, até a sétima semana. A princípio o primórdio da tireóide é oco, mas rapidamente se torna uma massa sólida de células que se divide em lobos direito e esquerdo. Com a continuação do desenvolvimento, a glândula tireoide desce para a frente do osso hioide e das cartilagens laríngeas. Ela alcança sua posição final na frente da traqueia na sétima semana. Nesse período, ela adquiriu um pequeno istmo mediano e dois lobos laterais. A tireoide começa a funcionar por volta do fim do terceiro mês, quando se tornam visíveis os primeiros folículos contendo coloide. As células foliculares produzem o coloide, que funciona como fonte de tiroxina (T4) e de tri-iodotironina (T3). HISTOGÊNESE DOS FOLÍCULOS TIREOIDIANOS O folículo tireoidiano constitui a unidade estrutural e funcional da glândula tireoide. Podemos defini-lo como um compartimento aproximadamente esférico, semelhante a um cisto, com uma parede formada por epitélio simples
As terceira e quarta bolsas faríngeas são caracterizadas em sua extremidade distal por uma aba dorsal e uma ventral. Na quinta semana, o epitélio da região dorsal da terceira bolsa se diferencia na glândula paratireoide inferior, enquanto a região ventral forma o timo. Ambos os primórdios glandulares perdem sua conexão com a parede faríngea, e o timo migra então no sentido caudal e medial, “puxando”, a glândula paratireoide inferior com ele. Embora a principal porção do timo se mova rapidamente até sua posição final na porção anterior do tórax, enquanto se funde com seu complemento do lado oposto, sua porção caudal persiste algumas vezes inserida na glândula tireoide ou como nichos tímicos isolados. O tecido paratireóideo da terceira bolsa finalmente se posiciona na superfície dorsal da glândula tireoide e forma a glândula paratireoide inferior. O epitélio da região dorsal da quarta bolsa faríngea forma a glândula paratireoide superior. Quando a glândula paratireoide perde contato com a parede da faringe, ela se liga à superfície dorsal da tireoide que migra caudalmente e se torna a glândula paratireoide superior. A região ventral da quarta bolsa dá origem ao corpo ultimobranquial, que mais tarde é incorporado à glândula tireoide. As células do corpo ultimobranquial dão origem às células parafoliculares, ou células C, da glândula tireoide, que secretam calcitonina, um hormônio envolvido na regulação dos níveis sanguíneos de cálcio. HISTOGÊNESE DAS GLÂNDULAS PARATIREOIDES O epitélio da porção dorsal das terceira e quarta bolsas faríngeas se prolifera durante a quinta semana de desenvolvimento e forma pequenos nódulos. Entre esses nódulos, logo há a formação de uma rede capilar pela diferenciação do mesênquima vascular.
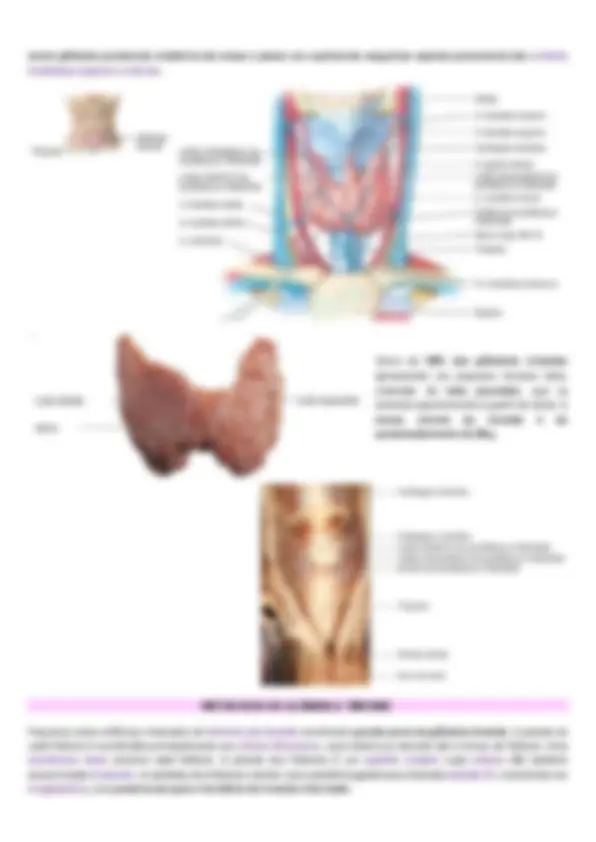
maior glândula puramente endócrina do corpo e possui um suprimento sanguíneo copioso proveniente das artérias tireóideas superior e inferior. Cerca de 50% das glândulas tireoides apresentam um pequeno terceiro lobo, chamado de lobo piramidal, que se estende superiormente a partir do istmo. A massa normal da tireoide é de aproximadamente de 30 g. HISTOLOGIA DA GLÂNDULA TIREOIDE Pequenos sacos esféricos chamados de folículos da tireoide constituem grande parte da glândula tireoide. A parede de cada folículo é constituída principalmente por células foliculares, cuja maioria se estende até o lúmen do folículo. Uma membrana basal envolve cada folículo. A parede dos folículos é um epitélio simples cujas células são também denominadas tireócitos. A cavidade dos folículos contém uma substância gelatinosa chamada coloide (F), consistindo em tiroglobulina, uma proteína da qual o hormônio da tireoide é derivado.
