¡Descarga BIOQUÍMICA II y más Apuntes en PDF de Bioquímica solo en Docsity!
TEMA 1
INTRODUCCIÓN AL METABOLISMO
El metabolismo es el conjunto de reacciones químicas que ocurren en las células de los seres vivos.
Da respuesta a dos preguntas:
_- ¿Como la célula extrae energía de su entorno físico y de los nutrientes para realizar sus procesos vitales?
- ¿Como la célula sintetiza las macromoléculas y los sillares estructurales?_
Las reacciones metabólicas están catalizadas por las enzimas. Se organizan en rutas metabólicas que tienen
un sustrato inicial y un producto final.
Los seres vivos comparten muchas rutas metabólicas, lo que es una prueba de que todos los organismos
tienen un mismo origen evolutivo.
Ejemplo: la glucólisis en bacterias o vertebrados.
Todos los seres vivos requieren para un metabolismo una fuente de energía. En función de esto se clasifican
en:
o Autótrofos : son capaces de generar materia orgánica a partir de moléculas inorgánicas sencillas, como las
plantas o ciertos tipos de bacterias
o Quimioautótrofos: son capaces de utilizar compuestos inorgánicos oxidados (CO2) como sustratos
para obtener energía y utilizarla en el metabolismo respiratorio. Es una facultad exclusiva de las super
bacterias conocida con el nombre de quimiosíntesis.
o Fotoautótrofos: son organismos (especialmente plantas) que efectúan fotosíntesis para obtener
energía. Los organismos fotoautótrofos utilizan la energía de la luz solar para fijar el dióxido de carbono
(CO2); este es combinado con agua (H2O) formando PGAL (fosfogliceraldehído)
o Heterótrofos :se alimentan de sustancias inorgánicas sintetizadas por otros organismos.
Metabolismo = anabolismo + catabolismo
Anabolismo: son los procesos del metabolismo que tienen como resultado la síntesis de moléculas complejas
a partir de moléculas más sencillas, por lo que recibe también el nombre de biosíntesis , invirtiendo energía
(en forma de ATP o poder reductor, NADH o FADH2). Las reacciones de anabolismo son reacciones de
reducción
Catabolismo: consiste estructuras en la transformación de complejas en moléculas sencillas consiguiendo
energía disponible en forma de ATP y poder reductor del catabolismo son reacciones de oxidación.
El poder reductor es necesario para la síntesis de cualquier tipo de molécula. La energía en forma de ATP
también sirve en rutas biosintéticas.
El catabolismo es exergónico y el anabolismo es endergónico.
Las vías catabólicas son convergentes, es decir, a partir de diferentes reactivos se produce un único producto
intermediario, que es común a todos los productos iniciales. Este producto común es el acetil CoA. Al
contrario, las vías anabólicas son divergentes. A partir de unos pocos reactivos iniciales, se producen todas
las moléculas dentro de la célula.
Las rutas anfibólicas son rutas que intervienen en el anabolismo y en el catabolismo como es el ciclo de
Krebs o el ciclo del ácido cítrico (TCA).
- La mayoría de las reacciones de una ruta metabólica están próximas al equilibrio, son reversibles.
- Cada ruta metabólica tiene al inicio una reacción de no-equilibrio que marca la direccionalidad de la
ruta.
- Las encimas que catalizan las reacciones de no-equilibrio son los puntos de control de las rutas
metabólicas.
- Las rutas metabólicas están reguladas a distintos niveles: interno, intracelular y extracelular.
- En las células eucariotas hay rutas metabólicas que están compartimentalizadas, que ocurren en
varios lugares de la célula como por ejemplo en el citosol y en la mitocondría.
Una ruta metabólica es la suma de las reacciones que convierte el reactivo en el producto final:
Ruta 1: S>A>B>C>D>P
Ruta 2: P>J>A>C>B>A>S
·En forma de ión hidruro H-
Las reacciones redox llevan asociada la variación de la energía libre asociada a un proceso redox.
ΔG = -nGΔE
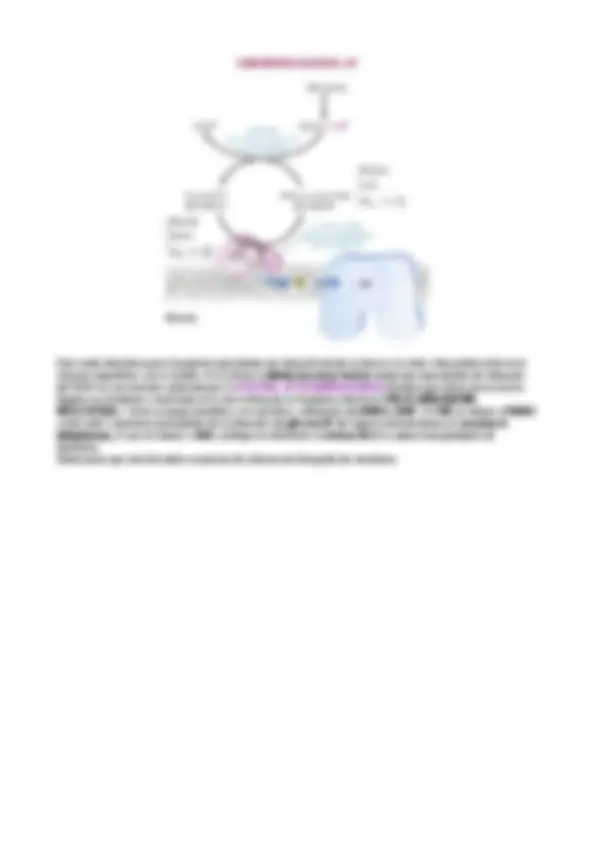
MOTIVOS RECURRENTES DEL METABOLISMO
-Transportadores activos en el metabolismo. -Tipos de reacciones (se reunen en seis clases) ·Oxidación-reducción: transferenca de electrones. ·Ligacioón requiere ATP y se forma enlaces covalentes. ·Isomerización. ·Transferencia de grupos. ·Hidrólisis. ·Adición-separación de grupos funcionales. NIVELES DE REGULACIÓN
1. Cantidad de enzima: cataliza las reacciones. - A nivel de síntesis de transcripción. -A nivel de degradación. 2. Actividad de la enzima. -Regulación alostérica. -Conversión molecular covalente reversible (fosforilación o desfosforilación) **3. Control hormonal.
