¡Descarga capitulo 20 langma ojo.docx y más Apuntes en PDF de Microbiología solo en Docsity!
OJO
COPA ÓPTICA Y VESÍCULA DEL
CRISTALINO
En el embrion de 22 dias el ojo aparece como un par de surcos poco profundos a ambos lados del prosencéfalo (Fig. 20.1). Al cerrarse el tubo neural, los surcos forman evaginaciones en el prosencefalo: las vesiculas opticas. Mas tarde, estas entran en contacto con el ectodermo superficial produciendo en el los cambios necesarios para la formacion del cristalino (Fig. 20.1). Poco despues las vesiculas opticas empiezan a invaginarse para constituir la copa optica de pared doble (Figs. 20.1 y 20.2 A ). En un principio las capas interna y externa de la copa están separadas por una luz: el espacio intrarretiniano (Fig. 20.2 B ). Pero pronto desaparece la luz y las dos capas se unen (Fig. 20.2 D,E ). La invaginacion no se limita a la porcion central de la copa, sino que incluye una parte de la superficie inferior (Fig. 20.2 A ) que forma la fisura coroidea. Esto permite a la arteria hialoidea llegar hasta la camara interna del ojo (Figs. 20.3 y 20.7). Durante la septima semana los labios de la fisura coroidea se fusionan y la boca de la copa optica se convierte en una abertura redonda: la futura pupila.
Durante los procesos anteriores las celulas del ectodermo superficial, inicialmente en contacto con la vesicula optica, comienza a alargarse para crear la placoda del cristalino (Fig. 20.1). Luego esta se invagina convirtiendose en la vesicula del cristalino. En la quinta semana la vesicula del cristalino pierde contacto con el ectodermo superficial para situarse en la boca de la copa optica (Figs. 20.2 C,E y 20.3).
RETINA, IRIS Y CUERPO CILIAR
Se da el nombre de capa pigmentaria de la retina a la capa externa de la copa optica que se caracteriza por pequenos granulos pigmentarios (Figs. 20.2 D,E y 20.6). El desarrollo de la capa interna (neural) de la cupula optica resulta mas complejo. Las cuatro quintas partes, parte óptica de la retina, contienen celulas alrededor del espacio intrarretiniano (Fig. 20.3) que se diferencian en bastones y conos (Fig. 20.4). Los bastones son mas numerosos (120 millones) y sensibles que los conos (de 6 a 7 millones), pero sin que detecten el color como los conos. Junto a este capa fotorreceptora se halla la capa del manto que, igual que en el cerebro, genera neuronas y células de soporte, entre ellas la capa nuclear externa , la capa nuclear interna y la capa de células ganglionares (Fig. 20.4). Sobre la superficie se encuentra una capa
fibrosa que contiene los axones de las neuronas de las capas mas profundas. En esta zona, las fibras nerviosas convergen en el tallo optico, el cual se convertira en el nervio optico (Fig. 20.3). Por ello los impulsos luminosos cruzan la mayoria de las capas de la retina antes de llegar a los bastones y conos.
La quinta parte anterior de la capa interna –la parte ciega de la retina – conserva el grosor de una capa. Mas tarde se dividira en parte irídea de la retina que constituye la capa interna del iris y en parte ciliar de la retina que participa en la formación del cuerpo ciliar (Figs. 20.5 y 20.6).
Mientras tanto la region entre la cupula optica y el epitelio superficial que la recubre esta llena de mesenquima laxo (Figs. 20.2 C y 20.6). Los musculos esfínter y dilatador de la pupila se forman en este tejido (Fig. 20.5). Se desarrollan a partir del ectodermo subyacente de la copa optica. En el adulto el iris esta constituido por la capa externa sin pigmento de la copa optica y por una capa de tejido conectivo muy vascularizado que contiene los musculos de la pupila (Fig. 20.5).
La parte ciliar de la retina se identifica fácilmente por sus pliegues pronunciados. Afuera esta cubierta por una capa de mesenquima que da origen al músculo ciliar ; adentro se conecta al cristalino mediante una red de fibras elasticas: el ligamento suspensorio (Fig. 20.6). La contraccion del musculo ciliar modifica la tension del ligamento y controla la curvatura del cristalino.