Parcialmente incrustadas na face posterior dos lobos direito e esquerdo da glândula tireoide, encontramos várias pequenas massas de tecido arredondadas chamadas de glândulas paratireoides. Cada uma pesa cerca de 40 mg (0, g). Em geral, uma glândula paratireoide inferior e uma superior estão fixadas em cada lobo da tireoide, em um total de Tais glândulas contêm dois tipos de células epiteliais. As células mais numerosas, chamadas de células principais, produzem o paratormônio (PTH), têm forma poligonal, núcleo vesicular e citoplasma fracamente acidófilo. As células oxifílicas aparecem aos 7 anos de idade e a partir daí aumentam progressivamente de número. São poligonais, maiores e mais claras que as células principais. A função dessas células é desconhecida. Além disso, no câncer de glândulas paratireoides, as células oxifílicas secretam PTH em excesso. quatro.
- Contemplar o metabolismo do cálcio. FISIOLOGIA DA GLÂNDULA TIREOIDE Essa glândula secreta dois hormônios principais, a tiroxina e a tri-iodotironina, usualmente chamados T4 e T3, respectivamente. Ambos aumentam intensamente o metabolismo do organismo. A ausência completa de secreção tireoidiana, em geral, faz com que o metabolismo basal caia para 40% a 50% do normal, e o excesso extremo de secreção pode aumentá-lo de 60% a 100%. A secreção tireoidiana é controlada, principalmente, pelo hormônio tireoestimulante (TSH), secretado pela hipófise anterior. A tireoide também secreta calcitonina, hormônio importante para o metabolismo do cálcio. SÍNTESE E SECREÇÃO DOS HORMÔNIOS METABÓLICOS TIREOIDIANOS Cerca de 93% dos hormônios metabolicamente ativos, secretados pela tireoide, consistem em tiroxina, e 7% são tri- iodotironina. Entretanto, quase toda a tiroxina é, por fim, convertida em tri-iodotironina nos tecidos, de modo que ambas são funcionalmente importantes. As funções desses dois hormônios são qualitativamente iguais, mas diferem na velocidade e na intensidade de ação. A tri-iodotironina é cerca de quatro vezes mais potente que a tiroxina, mas está presente no sangue em menor quantidade e persiste por um tempo muito curto. O IODO É NECESSÁRIO PARA A FORMAÇÃO DE TIROXINA Para formar quantidades normais de tiroxina, é necessária a ingestão de cerca de 50 miligramas de iodo na forma de iodeto a cada ano, ou cerca de 1 mg/semana. Para prevenir a deficiência de iodo, o sal comum de cozinha é suplementado com cerca de uma parte de iodeto de sódio para cada 100.000 partes de cloreto de sódio. DESTINO DOS IODETOS INGERIDOS . Os iodetos ingeridos por via oral são absorvidos pelo trato gastrointestinal para o sangue aproximadamente do mesmo modo que o cloreto. Nas condições normais, a maior parte do iodeto é rapidamente excretada pelos rins, mas por volta de um quinto é seletivamente removido do sangue circulante pelas células da tireoide e usado para a síntese dos hormônios tireoidianos.