- Accesibilidad de sustratos.** -Compartimentación. El metabolismo se regula modulando la actividad de las enzimas y la accesibilidad de los sustratos. Todo aquello que controla el acceso del sustrato es importante. Fe + Cu = Fe + Cu
- En forma de átomos de H AH 2 = A + 2e-^ + 2H+ AH 2 + B = A + BH 2
- En forma de ión hidruro H
NAD(P)
1 / 2 O 2 → R-CH 2 -OH Las reacciones redox llevan asociada la variación de la energía libre asociada a un proceso redox. ΔG = -nGΔE OXIDACIÓN REDUCCIÓN Pérdida de electrones Ganancia de electrones Ganancia de O 2 Pérdida de O 2 Pérdida de H 2 Ganancia de H 2 Liberación de energía Captación de energía Exoergónica Endoergónica MOTIVOS RECURRENTES DEL METABOLISMO
- Transportadores activos en el metabolismo
- Tipos de reacciones (se resumen en 6 clases) o Óxido-reducción: transferencia de electrones o Ligación requiere ATP y se forman enlaces covalentes o Isomerización o Transferencia de grupos o Hidrólisis o Adición-separación de grupos funcionales NIVELES DE REGULACIÓN
- Cantidad de enzima: cataliza las reacciones
- A nivel de síntesis de transcripción
- A nivel de degradación
- Actividad de la enzima:
- Regulación alostérica
- Conversión molecular covalente reversible (fosforilación o defosforilación)
- Control hormonal
- Accesibilidad de sustratos
TEMA 2
METABOLISMO DE CARBOHIDRATOS
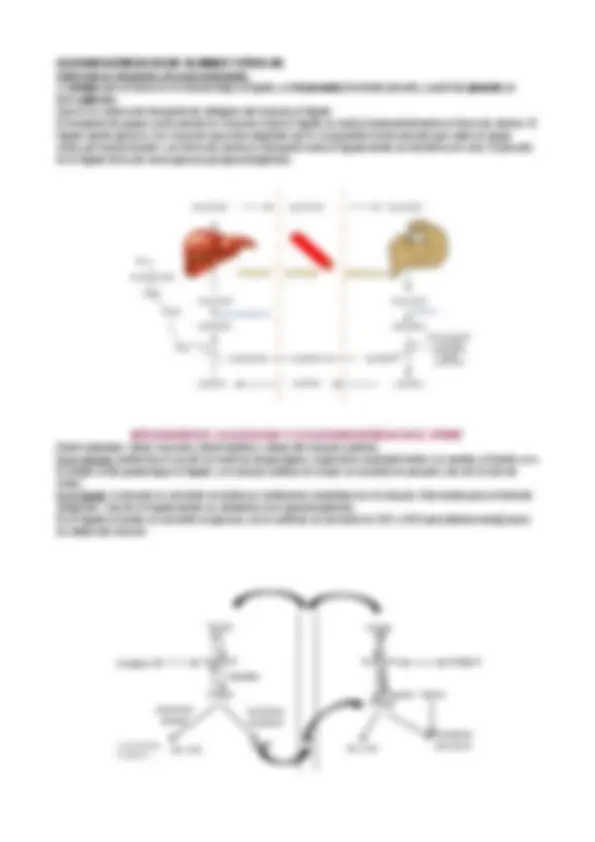
Un lugar central lo ocupa la glucosa. Puede proceder del glucógeno que es un polisacáridos de reserva animal. La ruta de las pentosas fosfato origina ribosa y NADPH, a partir de esta se puede regenerar glucosa. La glucosas también se puede sintetizar en la célula a partir de moléculas precursoras de carbohidratos que no son carbohidratos (glucógenogénesis). La glucosa se degrada en la glucólisis para convertirse en piruvato, en condiciones aerobias (por medio de la acetilCo-A ) que entra en el ciclo de Krebs, pero el acetilCo-A también interviene en la síntesis de grasas cuando la ingesta de glucosa es muy grande (por eso se dice que el azúcar engorda) DIGESTIÓN Alimentos con carbohidratos: polisacáridos, oligosacáridos (glucógeno, almidón, glucosa, sacarosa, almidón) La digestión comienza en la cavidad oral y la enzima que actúa es la amilasa salibar que convierte los polisacáridos en
carbohidratos más pequeños rompiendo enlaces ß1-
En el estómago las amilasas pancreáticas que actúa sobre los carbohidratos dan como resultado final: maltosa , dextrinas (mirar definición) y otros. Epitelio del intestino delgado se segregan: dextrinasa, disacaridasas. Enzimas que acabarán por degradar estos carbohidratos a sustancias más pequeñas, que ya podrán entrar en las diferentes rutas metabólicas en las que sean requeridas. TRANSPORTE DE GLUCOSA AL INTERIOR CELULAR. Glucosa y galactosa entran mediante transporte activo (simporte), fructosa mediante difusión facilitada. El transporte de glucosa es a favor de gradiente pero es un transporte activo, específicamente es un simporte ya que es simultáneo al transporte del Na+. Se necesita ATP para transportar el sodio mediante la bomba de sodio potasio, El transporte en si no gasta ATP pero hay que mantener el transporte de Na+-K+ (que ocurre solo en el epitelio intestinal) En las células entra a través de un receptor de membrana llamada GLUT , la entrada de la glucosa es por difusión facilitada, a favor de gradiente porque en el interior de la células la concentración es más baja que fuéra. Podría GLUT intervenir en el proceso contrario? Cuando se hecha fuera la glucosa es porque fuera de la célula su concentración es menor, entonces el gradiente es invertido. El sistema funciona exactamente igual, el GLUT también interviene en la exteriorización. GLUT CINÉTICA: análoga a una reacción enzimática. la velocidad inicial de la entrada de glucosa es: V0= Vmax ·[S]/ Kt + [S] mediante la digestión se convierten en monosacáridos que sufren absorción en el intestino pasando de esta manera a la sangre. La sangre los conduce a los tejidos participando así en el metabolismo. Los carbohidratos son ingeridos por la cavidad oral en la que la amilasa salivar empieza a degradarlos, de ahí pasa al estómago y al intestino, donde la amilasa rompe los enlaces α1-4 y las ramificaciones por las amilasas pancreáticas (polisacáridos maltosa y otros disacáridos). Al final de toda esta ruta se generan monosacáridos que se absorben por absorción intestinal. La glucosa y la galactosa son transportados por un transportador activo por un sistema simporte ligado al Na+^ para mantener el gradiente de Na+^ por una bomba Na+/k+. La entrada de glucosa se lleva a cabo por difusión facilitada pero hablamos de transporte activo porque hay que consumir ATP. La fructosa también entra por difusión facilitada. Cuando la glucosa llega a la sangre se distribuye por moléculas transportadoras de glucosa, denominadas GLUT, que transportan la glucosa al interior de la célula. TIPOS GLUT 1 Un receptor que está siempre captando glucosa. La lleva a todos los tejidos y capta glucosa aunque su concentración sea muy baja. Su Km es muy baja, aproximadamente 1mM, es decir que se saturan enseguida. GLUT 2 Estimula la segregación de insulina en el hígado al captar la glucosa (hígado y músculo almacena glucógeno). Almacenan glucógeno. GLUT 3 Un receptor que está siempre captando glucosa. La lleva a todos los tejidos y capta glucosa aunque su concentración sea muy baja. Su Km es muy baja, aproximadamente 1mM, es decir que se saturan enseguida. GLUT 4 Está en el músculo (consumidor de glucosa) y tejido adiposo (transfiere en acil-CoA- ac.grasos-triacilglicéridos) GLUT 5 Transportador mas bien de fructosa, está en el intestino delgado.