CRISTALINO
Poco despues de formarse la vesicula del cristalino (Fig. 20.2 C ), las celulas de la pared posterior empiezan a alargarse anteriormente para producir fibras largas que poco a poco van llenando la luz de la vesicula (Fig. 20.3). Hacia el final de la septima semana esas fibras primarias del cristalino llegan a la pared anterior de la vesicula del cristalino. Sin embargo, el crecimiento del cristalino no termina en esta fase, porque otras fibras (secundarias) se agregan constantemente al nucleo central.
COROIDES, ESCLERÓTICA Y CÓRNEA
Al final de la quinta semana, el primordio del ojo esta totalmente rodeado por mesenquina laxo (Fig. 20.3). Este tejido pronto se diferencia en una capa interna semejante a la piamadre del encefalo y en una capa externa semejante a la duramadre. La capa interna produce una capa pigmentaria muy vascularizada conocida como coroides; la capa externa se convierte en la esclerotica y se continua con la duramadre alrededor del nervio optico (Fig. 20.6). Se diferencian de otro modo las capas mesenquimatosas
superficial que da origen al cristalino. Conforme prosigue el desarrollo, al parecer PAX6 no es indispensable para la formación de la copa optica. Por el contrario, este proceso esta controlado por las senales interactivas entre la vesicula optica, el mesenquima circundante y la superficie suprayacente en la region donde se origina el cristalino (Fig. 20.9). Asi pues, los factoresde crecimiento de fibroblastos (FGF) procedentes del ectodermo superficial facilitan la diferenciación de la retina neural (capa interna). En cambio, el factor de transformacion del crecimiento β (TGF-β), secretado por el mesenquima circundante, dirige la formacion de la capa retinal pigmentaria (externa). En direccion 3’ con estos productos genicos los factores de transcripcion MITF y CHX10 se expresan y dirigen la diferenciacion de la capa pigmentaria y neural, respectivamente (Fig. 20.9). De ahi que el ectodermo del cristalino sea esencial para que se forme bien la copa optica, de modo que esta no se invaginaria sin una placoda del cristalino.
La diferenciacion del cristalino depende del gen PAX6 , aunque no se encarga de la actividad inductora de la vesicula optica. Por el contrario, actua en el ectodermo superficial para regular el desarrollo del cristalino (Fig. 20.9 C ). Esta expresion regula al alza el factor de transcripcion SOX2 , manteniendo tambien la expresion de PAX6 en el futuro ectodermo del cristalino. Por su parte, la vesicula óptica segrega la proteina BMP4 que tambien regula al alza y mantiene la expresion de SOX2, asi como la de LMAF, otro factor de transcripcion (Fig. 20.9 C ).
Despues PAX6 controla la expresion de dos genes de homeodominio: SIX3 y PROX1. La expresion combinada de PAX6, SOX2 y LMAF inicia la expresión de los genes encargados de formar el cristalino, entre ellos PROX. SIX3 tambien actua como regulador en la produccion de cristalino inhibiendo el gen respectivo. Por ultimo PAX controla la proliferación celular en el cristalino mediante FOX3.
