¡Descarga biologia bloque 4 resumen y más Apuntes en PDF de Biología solo en Docsity!
Cuarto Parcial de Biología
Tipos de Tejidos
Tejido epitelial: recubrimiento, está fijo a una lámina basal. Tejido conectivo o conjuntivo: es el más abundante, conecta y protege a otros tejidos. Es el principal productor de la matriz extracelular. Tejido muscular Tejido nervioso Tejido linfoide Tejido sanguineo
Matriz Extracelular
Los tejidos además de células también están compuestos por sustancias y elementos intercelulares que en conjunto se denominan matriz extracelular. La matriz extracelular es una red de materiales extracelulares, una mezcla amorfa de proteinas y polisacáridos. Esta rellena espacios entre células, une células entre sí y tejidos. Funciones de la matriz extracelular Rellenar el espacio existente entre las células otorgando resistencia a la compresión y al estiramiento (estas propiedades decaen con el envejecimiento). Medio por donde llega los nutrientes y se eliminan los desechos celulares (función que se encuentra alterada en la celulitis). Proveer a la célula “puntos fijos” donde aferrarse. Espacio por donde migran las células cuando se desplazan de un punto a otro del organismo. Medio por el cual arriban a las células las señales bioquímicas (por ejemplo hormonas y citoquinas). Depende del tejido de hidratación, dureza (hueso y dientes). Células formadoras: células del tejido conectivo (fibroblastos). Conformación de la Matriz extracelular La matriz extracelular tiene dos componentes principales que son los proteínas estructurales fibrosas y un gel formado por carbohidratos. Las proteínas estructurales fibrosas son el componente principal de los tejidos conjuntivos, estas proteínas son: colágeno, elastina (proteinas estrcutructurales) fibronectina y laminita (proteínas adhesivas). El gel está formado a partir de polisacáridos denominados Glucosaminoglicanos o GAGs.
Proteínas estructurales Fibrosas
Entre las proteínas que conforman la matriz extracelular están: Colágeno El colágeno es la glucoproteína estructural más abundante (1/4 de la proteína total) y esta es sintetizada por los fibroblastos. Es un componente esencial de los tendones y de la sustancia extracelular que rodea las células óseas en el hueso, hay 15 tipos distintos de colágeno y según la combinación puede haber 25 cadenas distintas. El colágeno en general se encuentra en piel, tendones, cartílagos, vasos sanguíneos, dientes, membranas basales, cornea, cuerpo vítreo, tejidos de sostén, etc. Son filamentos insolubles que soportan grandes cargas, más de 10 enfermedades son ocasionadas por mutaciones en uno de los genes que codifican el colágeno (12 cromosomas distintos). Estructura de la Colágena Posee una secuencia de AA simple y periódica. Cada 3 AA hay una glicina. Su estructura general es: Gly – X – Y Disposición de trenza X = prolina o hidroxiprolina Y = lisina o hidroxilisina Las cadenas de colágeno se agrupan en triples hélices (tres cadenas forman una triple hélice) El colágeno Tipo I se encuentra en la dermis (el 70% de la piel es colágeno), los tendones, el hueso y las arterias.. Esta formado por una triple hélice (2 cadenas alfa I y 1 cadena alfa II) El las láminas basales se encuentra el colágeno Tipo IV.
Glucosaminoglicanos y proteoglicanos
Los glicosaminoglicanos son polisacáridos (osea, carbohidratos o azúcares) complejos. Entre los más abundantes esta el ácido hialurónico (es el de mayor tamaño y no se encuentra sulfatado), el condroitín sulfato, el dermatán sulfato, el heparán sulfato y el queratán sulfato. Cuando estos se combinan con proteinas pasan a llamarse proteoglicanos. Los glucosaminoglicanos también son llamados mucopolisacáridos. Los glicosaminoglicanos son moléculas muy ácidas, con numerosas cargas negativas que atraen grandes cantidades de ion sodio (Na) y, por lo tanto, de agua, lo cual aumenta la turgencia de la matriz extracelular. Son los responsables de que la dermis posea una masa gelatinosa con gran capacidad de hidratación. Tienen gran influencia en la turgencia y tirantez de la piel. Los proteoglicanos tienen la función de “hidratación” (gel), dan resistencia a tensión y comprensión, este se une a la colágena, receptores de superficie celular, estos adhieren las células a la matriz extracelular. Adherencia Celular La adhesión intercelular y la de células con componentes de la matriz extracelular son fenómenos que tienen un papel clave en la organización general de los seres vivos multicelulares. La integridad y organización general de los diversos tejidos y órganos de un individuo dependen de la adecuada interacción entre los elementos que los componen.