TEMA 3
LA GLUCÓLISIS Y SU REGULACIÓN.
Digestión metabólica de la glucosa dentro de la
células
Glucolisis : conjunto de reacciones que tranforman glucosa en dos moléculas de piruvato con producción neta, concomitante en dos moléculas de ATP.
- Piruvato es un metabolito central que va a actuar en muchos procesos. -Es una ruta metabólica que ocurre en el citosol. -Es una ruta universal (animales. plantas, microorganismo) -Romper los glúcidos. -Papel muy importante dentro de metabolismo. -Conectada con otras rutas metabólicas. -Es la que mejor se conoce. Para una fuente de energía rápida, la glucosa se convierte en piruvato por la glicólisis, posteriormente se puede convertir en condiciones anaeróbica ( fermentación ) en lactato (fermentación láctica) o etanol (fermentación alcohólica). En condiciones aeróbicas, el piruvato puede oxidarse completamente a CO2,, lo que genera mucho más ATP. HISTORIA Los hermanos Buchner observan la conversión de sacarosa en etanol en presencia de extractos de levadura. A este proceso lo llamaron fermentación. Hans Buchner y Eduard Buchner hicieron en 1897 un descubrimiento clave, totalmente por accidente. Los Buchner estaban interesados en comercializar extractos de levadura libres de células para un posible uso terapéutico. Estos extractos se conservaron con sacarosa. Resultado: La sacarosa fermentó rápidamente hasta alcohol en el jugo de levadura por tanto demostraron que la fermentación podía tener lugar fuera de las células vivas. En 1940 estaba aclarada toda la vía glucolítica: Meyerhof, Emben, Parnas, Cari y Gertey Cori, warburg. [Este experimento supuso: el fin del dogma VITALISTA. gracias a esto la biología se liberaría de otro fragmento del miticismo. La rotura del azúcar en CO2 y alcohol ya no es el efecto de un principio vital. También el nacimiento de la bioquímica como ciencia] TEMA III: GLICÓLISIS Y SU REGULACIÓN
- Glicólisis Es la secuencia de reacciones que convierte una molécula de glucosa en dos moléculas de piruvato con la producción neta concomitante en dos moléculas de ATP.
- Ocurre en el citosol
- Es universal: ocurre en animales, plantas y microorganismos.
- Su nombre significa “romper el dulce”
- Tiene un papel central en el metabolismo energético y está interconectada con otras rutas metabólicas
- Se conoce bien. Para una fuente de energía rápida, la glucosa se convierte en piruvato por la glicólisis, posteriormente se puede convertir de forma anaeróbica ( fermentación ) en lactato ( fermentación láctica ) o etanol ( fermentación alcohólica ). En condiciones aeróbicas, el piruvato puede oxidarse completamente a CO2,, lo que genera mucho más ATP.
- Historia Hans Buchner y Eduard Buchner hicieron en 1897 un descubrimiento clave, totalmente por accidente. Los Buchner estaban interesados en comercializar extractos de levadura libres de células para un posible uso terapéutico. Estos extractos se conservaron con sacarosa. Resultado: La sacarosa fermentó rápidamente hasta alcohol en el jugo de levadura por tanto demostraron que la fermentación podía tener lugar fuera de las células vivas. En 1940 estaba aclarada toda la vía glucolítica: Meyerhof, Emben, Parnas, Cari y Gertey Cori, warburg. FASES Y REACCIONES
1ª fase de la glucólisis
Está catalizada por la hexoquinasa o Glucoquinasa que marcan la glucosa con un Pi , el cual es esterificado con el grupo hidroxilo del carbono 6.
FASES Y ETAPAS DE LA GLUCÓLISIS 1º-> inversión de ATP para finalmente obtener más cantidad del mismo. 2º-> rentabilidad, obtención de productos de glucólisis. 1º FASE 1º Reacción. Catalizada por la HEXOQUINASA (HK) o GLUCOQUINASA marca la glucosa con un Pi, el cual es esterificado con un grupo hidroxilo del carbono-6. Enzima catalitica intermediaria de la reacción que convierte la glucosa en glucosa 6-P mediante la transferencia de fosforilos (fosforilación) Reacción de no equilibrio (ΔG: -40 kcal/mol) por tanto es un punto de control, el primero en esta ruta. HEXOQUINASA: enzima formada por 4 isoenzimas (enzimas catalíticas cuya acción es la misma pero estructuralmente diferentes porque están sintetizadas por genes diferentes). La hexoquinasa IV también se denomina GLUCOQUINASA y actúa exclusivamente (o casi) en el hígado. Cuando la glucosa entra en la células es inmediatamente fosforilada por la HK por 2 motivos: -Evitar que la glucosa salga otra vez de la célula. -La glucosa 6-P es la molécula activa en el metabolismo (glucólisis) Diferencia entre HEXOQUINASA y GLUCOQUINASA: principalmente es la cinética, la glucoquinasa tiene una Km muy alta, frente a la Km baja de las otras hexoquinasas. Hexoquinasas -> específicas de cualquier hexosa (además de la glucosa) Glucoquinasas -> solo fosforilan la glucosa. La síntesis de glucoquinasas es producida por la insulina, enzima específica de la glucosa. De manera que la glucosa se convierte en glucosa 6P, que al ser una reacción de no equilibrio se gasta un ATP en el proceso. La glucosa-6P no puede salir de la célula porque está cargada eléctricamente –, de manera que el proceso de fosforilación de la glucosa sirve de mantener la glucosa y todos sus intermediarios dentro de la célula. La diferencia entre la glucoquinasa y la hexoquinasa es la Km, de manera que la Km de la hexoquinasa es baja mientras que la de la glucoquinasa es mucho mayor y está en el hígado, de manera que se fosforila muy rápido.