A peroxidase localiza-se na membrana apical da célula ou ligada a ela, produzindo, assim, o iodo oxidado, exatamente no ponto da célula em que a molécula de tireoglobulina surge, vinda do aparelho de Golgi e através da membrana celular, sendo armazenada no coloide da tireoide. Quando o sistema da peroxidase é bloqueado ou quando está hereditariamente ausente das células, a formação de hormônios tireoidianos cai para zero. A ligação do iodo com a molécula de tireoglobulina é chamada organificação da tireoglobulina. O iodo oxidado, até mesmo na forma molecular, liga-se diretamente, embora de forma muito lenta, ao aminoácido tirosina. Nas células da tireoide, entretanto, o iodo oxidado está associado à enzima peroxidase tireoidiana, que reduz a duração desse processo para segundos ou minutos. Por isso, no intervalo de tempo em que a molécula de tireoglobulina leva para ser liberada pelo aparelho de Golgi, ou para ser secretada pela membrana apical da célula para o folículo, o iodo se liga a cerca de um sexto de seus aminoácidos tirosina.
coloide, formando vesículas pinocíticas que penetram pelo ápice da célula. Então, lisossomos no citoplasma celular imediatamente se fundem com as vesículas para formar vesículas digestivas que contêm as enzimas digestivas dos lisossomos misturadas com o coloide. Múltiplas proteases entre as enzimas digerem as moléculas de tireoglobulina e liberam tiroxina e tri- iodotironina, em sua forma livre, que se difundem pela base da célula tireoidiana para os capilares adjacentes. Assim, os hormônios tireoidianos são liberados no sangue. Parte da tireoglobulina do coloide entra na célula tireoidiana por endocitose, depois de se ligar à megalina, uma proteína localizada na membrana luminal das células. Em seguida, o complexo megalina- tireoglobulina é transportado através da célula por transcitose até a membrana basolateral, onde uma parte da megalina permanece unida à tireoglobulina e é liberada no sangue capilar. Cerca de três quartos da tirosina iodada na tireoglobulina nunca se torna hormônio, permanecendo como monoiodotirosina e di-iodotirosina. Durante a digestão da molécula de tireoglobulina para provocar a liberação de tiroxina e tri-iodotironina, essas tirosinas iodadas também são liberadas das moléculas de tireoglobulina. Entretanto, não são secretadas para o sangue. Ao contrário, seu iodo é clivado pela enzima deiodinase, que disponibiliza praticamente todo o iodo para a reciclagem na glândula e a formação de novas moléculas de hormônios tireoidianos. Na ausência congênita da deiodinase, muitas pessoas podem apresentar deficiência de iodo devido à falha desse processo de reciclagem. Estão Ligadas a Proteínas Plasmáticas. Ao serem liberadas no sangue, mais de 99% da tiroxina e triiodotironina se combinam imediatamente às diversas proteínas plasmáticas sintetizadas pelo fígado. Elas se combinam, principalmente, com a globulina de ligação de tiroxina e muito menos com a pré-albumina de ligação de tiroxina e albumina. A Tiroxina e a Tri-iodotironina são liberadas lentamente para as Células Teciduais, devido à alta afinidade das proteínas plasmáticas de ligação dos hormônios tireoidianos, essas substâncias — em particular, a tiroxina — são liberadas de forma lenta para as células teciduais. A metade da tiroxina sanguínea é liberada, aproximadamente, a cada seis dias, enquanto a metade da tri-iodotironina — devido à sua menor afinidade com as proteínas transportadoras — é liberada para as células em cerca de um dia. Ao penetrar as células, a tiroxina e a tri-iodotironina se ligam, novamente, a proteínas intracelulares; a ligação da tiroxina é mais forte que a da triiodotironina. Portanto, são de novo armazenadas, mas dessa vez nas próprias células-alvo, e são usadas, lentamente, ao longo de dias ou semanas. A ação dos hormônios tireoidianos tem início lento e longa duração. Após a injeção de grande quantidade de tiroxina no ser humano, praticamente não se detectam efeitos no metabolismo por 2 a 3 dias, o que demonstra seu longo período de latência, antes do início da atividade da tiroxina. Uma vez que a atividade se inicia, ela aumenta progressivamente, até atingir um máximo em 10 a 12 dias. Depois disso, declina com uma meia- vida de cerca de 15 dias. Parte da atividade persiste por 6 semanas a 2 meses. As ações da tri-iodotironina são cerca de quatro vezes mais rápidas que as da tiroxina, com período de latência de apenas 6 a 12 horas e atividade celular máxima ocorrendo de 2 a 3 dias.
O hormônio tireoidiano provoca efeitos gerais e específicos no crescimento. Um importante efeito do hormônio tireoidiano é promover o crescimento e desenvolvimento do cérebro durante a vida fetal e nos primeiros anos de vida pós- natal. Se o feto não secretar quantidade suficiente de hormônio tireoidiano, o crescimento e a maturação do cérebro, antes e após o nascimento, são muito retardados, e o cérebro permanece menor que o normal. Sem o tratamento específico dentro de dias ou semanas após o nascimento, a criança que não tem a glândula tireoide permanece mentalmente deficiente para o resto da vida.
excesso ou a redução da produção de hormônio tireoidiano.