Proteínas o moléculas de adhesión celular (CAM)
Existen interacciones transitorias célula-célula y célula-matriz. Entre receptores y ligandos, por ejemplo: Interacciones de los leucocitos y las plaquetas con el endotelio vascular en la inflamación. En la reacción inmunitaria. Entre las células y la matriz (locomoción celular, en la formación de vasos en la respiración) Pero también existen interacciones estables célula-célula y célula- matriz. Tanto las interacciones estables como las transitorias se dan gracias a las CAM. Todas las CAM son proteínas integrales de membrana. Las CAM se dividen en: La familia de las selectinas La familia de las integrinas (adhesión focal / hemidesmosoma)
Las superfamilias de las Inmunoglobulinas (lg) Las cadeherinas (Unión adherente / Desmosoma) La familia de las Selectinas Participan en la extravasación de los leucocitos hacia los tejidos. Los ligandos son los carbohidratos de las membranas. La unión es dependiente de Ca+2^ y Mg+2. Estas crean uniones transitorias en los vasos sanguineos. La familia de las Integrinas Median interacciones débiles entre las células y la matriz, dependiente de Ca+2^ y Mg+2. La familia de las integrinas comprende a un grupo amplio de moléculas heterodiméricas constituidas por dos subunidades polipeptídicas transmembranales denominadas alfa y beta. La integrina está compuesta por cadenas polipetídicas α y β. La superfamilia de las Inmunoglobulinas Participan en la adhesión homofila (una molécula de adhesión de la superficie se une a la misma molécula de superficie de otra célula) es
Uniones estrechas o impermeables La unión estrecha u oclusiva se encuentra separando los líquidos extracelulares que bañan las regiones apicales y basales de las células (con el objeto de que cumplan sus respectivas funciones) y forman barreras que tornan impermeables determinadas cavidades (como la luz del intestino, la vejiga, etc). Esta relación se da entre proteinas transmembrana de las dos células involucradas, las proteínas de este tipo de unión son la ocludina y la claudina al fusionarse, estas forman uniones extremadamente fuertes y prácticamente fusionan dos células estableciendo una unión impermeable. Uniones comunicantes También son conocidas como uniones “GAP”. Aquí las membranas de dos células poseen proteínas que conforman semicanales transmembrana, que las interconectan y permiten el paso de moléculas entre ambas. La unión esta constituida por un anillo de seis subunidades proteicas llamadas conexinas que juntas forman poros o conexones. Tienen la capacidad de abrirse o cerrarse, se cree que es estímulo de fosforilación de la conexina. Estas uniones permiten el paso de iones y moléculas hidrosolubles orgánicas de pequeño tamaño. Es
células vegetales las uniones comunicantes son denominadas plasmodesmos, estos atraviesan la pared celular formando el conducto por la membrana citoplásmica, el conducto es denominado desmotúbulo
. Los plasmodesmos son sintetizados en el RE liso. Uniones Basales (célula-Matriz) Intervienen integrinas que se unen a proteínas de la matriz extracelular. Las hay de dos tipos: Contactos focales: aquí las integrinas están ancladas a filamentos de actina. Hemidesmosomas: actúa la integrina α 6 β 4 unida a filamentos intermedios.