- La síntesis de glucoquinasa es producida por la insulina, es una enzima específica de la glucosa.
- La hexoquinasa siempre está en las células y fosforila a otras hexosas. Después tiene lugar la isomerización , en la que se reordenan los átomos para convertir a una aldosa en una cetosa. Esta reacción está próxima al equilibrio por tanto es reversible. La enzima que cataliza esta reacciñon es la fosfoglucosa isomerasa, que transforma la glucosa 6P a fructosa. Otra de las reacciones que ocurren es la fosforilación, en la que la enzima necesaria es la fosfofructoquinasa (PFK), que fosforila el C1 de la fructosa 6P a fructosa 1,6P 2. Quien cede ese fosfato es el ATP.
A efectos prácticos es
necesaria la una enzima
posea una afinidad alta
por el sustrato (Km alta) para que se
permita la entrada
masiva de glucosa en la
célula
5º Reacción La dihidroxiacetona fosfato se isomeriza a gliceraldehído-3P por la actividad catalítica de la enzima TRIOSA FOSFATO ISOMERASA. Es una reacción próxima al equilibrio. Es el punto final de la primera fase. Balance: se han invertido 2 moléculas de ATP para activar la molécula de glucosa, provocando su rotura en 2 molécula de 3 carbonos ( TRIOSAS ), se eleva el contenido de ΔG de los intermediarios y las cadenas de C de todas las hexosas metabolizadas se convierten en un producto común: Gliceraldehído-3-P. 2º FASE 6º Reacción Gliceraldehído-3-P (los 2) reaccionan con el NAD+ (coenzima de óxido-reducción) + Pi (fosfato inorgánico, el cual fosforila al gliceraldehído-3-P) para generar NADH+ + 1,3-Bifosfoglicerato (2 fosfatos, uno de ellos, el que genera en la reacción en una molécula con un gran potencial de transferencia de grupos fosfato) Es una reacción próxima al equilibrio y por tanto es reversible. Esta reacción es catalizada por la GLICERALDEHÍDO-3P DESHIDROGENASA. Se forman dos moléculas de NADH porque se oxidan dos moléculas de gliceraldehído-3P mediante una molécula de NAD+ casa una, así como, 2 Pi que fosforilan una molécula cada uno. 7º Reacción Primera reacción formadora de ATP (2) lo que ocurre es que la FOGLICERATO QUINASA (PHK) desfosforila el 1,3- Bifosfoglicerato (el cual posee un fosfato que obtiene cuando un aldehído se convierte en un ácido) de manera que esa molécula posee un alto potencial de transferencia de ese grupo fosfato pasando, por tanto, a 3-fosfoglicerato , fosforilando un ADP a ATP. Recordemos que se forman dos moléculas de ATP, porque, inicialmente partimos de 2 moléculas de gliceraldehído-3P. Es una reacción próxima al equilibrio.
8º Reacción Del 3-fosfoglicerato se pasa a 2-fosfoglicerato , reacción catalizada por la FOSFOGLICERATO MUTASA (PGM) la cual tiene el poder de reorganizar los átomos de la primera para formar la segunda. Es una reacción próxima al equilibrio. 9º Reacción La 2-fosfoglicerato es transformada (deshidratada) a fosfoenolpiruvato (PEP) por la acción de la ENOLASA. En este proceso se produce H2O. 10º Reacción La PEP tiene un gran potencial de transferencia de grupo fosfato, con lo cual fosforila una ADP para formar ATP (2) Como consecuencia de esta transferencia el PEP se convierte en PIRUVATO. Última reacción de la glucólisis, es una reacción de no-equilibrio y está catalizada por la PIRUVATO QUINASA (PK). Balance: rendimiento neto de esta fase son 2 ATP (por moléculas de glucosa implicada) y energía en forma de 2 NADH por moléculas de glucosa.
SACAROSA (azúcar de mesa) Disacárido hidrolizado por la SACARASA y se convierte en glucosa y fructosa que se integra de dos formas en la glucólisis: ~ a través de la HEXOQUINASA que da lugar a la glucosa-6P directamente. ~ se convierte en fructosa-1P (que no es un metabolito de la R.M.) mediante la HEXOQUINASA (fructoquinasa). Posteriormente, la fructosa-1P se rompe por la ALDOLASA en dos triosas ( gliceraldehído + triosa-P que ya forma parte de la glucólisis) MANOSA Monosacárido que se fosforila a manosa-6P mediante la HEXOQUINASA. La manosa-6P es convertida a fructosa-6P mediante la FOSFOMANOSA ISOMERASA. FRUCTOSA Monosacárido que puede degradarse de diferentes fomas según el tejido en el que se encuentre. ~ Hígado: la fructosa es fosforilada a fructosa-1P por la FRUCTOSA QUINASA esta reacción consume 1 ATP. La fructosa-1P es fosforilada por la ALDOLASA en 2 triosas (dihidroxiacetona fosfato -> se convierte en gliceraldehído-3P mediante TRIOSAFOSFATO ISOMERASA; y el gliceraldehído-3P -> metabolito de la glucólisis) ~ Tejido adiposo : Fructosa fosforilada a fructosa-6P directamente y esta entra en la R.M, la reacción catalizada por la HEXOQUINASA. GALACTOSA Primero la galactosa es fosforilada por la GALACTOQUINASA a galactosa-1P. La galactosa y la glucosa son esteroisómeros epímeros (carbono quiral), por tanto no se puede pasar directamente de galactosa-1P a glucosa-6P o fructosa-6P. La FOSFOGLUCOMUTASA (PGM) intercambia la galactosa con la glucosa el complejo UDP-glucosa , formando UDP-galactosa. La glucosa-6P sigue la ruta de la glucólisis, y la UDP-galactosa es transformada en glucosa por la actividad de la UDP-GALACTOSA EPIMERASA. Esta reacción regenera la UDP-glucosa La PGM tiene un grupo fosfato en su centro activo. La enzima realmente intercambia el grupo fosfato, uniendo el fosfato con el C6 de la glucosa. Por una fracción de segundo la glucosa tiene 2 fosfatos unidos. Entonces la PGM se regenera uniendo el fosfato en posición 1 a su centro activo. Este mecanismo se observa cuando el centro activo de la enzima se marca radiactivamente, entonces se observa el centro activo antes de la reacción y después de esta y como se esperaba: NO ES IGUAL. Patologías: ~ Deficiencias en culaquiera de las tres enzimas que intervienen en la degradación de la galactosa ( GALACTOQUINASA, UDP-GLUCOSA: GALACTOSA-1P URIDILTRANSFERASA, FOSFOGLUCOMUTASA (PGM) ) producen la enfermedad conocida como galactosemia, en los seres humanos.