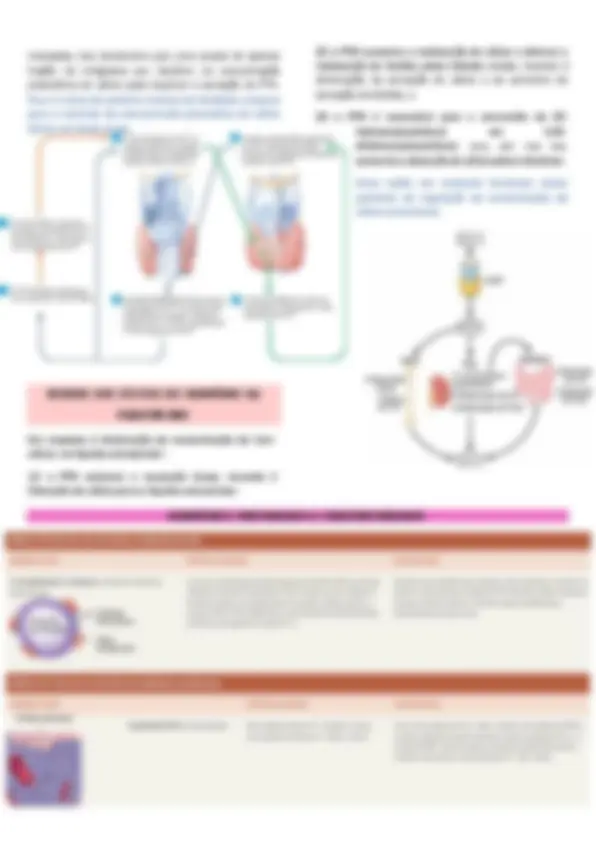
(1) cerca de 41% (1 mmol/L) do cálcio encontram-se combinado às proteínas plasmáticas e, nessa forma, não é difusível através da membrana dos capilares; (2) aproximadamente 9% do cálcio (0,2 mmol/L) são difusíveis através da membrana dos capilares, mas está combinado às substâncias aniônicas do plasma e líquidos intersticiais (p. ex., citrato e fosfato) e, desse modo, não é ionizado; e (3) os 50% restantes do cálcio no plasma apresentam-se como difusíveis através da membrana dos capilares e ionizados. Dessa forma, os líquidos plasmáticos e intersticiais mostram concentração normal do cálcio iônico de aproximadamente 1,2 mmol/L (ou 2,4 mEq/L, por ser íon divalente), nível correspondente apenas à metade da concentração plasmática total desse elemento. Esse cálcio iônico é a forma relevante para a maior parte das funções do cálcio no corpo, incluindo seu efeito sobre o coração, o sistema nervoso e a formação óssea. Se forem injetados sais solúveis de cálcio por via intravenosa, a concentração do cálcio iônico poderá aumentar imediatamente para níveis elevados. Entretanto, essa concentração de cálcio iônico retorna ao normal dentro de 30 a 60 minutos. Do mesmo modo, o contrário. Esses efeitos se devem, em grande parte, à presença de cálcio do tipo intercambiável na composição óssea, que sempre está em equilíbrio com os íons cálcio nos líquidos extracelulares. Pequena porção desse cálcio intercambiável também corresponde ao cálcio encontrado em todas as células, particularmente nos tipos celulares com alta permeabilidade, como as células do fígado e do trato gastrointestinal. No entanto, a maior parte do cálcio intercambiável está no osso, representando, normalmente, um valor em torno de 0,4% a 1% do cálcio ósseo total. Esse cálcio é depositado nos ossos em forma de sal prontamente mobilizável, como o CaHPO4 e outros sais cálcicos amorfos. A importância do cálcio intercambiável está na provisão de um mecanismo rápido de tamponamento para manter a concentração de cálcio iônico nos líquidos extracelulares, evitando sua ascensão a níveis excessivos, ou sua queda a níveis baixos, em condições transitórias de alta ou baixa disponibilidade de cálcio O 1,25-di-hidroxicolecalciferol atua como um tipo de “hormônio” para promover a absorção intestinal de cálcio. Favorece essa absorção, em grande parte, por meio do aumento, em torno de dois dias, da formação de calbindina, uma proteína ligante do cálcio, nas células epiteliais intestinais. Essa proteína atua na borda em escova dessas células para transportar o cálcio para o citoplasma celular. O cálcio, então, desloca-se através da membrana basolateral da célula por difusão facilitada. A absorção de cálcio é diretamente proporcional à quantidade dessa proteína ligante. Além disso, essa proteína permanece nas células por algumas semanas, após a remoção do 1,25-di-hidroxicolecalciferol do corpo, causando um efeito prolongado de absorção do cálcio. Outros efeitos do 1,25-di-hidroxicolecalciferol que podem ter participação no estímulo da absorção de cálcio incluem a formação de (1) adenosina trifosfatase estimulada pelo cálcio na borda em escova das células epiteliais; e (2) fosfatase alcalina nas células epiteliais. Os detalhes precisos de todos esses efeitos não estão esclarecidos.