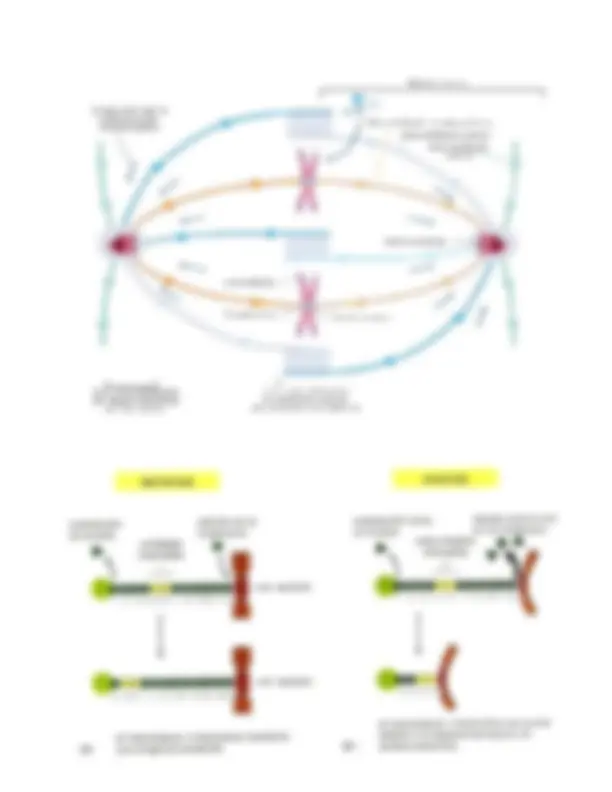
- Esta sería la primera unión célula- matríz (union basal), este es un hemidesmosoma el cuál está formado por integrina α 6 β 4 (cositas verdes) una plectina (base negra) y filamentos intermedios (cositas fusia). Entonces hemidesmosoma = integrina α 6 β 4 + plectina + filamentos intermedios.
- Esta sería la segunda y última unión célula matríz, esta es una unión de contacto focal que esta formada por integrina (cositas verdes) y filamentos de actina (cositas verdes). Contacto focal = integrina + filamentos de actina. El número 1,2,3, y 4 son uniones célula-célula. Y los números 5 y 6 son uniones basales o uniones célula matriz. Solo falta recordar que las uniones de adhesión célula – célula pueden ser interacciones homofílicas) y pueden ser interacciones heterofílicas: Homofílicas: Cadherina-Cadherina Ig – Ig (recordemos que Ig = inmonuglobulina) Heterofílicas: Selectina – Glicoproteína Integrina – Ig Bueno creo que eso es todo, espero ayude en algo.
Comunicación Intercelular Ok, empezemos. La célula responde a estímulos, este estímulo es llevado a la célula blanco (osea la célula que recibe el mensaje) por un mensajero. Este mensajero generalmente es específico, y un mismo mensajero puede estimular diferentes células. El mensaje se encuentra en los ligandos (también llamados moléculas señal, primeros mensajeros, etc) estos ligandos pueden ser proteínas, pequeños péptidos, aminoácidos, nucleótidos, esteroides, retinoides, derivados de ácidos grasos e incluso gases disueltos como el óxido nítrico y el monóxido de carbono. En resumen todos estos pueden ser ligandos (o mensajes), estos ligandos son liberados por las denominadas células señal (osea, la célula que emite el mensaje), este mensaje llega a la célula receptora (también conocida como célula blanco, diana u objetivo).
Secuencia de eventos en la comunicación entre células
Se sintetiza la señal química por la célula secreto (emisora) Se exocita la señal (se libera la señal) Se transporta la señal hacia la célula blanco o diana (célula receptora) Re recibe el mensaje por medio de proteínas receptoras (específicas en las células objetivo). Se dan cambios en el metabolismo de la célula receptora (según lo que decía el mensaje) Finalmente se elimina la señal. La señal puede ser eliminada por difusión en el espacio extracelular, inactivación enzimática o recaptación.
Ligandos y Receptores
Los ligandos: son los primeros mensajeros o señales químicas, son moléculas de naturaleza proteica o lípidos, estos se ponen en contacto con células vecinas o distantes.