- En la galactosemia causada por una deficiencia de GALACTOQUINASA se encuentran elevadas concentraciones de galactosa en la sangre y en la orina. Los recién nacidos desarrollan cataratas debido al depósito de galactitol en el cristalino del ojo. Los síntomas de esta enfermedad son relativamente suaves y una limitación estricta de la galactosa en la dieta disminuye notablemente su gravedad.
- En la galactosemia causada por una deficiencia de la UDP-GLUCOSA: GALACTOSA-1P URIDILTRANSFERASA es más grave; se caracteriza por poco crecimiento den los niños, anomalías del habla, deficiencia mental y lesión hepática que puede ser fatal, incluso cuando se suprime la galactosa de la dieta. - En el tejido adiposo : La fructosa es fosforilada por la hexoquinasa (HK) a fructosa 6P, que sigue directamente la ruta de la glucólisis.
Galactosa
Primeramente la galactosa es fosforilada por la acción de la galactoquinasa , a Galactosa 1P. La galactosa y la glucosa son estereoisómeros epiméros (un carbono quiral), por tanto no se puede pasar directamente de galactosa 1P a glucosa 6P o fructosa 6P. La fosfoglucosmutasa ( PGM ) intercambia la galactosa con la glucosa del complejo UDP-glucosa, formando UDP-galactosa. La glucosa 6P sigue la ruta de la glucólisis, y la UDP-galactosa es transformada en glucosa por la actividad de la epimerasa. Esta reacción regenera la UDP-glucosa. UDP-galactosa UDP-glucosa UDP-Gal epimerasa La PGM tiene un grupo fosfato en su centro activo. La enzima realmente intercambia el grupo fosfato, uniendo el fosfato del centro activo con el carbono 6 de la glucosa. Por una fracción de segundo, la glucosa tiene dos fosfatos unidos. Entonces la PGM se regenera uniendo el fosfato en posición 1 a su centro activo. Este mecanismo se observa cuando el centro activo de la enzima se marca radiactivamente, entonces se observa que el centro activo antes y después de la reacción no es igual. PATOLOGÍAS
- Intolerancia a la lactosa: deficiencia de la lactasa.
- Galactosemias: la más común es la deficiencia por la galactosa 1P uridil transferasa cuyas consecuencias son graves: retraso mental, cataratas, diarreas, cirrosis, vómitos. Las cataratas porque en el cristalino la galactosa se convierte en galactitol que produce un entubamiento del cristalino y por tanto provocan las cataratas.
- En la galactosemia causada por un déficit de la UDP-GALACTOSA EPIMERASA conduce a síntomas similares pero es menos grave cuando se controla la galactosa de la dieta. LACTOSA Disacárido que se degrada mediante la LACTASA en glucosa y galactosa. Patologías
- Deficiencia en LACTASAS provoca una patología que vuelve a los que la sufren intolerantes a la lactosa. Es una de las patología más comunes, apenas tiene gravedad, tiene una causa genética entre otras (agresión de la mucosa intestinal) En un intestino normal la glucosa sigue una ruta normal, pero, cuando una persona es intolerante la lactosa es fermentada por bacterias de la flora intestinal, de esa fermentación se generan una serie de gases (metano, hidrógeno…) además, se forman otros dos producto: lactato y lactoda ; todo esto provoca que entre agua al intestino para diluir el lactato y consecuentemente, esto deriva en una diarrea. ¿Es natural que los adultos beban leche?
- En general, los mamíferos solo toman leche durante la lactancia, por tanto en muchas poblaciones del mundo so muchas las personas intolerantes a ella (Sureste asiático, Japón…) DESTINOS DEL PIRUVATO En general puede seguir tres rutas catabólicas: Condiciones aerobias (en tejidos aerobios) => descarboxilación oxidativa y complejo de la PIRUVATO DESHIDROGENASA. En este ámbito la glucólisis solo constituye el primer paso de la degradación de la glucosa. El piruvato se oxida perdiendo su grupo carboxílico en forma de CO2 y da lugar al Acetil-CoA , el cual se oxida por completo a CO2 por medio del ciclo de Krebs. Los electrones de estas oxidaciones pasan al oxígeno a través de una cadena de transporte de electrones en la membrana de la mitocondria que finalmente dará como producto H2O. La energía que proviene de las reacciones de transferencia de electrones impulsa la síntesis de ATP (tb en la mitocondria). Efecto Pasteur Inhibición de la glucólisis por la respiración (en presencia de oxígeno) o la inhibición anaerobia de la glucolisis. El proceso va más lento. Lo observa en las levaduras. Ocurre porque en condiciones aerobias se obtiene mucho ATP consumiendo poca glucosa, sin embargo, en condiciones anaerobias se produce la misma cantidad de ATP pero con un mayor consumo de glucosa. Condiciones anaerobias => FERMENTACIÓN: Proceso en que se extrae energía en forma de ATP pero sin consume de oxígeno ni cambio en las concentraciones de NAD* o NADH. Es un proceso necesario para la obtención de ATP y para la reoxidación de NADH a NAD* y así evitar que se detenga la glucólisis. 2 procesos fundamentales. ~ Fermentación láctica. piruvato --(LACTATO DESHIDROGENASA)--> 2 lactato Se produce en el músculo con contracción vigorosa (el cual funciona con concentraciones de oxígeno bajas (hipoxia)), en eritrocitos y en algunas células y microorganismos (que convierten la glucosa en lactato incluso en condiciones aerobias) Es la conversión de la glucosa en ácido láctico ( lactato ) Cuando los tejidos animales no se les puede suministrar oxígeno suficiente para mantener la oxidación aeróbica del piruvato y la regeneración del NADH a NAD* (siendo este el receptor de electrones principal para la oxidación del piruvato) producidos en la glucólisis. En estas condiciones el piruvato es reducido a ácido láctico, aceptando los electrones del NADH y regenerándose de este modo el NAD * (de este modo la glucólisis puede continuar). La reducción del piruvato está catalizada por la LACTATO DESHIDROGENASA (LDH) y es una reacción que produce ATP, está próxima al equilibrio (reversible) por eso puede producirse en sentido contrario ( lactato-piruvato ) Por eso, cuando el lactato es producido en exceso no se acumula, se recicla produciendo piruvato, el cual sigue dos destinos diferentes: Ciclo de Krebs. Gluconeogénesis: viaja por el torrente sanguíneo hasta el hígado, donde se produce síntesis de glucosa a partir de precursores no carbohidratos.