CONSIDERACIONES CLÍNICAS
Anomalías oculares
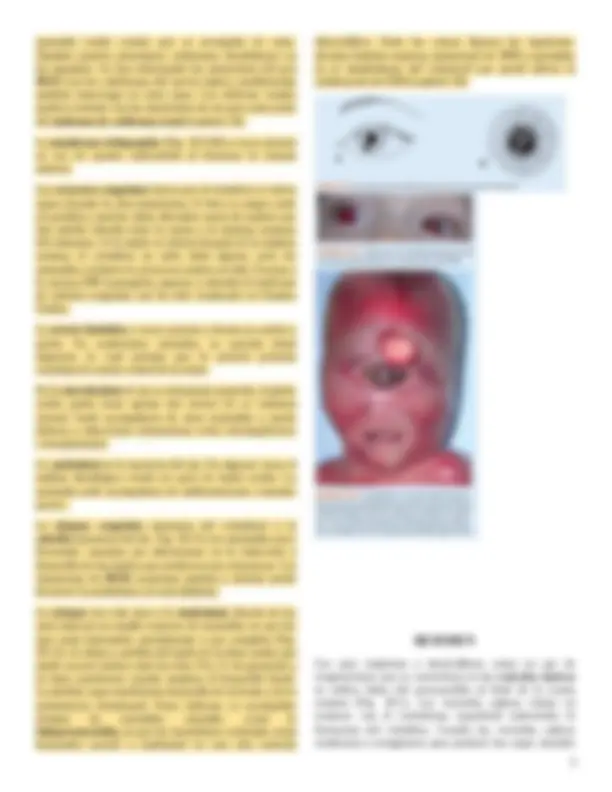
El coloboma puede ocurrir si la fisura coroidea no cierra. En condiciones normales lo hace durante la séptima semana del desarrollo, (Fig. 20.7). De no ser así, persiste una hendidura. Aunque éste casi siempre se localiza en el iris exclusivamente, coloboma del iris (Fig. 20.10 A ), puede extenderse hasta el interior del cuerpo ciliar, la retina, la coroides y el nervio óptico. El coloboma es una
anomalía ocular común que se acompaña de otras. También pueden presentarse colobomas (hendiduras) en los párpados. Se han relacionado las mutaciones del gen PAX2 con los colobomas del nervio óptico; posiblemente también intervenga en otros tipos. Los defectos renales pueden coexistir con las mutaciones de ese gen como parte del síndrome de coloboma renal (capítulo 16).
La membrana iridopupilar (Fig. 20.10 B ) a veces persiste en vez de quedar reabsorbida al formarse la cámara anterior.
Las cataratas congénitas hacen que el cristalino se vuelva opaco durante la vida intrauterina. Si bien su origen suele ser genético, muchos niños afectados nacen de madres que han sufrido rubeola entre la cuarta y la séptima semanas del embarazo. Si la madre se infecta después de la séptima semana, el cristalino no sufre daño alguno, pero las anomalías cocleares le provocan sordera al niño. Gracias a la vacuna SPR (sarampión, paperas y rubeola) el síndrome de rubéola congénita casi ha sido erradicado en Estados Unidos.
La arteria hialoidea a veces persiste y forma un cordón o quiste. En condiciones normales, su porción distal degenera, lo cual permite que la porción próxima constituya la arteria central de la retina.
En la microftalmía el ojo es demasiado pequeño; el globo ocular puede tener apenas dos tercios de su volumen normal. Suele acompañarse de otras anomalías y puede deberse a infecciones intrauterinas como citomegalovirus o toxoplasmosis.
La anoftalmía es la ausencia del ojo. En algunos casos el análisis histológico revela un poco de tejido ocular. La anomalía suele acompañarse de malformaciones craneales graves.
La afaquia congénita (ausencia del cristalino) y la aniridia (ausencia del iris. Fig. 20.11) son anomalías poco frecuentes causadas por alteraciones en la inducción y desarrollo de los tejidos que producen esas estructuras. Las mutaciones de PAX6 ocasionan aniridia y además puede favorecer la anoftalmía y la microftalmía.
La ciclopía (un solo ojo) y la sinoftalmía (fusión de los ojos) abarcan un amplio espectro de anomalías en que los ojos están fusionados parcialmente o por completo (Fig. 20.12). Se deben a pérdida del tejido de la línea media que puede ocurrir incluso entre los días 19 y 21 de gestación o en fases posteriores cuando empieza el desarrollo facial. La pérdida causa insuficiente desarrollo de la frente y de la prominencia frontonasal. Estos defectos se acompañan siempre de anomalías craneales como la holoprosencefalia, en que los hemisferios cerebrales están fusionados parcial o totalmente en una sola vesícula
telencefálica. Entre las causas figuran las siguientes: alcohol, diabetes materna, mutaciones de SHH y anomalías en el metabolismo del colesterol que puede alterar la señalización de SHH (capítulo 18).
RESUMEN
Los ojos empiezan a desarrollarse como un par de evaginaciones que se convertiran en las vesículas ópticas en ambos lados del prosencefalo al final de la cuarta semana (Fig. 20.1). Las vesiculas opticas entran en contacto con el ecdodermo superficial induciendo la formacion del cristalino. Cuando las vesiculas opticas comienzan a invaginarse para producir las capas neurales