Con receptores intracelulares: esteroideas, ácido retinoico, tiroxina. Con receptores de superficie: prostaglandinas, leucotrienos, tromboxanos, etc. Óxido Nítrico (NO) (segundo mensajero) Este es un gas diatómico hidrofóbico, sintetizado en distintas células del organismo, este tiene un papel crucial como molécula de señalización celular. Es relevante en el cerebro y en el sistema cardiovascular, es sintetizado por las óxido nítrico sintetasas. Es una molécula paracrina y en diversas patologías como la hipertensión, hipercoleresterolemia, diabetes, envejecimiento, tabaquismo, disfunción eréctil e insuficencia cardiaca, se han observado alteraciones en la producción o en la biodisponibilidad del NO. Actúa como un mediador local difundiendo a través de la membrana de célula diana y activando la guanilato ciclasa (su receptor) para producir GMPc en la célula diana (segundo mensajero).
Receptores de señal
Los receptores de señal pueden ser extracelulares e intracelulares. Receptores extracelulares Son receptores que reciben las señales hidrofílicas también llamados receptores de superficie o membranales, son proteínas integrales específicas para un tipo de ligando. Estos pueden estar apagados o encendidos .Se dividen en 3 grupos: Los que abren canales iónicos por efecto del ligando (ionotrópicos) estos se abren y se cierran por medio de señales de neurotransmisores. Receptores con actividad de tirosina cinasa. Receptores con actividad enzimática intrinseca Acoplados a proteína G: estos receptores comunican con el citoplasma estimulando proteínas G haciendo que estas se activen
o desactiven. Una proteína G activada envía señales a enzimas AMPc o GMPc las cuales actúan sobre ATP o GMP (estos ya serían 2dos. Mensajeros) así se activa a la fosfolipasa C-β, la cual hidroliza la fosfatidilinositol bifosfato (PIP2) generando dos mediadores intracelulares: el inositol trifosfato (IP3) y el diacilglicerol. (para complementar esto les recomiendo leer “ciclo de activación/inactivación de proteínas G” página 434 y 436 del libro de biología ;). El AMPc se origina a partir de ATP por acción de la adenilciclasa. Segundos mensajeros: las señales membranales hacen uso de segundos mensajeros como: AMPc: formado por la adenil ciclasa, a partir de ATP. Las fosfodiesterasas (PDEs) se encargan de su degradación. Sus efectos son variados. GMPc: formado por la guanil ciclasa a partir de GTP. Las fosfodiesterasas (PDEs), se encargan de su degradación. Ca++^ : se libera del RE o entra al citosol desde el espacio extracelular al abrirse canales específicos. Inositol trifosfato (IP3) : que libera Ca2+ desde el REL, incrementando así la concentración de Ca2+ en el citosol se forma a partir del fosfatidilinositol. (esto lo hace la fosfolipasa C). Diacilglicerol (DAG) que permanece en la membrana plasmática y activa la quinasa C, se forma a partir del fosfatidilinositol. (esto lo hace la fosfolipasa C).