La acción de la ADH y CYPP2E1 contribuye a un incremento de la cantidad de acetaldehído que provoca formación de aductos: el acetaldehído reacciona con nucletótidos formando una nueva estructura dentro del DNA. DESTINO DEL ACETATO GENERADO A PARTIR DE ETANOL El acetato sale a la sangre a los tejido y se convierte en Acetil-CoA: -Puede continuar en el ciclo de Krebs ( ej: personas que no comen y beben obtienen energía así )
- Acetil-CoA se puede utilizar para la síntesis de cuerpos tetónicos (KB) FFA (ácidos grasos libres) algunos aa y colesterol (como precursor metabólico cuando este no entra en el ciclo de Krebs) EFECTOS DEL ETANOL Directos. -Sobre las membranas. -Sobre las proteínas… Indirectos. -Genera hepoxia en el hígado. -Formación de compuestos peligrosos para la célula: acetaldehído , ROS (especies de oxígeno reactivas muy peligrosas en la célula) Todas producen daños en lípidos, DNA y proteínas. -Cambios en la ratio NAD/NADH* : están presentes en todas las reacciones que intervienen en el metabolismo. El cambio en la ratio vienen provocada porque si hay mucha entrada de etanol se va a consumir mucho NAD* y, consecuentemente, se va a forman mucho NADH. Ratio normal NAD/NADH= 1000. Ratio alterada NAD/NADH=** NAD* + reactante reducido -> NADH + reactante oxidado + H* CONSUMO EXCESIVO DE ETANOL PUEDE CAUSAR PATOLOGÍAS COMO : ~ Disfunción neuronal. ~ Hígado graso ==> Acumulación de triacilglicéridos (TAG), descenso de la cantidad de RER, daño mitocondrial como consecuencia de las elevadas concentraciones de etanol, daño y muerte celular. 3 estadios:
- formación de un hígado graso por un incremento en la concentración de grasa que a su vez es provocada por el mal ratio NAD*/NADH.
- inflamación del hígado y fibrosis.
- cirrosis hepática (prolongada inflamación, incremento de la fibrosis, ralentiza el ciclo de la urea y la eliminación de amonio, retención de Na, pérdida de función hepática que puede conducir a la muerte) ~ Función gonadal ===> 1- En hombres lesión testicular; afecta a la síntesis de la testosterona y producción de esperma. Todo ello causa impotencia, ginecomastia (desarrollo de caracteres femeninos: crecimiento de mamas en el hombre), esterilidad y cambios en la distribución del pelo. [SHAKESPEARE (MACBETH, ACTO II, ESCENA II)] 2- Reducción del metabolismo de las hormonas esteroideas que produce un incremento de los estrógenos. 3- Reducción de la secreción de testosterona debido a una alteración de la relación NAD/NADH en las células de Leydig. ~ Hipoglucemia ===> Baja concentración de glucosa en sangre. El alcohol inhibe la gluconeogénesis. 1- Altera la ratio NAD/NADH* : desplaza el equilibrio de las reacciones catalizadas por LDH (LACTATO DESHIDROGENASA) y MDH (MALATO DESHIDROGENASA) hacia lactato y malato, lo que provoca una carencia de lactato y oxalacetato para la gluconeogéniesis 2- Inhibición de la gluconeogénesis sumada al ayuno nocturno y falta de desayuno, también asociada al consume de alcohol, produce la hipoglucemia. 3- Falta de glucosa en sangre y el exceso de ácido lactico en sangre (acidosis láctica, descenso del pH sanguíneo) lleva a producirse un coma etílico. El consumo de etanol afecta a la termoregulación , crea sensación de calor por la vasodilatación pero no se produce calor, sino que se pierde este mucho más rápido. En todo caso, lo más recomendable ante situaciones críticas por ingesta desbocada de etanol es el suministro de sustancias calientes con azúcar al individuo con el fin de degradar ese azúcar mediante la glucólisis y consecuentemente generar ATP (energía para que su propio cuerpo vaya aumentado su temperatura hasta controlar su termoregulación)
REOXIDACIÓN DEL NADH CITOSÓLICO El NADH generado en la glucólisis debe reoxidarse y generar NAD* Si esto no es así, la sexta reacción de la glucólisis, catalizada por la GLICERALDEHÍDO-3P DESHIDROGENASA , no puede ocurrir, porque la enzima no tiene cofactor oxidado para aceptar los H. Hay dos formas de reoxidar NADH glucolítico:
- Mediante las fermentaciones:
- fermentación láctica: piruvato --(LACTATO DESHIDROGENASA)--> 2 lactato
- fermentación alcohólica: piruvato --(PIRUVATO DESCARBOXILASA)--> acetaldehído -- (ALCOHOL DESHIDROGENASA )--> etanol
- Mediante las lanzaderas: constituyen una estrategia aerobia de regeneración del NADH + H. El NADH no puede atravesar la membrana mitocondrial para ceder sus electrones a la cadena respiratoria, estas lanzaderas se lo permiten. Existen 2 tipos principales de lanzaderas: LANZADERA MALATO-ASPARTATO Esta lanzadera se encuentra en el hígado, en los riñones y en el corazón para transportar equivalentes de reducción desde el NADH citosólico a la matriz mitocondrial. 1) El NADH del citosol (espacio intermembrana) pasa dos equivalentes de reducción al oxalacetato produciendo malato. 2) El malato atraviesa la membrana interna por el transportador malato-alfa-cetoglutarato mediante un sistema de antiporte.*
- En la matriz el malato pasa por dos equivalentes de reducción al NAD* ; el NADH resultante es oxidado por la cadena transportadora de electrones. El oxalacetato formado a partir del malato no puede pasar directamente al citosol.
- Primero se transamina a aspartato.
- Este aspartato puede salir directamente a través del transportador glutamato-aspartato.
- En el citosol se regenera el oxalacetato , con lo que se completa el ciclo. Nucleótido nicotiamida transhidrogenasa
El Complejo I, II, III y IV son bombas protónicas desde la matriz mitocondrial y se nutren de
NADH y FADH 2. El NADH y FADH 2 proviene sobre todo del ciclo de Krebs, aunque el NADH
también proviene de la glucolisis a través de las lanzaderas.
El poder reductor también se puede alcanzar mediante el NADPH que se da en procesos
anabólicos, este podemos convertirlo en NAD obtienendo finalmente NADH, por tanto poder
reductor.