A calcitonina, hormônio peptídico secretado pela glândula tireoide, tende a diminuir a concentração plasmática do cálcio e, em geral, tem efeitos opostos aos do PTH. No entanto, o papel quantitativo da calcitonina nos seres humanos é bem menor que o do PTH na regulação da concentração do cálcio iônico. A síntese e a secreção da calcitonina ocorrem nas células parafoliculares, ou células C, situadas no líquido intersticial entre os folículos da glândula tireoide. Essas células constituem apenas cerca de 0,1% da glândula tireoide humana e representam os resquícios das glândulas ultimobranquiais de peixes, anfíbios, répteis e aves. A calcitonina é um peptídeo com 32 aminoácidos e peso molecular de aproximadamente 3.400. O Aumento da Concentração Plasmática do Cálcio Estimula a Secreção de Calcitonina. O principal estímulo para a secreção de calcitonina é a elevação da concentração de cálcio iônico no líquido extracelular. Em contraste, a secreção do PTH é estimulada pela queda na concentração de cálcio. Em animais jovens, mas muito menos em animais com mais idade e em seres humanos, o aumento da concentração plasmática do cálcio em torno de 10% provoca elevação imediata de duas vezes ou mais na secreção de calcitonina. Isso gera um segundo mecanismo de feedback hormonal para o controle da concentração plasmática do cálcio iônico; no entanto, esse mecanismo é relativamente fraco e atua de modo oposto ao sistema representado pelo PTH.
a difusão dos íons cálcio até as membranas celulares do líquido ósseo. Então, a bomba de cálcio, presente do outro lado da membrana celular, transfere os íons cálcio para o líquido extracelular no restante do percurso. ➢ Fase Lenta da Reabsorção Óssea e Liberação do Fosfato de Cálcio — Ativação dos Osteoclastos. Um efeito muito mais conhecido e evidente do PTH consiste na ativação dos osteoclastos. No entanto, essas células não têm receptores proteicos em suas membranas para o PTH. Em vez disso, acredita-se que os osteoblastos e os osteócitos ativados emitam “sinais” secundários para os osteoclastos. Um sinal secundário importante é o RANKL, que ativa receptores nas células préosteoclastas e as transforma em osteoclastos maduros, que começam sua tarefa habitual de engolfamento do osso em semanas ou meses. A ativação do sistema osteoclástico ocorre em dois estágios: (1) ativação imediata dos osteoclastos já formados; e (2) formação de novos osteoclastos. Após alguns meses de níveis excessivos de PTH, a ressorção osteoclástica pode levar ao enfraquecimento ósseo e à estimulação secundária dos osteoblastos, na tentativa de corrigir a condição enfraquecida do osso. Portanto, o efeito tardio consiste, efetivamente, na intensificação das atividades osteoblástica e osteoclástica.
tracejada; isso demonstra que uma queda de apenas fração de miligrama por decilitro na concentração plasmática do cálcio pode duplicar a secreção de PTH. Essa é a base do potente sistema de feedback corporal para o controle da concentração plasmática do cálcio iônico em longo prazo. (2) o PTH aumenta a reabsorção de cálcio e diminui a reabsorção de fosfato pelos túbulos renais, levando à diminuição da excreção de cálcio e ao aumento da excreção de fosfato; e (3) o PTH é necessário para a conversão de 25 - hidroxicolecalciferol em 1,25- dihidroxicolecalciferol, que, por sua vez, aumenta a absorção de cálcio pelos intestinos. Essas ações em conjunto fornecem meios potentes de regulação da concentração do cálcio extracelular. Em resposta à diminuição da concentração de íons cálcio, no líquido extracelular: (1) o PTH estimula a ressorção óssea, levando à liberação de cálcio para o líquido extracelular; HORMÔNIOS TIREOIDIANOS E PARATIREOIDIANOS