Este tipo de comunicación ocurre entre células a distancia, por medio de señales químicas llamadas hormonas. Aquí la glándula secretora emite una señal endocrina por medio de una hormona la cual viaja por el torrente sanguíneo hasta llegar a las células blanco que se encuentran a distancia. Comunicación Parácrina Esta comunicación se da entre células cercanas o adyacentes. Un claro ejemplo de la comunicación parácrina es la sinapsis entre neuronas donde la región presináptica libera el neuro transmisor, pasa por un pequeño espacio sinaptico y llega a la región post sinática donde se recibe el estímulo. Esto se da por ejemplo entre la neurona y la fibra muscular. Comunicación Yuxtácrina (por contacto) Esta se da por contacto con otras células o con la matriz extracelular mediante moléculas de adhesión celular. Esto significa que las células están literalmente unidas cuando se da este tipo de comunicación. Esta comunicación entre células homólogas (iguales) es esencial para
el control celular y la formación de tejidos. Entre células heterólogas (diferentes) es muy importante para el reconocimiento que realiza el sistema inmune. Esta comunicación ocurre mediante uniones tipo GAP (canal). Un ejemplo de este tipo de señalización es cuando las Cadherinas-Integrinas (como se observa en la imagen) regulan la comunicación al contacto célula célula o célula -matriz. Comunicación Autócrina Aquí la célula responde a señales liberadas por ella misma. Señalización celular y apoptosis La traducción de señal no es únicamente importante en la regulación hormonal y la señalización de los factores de crecimiento. La señalización celular también regula un tipo programado de muerte celular o apoptosis. Cuando las células son infectas por patógenos o cuando los leucocitos alcanzan el final de su ciclo vital, son eliminados a través de apoptosis. En la apoptosis primero se condensa la cromatina y se retrae el citoplasma, luego se fragmenta el núcleo, el DNA y se forman envaginaciónes y fragmentación celular finalmente las células fagocíticas destruyen los cuerpos apoptóticos. Algunas señales de apoptosis son el factor de necrosis tumoral y la CDp5/Fas. Hay proteínas anti-apoptóticas como la Bcl-2 y hay proteínas pro-apotóticas como la Bax y Bad. Las mitocondrias desencadenan la apotosis liberando citocromo C al citosol.
hidrógeno y este apareamiento está condicionado químicamente de forma las bases solo pueden unirse: A con T , G con C. La replicación del AND ocurre durante la fase S previo a la mitosis, la replicación de ADN ocurre más rápidamente en procariotas y los virus necesitan de células huésped para llevarlo a cabo. La síntesis de ADN/replicación se da en dirección 5´-3´ en ambas hebras, esta replicación es complementaria y antiparalela. Esquemas de duplicación Hebras continuas o discontinuas Durante la replicación una hebra recibe el nombre de rezagada o discontinua ( a esto también se le conoce como fragmentos de Okasaki) ya que su replicación se lleva a cabo por partes. La otra hebra se denomina continua ya que su replicación se lleva a cabo sin pausas. Enzimas que actúan en la replicación de ADN Construye las cadenas de ADN Polimerasas ADN (especialmente la III y la exonucleasa) Se encargan del enrollamiento y desernrrollameinto del ADN Topoisomerasas Corta el enlace 3’-5´fosfodiester Topoisomerasa I
Corta hebras enredadas de ADN Topoisomerasa II Se unen a las hebras molde evitando que vuelvan a enrollarse proteínas SSBP Rompe enlaces de hidrógenos Helicasa Síntesis de RNA cebador ó primario Primasa RNA cebador DNTP’s/A,C,T y G/uracilo Une fragmentos de Okazaki por medio de enlaces covalentes Ligasa Reemplaza los espacios iniciadores o cebadores de los fragmentos de okazaki polimerasa I Polimerasas en Procariotas DNA Polimerasa I: sintetiza ADN, exonucleasa (correctora) 3’-5´y de 5 ´-3´, sustituye a los cebadores. DNA polimerasa III: sintetiza ADN en ambas cadenas, exonucleasa 3´- 5´. Polimerasas en Eucariotas Polimerasa Alfa: se encarga de la síntesis de ADN nuclear, reparación del ADN en ambas hebras y remueve los iniciadores de ARN. Polimerasa Gama: se encarga de la síntesis de ADN mitocondrial. Polimerasa Delta: se encarga de la síntesis nuclear y de la reparación del ADN exonucleasa 3´-5´. Polimerasa Epsilon: se encarga de la síntesis de ADN nuclear, la reparación del ADN, exonucleasa 3´- 5´ Reparación de ADN Al detectarse un daño en la hebra de ADN, se debe quitar la zona quitada, esto lo hace la endonucleasa. Seguidamente llega la polimerasa I e inserta los nucleótidos corretos, finalmente al estar listo el segmento corregido se une con el resto por medio de la ligasa.