NAD
+ NADPH
NADPH + NADP+
LANZADERAS
+ Malato-Aspartato
El NADH no puede atravesar la membrana mitocondrial por eso se necesitan las lanzaderas. El
oxalacetato tampoco puede atravesar la membrana por eso se necesitan las AAT
(aminotransferasa).
No pierde energía – los electrones pasan como del NADH a la CTE.
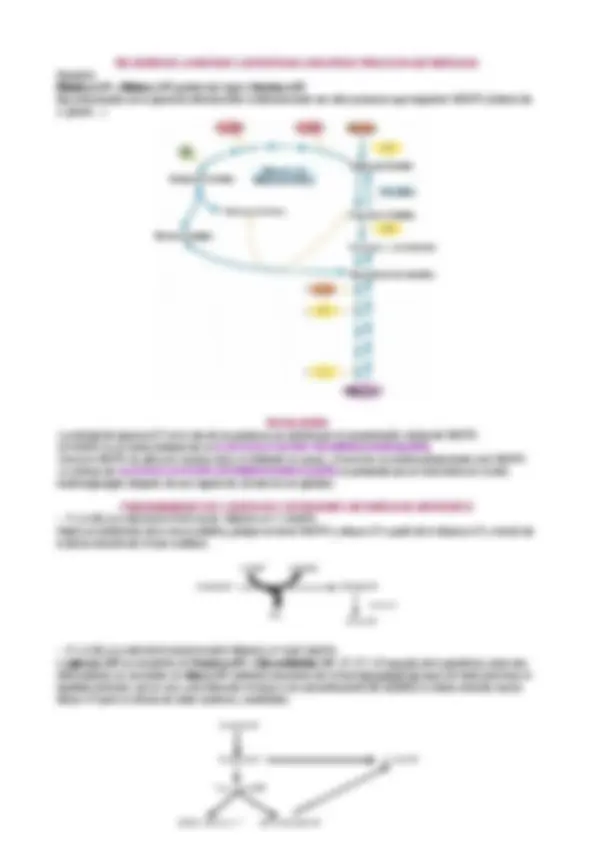
REGULACIÓN DE LA GLUCÓLISIS
La glucólisis tiende dos principales funciones:
- Generar ATP
- Sintetizar moléculas precursoras para otras rutas metabólicas.
Se hace en base a las necesidades energéticas de la células y las necesidades del aporte de precursores. En la glucólisis, existen varias reacciones que puede servir como puntos de control, debido a su ΔGº<0 (reacciones de NO-EQUILIBRIO, irreversibles), estas reacciones son: ~ Fosforilación de glucosa a glucosa-6P. ~ La fosforilación de fructosa a fructosa-1,6BP. ~ La desfosforilación de PEP a piruvato. La regulación se realiza a varios niveles: · Concentración de sustratos y productos. · Modulación alostérica: permite control inmediato del flujo a través de esa vía, la sufren las enzimas reguladoras. · Unión a proteínas moduladoras. · Conversión molecular covalente reversible: activadas por hormonas, fosfolilación y desfosforilación · Regulación a nivel transcripción. · Hormonal: control a nivel fisiológico. Enzimas reguladoras. HEXOQUINASA (HK) : enzima que cataliza la fosforilación de glucosa a glucosa-6P. Esta enzima tiene cuatro isozimas codificadas por 4 genes diferentes : I, II y III: inhibidas alostéricamente (temporal y reversiblemente) por la glucosa-6P y con una baja Km. IV (GLUCOQUINASA): no inhibida por la glucosa-6P y tiene una Km muy elevada que es la que le permite responder a grandes concentraciones de glucosa en sangre. Esta enzima, se encuentra secuestrada en el núcleo, unida a una proteína reguladora; lo que induce a su salida del núcleo es la concentración de glucosa. Cuando hay una elevada concentración de fructosa-6P la enzima vuelve a introducirse en el núcleo. También está regulada a nivel de la transcripción del gen que la codifica, una bajada en los niveles de ATP activa la transcripción de la HEXOQUINASA. FOSFOFRUCTOQUINASA (PFK o PFK-1) : enzima que cataliza la fosforilación de fructosa a fructosa-6P, es la principal reacción reguladora de la glucólisis. Permite el control coordinado de la glucólisis y de la gluconeogénesis. Esta enzima tienen varios sitios reguladores en los que se fijan activadores o inhibidores alostéricos, los principales sitios reguladores alostéricos de esta enzima son ===> la carga energética: ATP (inhibidor), AMP y ADP (activadores), citrato (inhibidor), fructosa-2,6BP (activa) CONTROL DE LA PFK-1 POR NIVELES DE ATP El ATP no es solo un sustrato de la PFK-1 sino también uno de los productos finales de la ruta glucolítica. Cuando la célula produce niveles elevados de ATP, éste inhibe la PFK-1 al fijarse a un sitio alostérico y disminuir la afinidad del enzima por la fructosa-6P. El ADP y el AMP , que aumentan su concentración cuando el consumo de ATP es superior a su producción, actúan alostéricamente para aliviar la inhibición por el ATP (son activadores). Estos efectos se combinan para producir una actividad enzimática superior cuando se acumulan ADP , AMP, fructosa-6P y disminuyen la actividad cuando se acumula ATP. La actividad de la PFK-1 aumenta cuando la relación ATP/AMP disminuye, esta relación se establece en una reacción catalizada por la ADENILATOQUINASA ADP + ADP ====> ATP + AMP Imaginemos que la célula consume ATP y aumenta la concentración de AMP, consecuentemente, la glucólisis va más rápido. l músculo Reguladores de enzimas es inhibida por la concentración de glucosa 6P de manera que cuando las s son elevadas se disminuye su actividad. nasa : es el principal punto de control. stéricos tales como: ADP como indicadores de bajos niveles de ATP que interviene en la regulación coordinada de la gluconeogénesis. trato (indican que niveles de energía están disponibles) es inhibida por acetil-CoA, ATP, ácidos grasos y alanina, fosforilación e AMPc (es un segundo mensajero de muchas hormonas que tocan en su ue produce la síntesis de este AMPc es un activador alostérico de la proteín sforila unos enzimas) or F1,6BP. Control de la PFK por niveles de ATP P son moduladores alostéricos de la PFK-1, que es receptiva al estado energético Cuando en ATP es bajo, la PFK-1 se activa, por lo contrario, cuando la de ATP es elevada inhibe su actividad. La concentración del citrato en el citosol la glucólisis. mbién es un activador por la reacción catizada por la adenilato kinasa en la que P a través de ADP cuando las concentraciones de este son elevadas. ADP + ADP AMP + ATP Resumen de los reguladores que afectan a la actividad de la PFK-1.
- En el músculo Reguladores de enzimas Hexoquinasa : es inhibida por la concentración de glucosa 6P de manera que cuando las concentraciones son elevadas se disminuye su actividad. Fosfofructoquinasa : es el principal punto de control. Activadores alostéricos tales como:
- AMP y ADP como indicadores de bajos niveles de ATP
- F2,6BP que interviene en la regulación coordinada de la gluconeogénesis.
Inhibidores:
- ATP, citrato (indican que niveles de energía están disponibles)
Fosfoquinasa : es inhibida por acetil-CoA, ATP, ácidos grasos y alanina, fosforilación dependiente de AMPc (es un segundo mensajero de muchas hormonas que tocan en su receptor y lo que produce la síntesis de este AMPc es un activador alostérico de la proteín kinasa a que fosforila unos enzimas) Y es activada por F1,6BP. Control de la PFK por niveles de ATP ATP y AMP son moduladores alostéricos de la PFK-1, que es receptiva al estado energético de la célula. Cuando en ATP es bajo, la PFK-1 se activa, por lo contrario, cuando la concentración de ATP es elevada inhibe su actividad. La concentración del citrato en el citosol también frena la glucólisis. El AMP también es un activador por la reacción catizada por la adenilato kinasa en la que se produce AMP a través de ADP cuando las concentraciones de este son elevadas. ADP + ADP AMP + ATP
CONTROL DE LA PFK-1 POR CITRATO
Regulador alostérico (inhibidor) de la PFK-1. La inhibe porque los niveles elevados de este aumentan el efecto inhibidor del ATP, con lo cual, reduce aún más el flujo de glucosa a través de la glucólisis. El citrato sirve como señal intracelular de que la célula está alcanzando sus niveles necesarios de metabolismo productor de energía y de intermediarios biosintéticos. Si la reacción de la PFK-1 se ralentiza, la glucosa se convierte en glucosa-6P y ésta se va almacenando en forma de glucógeno (en el hígado y en el músculo que es donde hay reservas de glucógeno); en otros casos, la glucosa-6P también se puede introducir en la ruta de las pentosas fosfato. Todo esto puede dar la vuelta cuando hay necesidad de ATP. CONTROL DE LA PFK-1 POR LA FRUCTOSA-2,6BP Potente activador de la PFK-1, y por tanto de la glucólisis; consecuentemente también es inhibidora de la FRUCTOSA-1,6-BIFOSFATASA 1 , así como de la gluconeogénesis. Regulador alostérico de la PFK-1 porque permite coordinar la glucólisis con la gluconeogénesis (síntesis de glucosa a partir de precursores NO CARBOHIDRATOS) La fructosa-2,6P se produce a partir de fructosa-6P, reacción catalizada por la PFK-. La cinética de la PFK-1 se ve afectada por esta enzima ( PFK-2) que actúa incrementando la afinidad de la enzima por el sustrato. Interesante es el efecto que tienen en el caso de la velocidad frente a [ATP]: el ATP es un cosustrato de la reacción de la PFK-2 , inicialmente actúa a concentraciones bajas de ATP como un activador pero a medida que va aumentado la concentración de ATP inhibe la reacción. En el marco celular, para vencer esta inhibición actúa la FRUCTOSA-1,6-BIFOSFATASA 1 para que siga funcionando la reacción de la PFK-2 que seguirá formando fructosa-2,6BP a partir de fructosa-6P. La PFK-1 forma parte de una enzima bifuncional que tiene dos actividades: Actividad PFK-1 (convierte la fructosas-6P + ATP ====> fructosa-2,6BP + ADP. Actividad FRUCTOSA-1,6-BIFOSFATASA 1 hidrolizando el fosfato de la PFK-. [¿Cuantos genes codifican para esta enzima? uno solo porque es un polipéptido] COMO SE REGULA LA ENZIMA BIFUNCIONAL Se regula por conversión molecular covalente reversible, que va a estar determinada por las dos hormonas más importantes del metabolismo de la glucosa: la insulina y el glucagón. Se produce fosforilación, desfosforilación y también de forma alostérica. (Lenihnger 5º edición 15-fig 17): Esquema que representa PFK-1 / FRUCTOSA-1,6-BIFOSFATASA 1. La conversión de una en otra se produce mediante la fosforilación que inactiva la PFK-2 y activa la FRUCTOSA-1,6-BIFOSFATASA 1. La PFK-1 es fosforilada por una PROTEÍNA QUINASA dependiente de AMPc-> se va a producir en la célula como respuesta a la secreción de glucagón que es una hormona que a su vez responde a los bajos niveles de glucosa en sangre. Se inactiva la PFK-1 y no se forma la fructosa-2,6BP Finalmente lo que se busca es que no se gaste glucosa innecesariamente. La insulina se produce como respuesta a un incremento de glucosa en sangre, produce la activación de la PROTEÍNA FOSFATASA que revierte el proceso activando la glucólisis para consumir la glucosa excedente en sangre. CONTROL SIMULTÁNEO DE LA PFK Y LA FRUCTOSA-6-BIFOSFATASA 2 POR FRUCTOSA-1,6BP (??????) El paso F1,6 --> F2,6BP no es una rama de la glucólisis como ruta metabólica sino que sirve solamente como una reacción reguladora. Fructosa 6-P ATP Pi PFK-2 FBpasa- ADP Fructosa 2,6 bifosfato Control por piruvato quinasa La reacción que cataliza la piruvato quinasa también sirve de punto de control, pero de menor importancia. En animales hay diferentes tipos de piruvato quinasa: L (liver) y M (muscle). El metabolito fructosa 1,6BP activa la piruvato quinasa , ya que la fructosa 1,6BP es el producto de la reacción reguladora, y su destino principal es transformarse en piruvato.
- En el hígado, la piruvato quinasa es regulada por la PK-A (la proteína-quinasa que regula la concentración de la F2,6P 2 ).
- En el músculo, la piruvato quinasa es controlada por otro método de control fosforilante. En una dieta rica en carbohidratos, al cabo de unos días aumenta la concentración de la piruvato quinasa, como consecuencia de la actividad de regulación de la expresión génica. Piruvato kinasa fosforilada Bajo nivel de glucosa en sangre Glucagón H 2 O ADP PKA AMPc Pi ATP + Desfosforilada Isozimas: 2 enzimas que catalizan la misma reacción pero que están codificadas por el mismo gen. + Isozimas de la PK
corrixir debuxo