¡Descarga CASOS INTEGRADORES 2 y más Apuntes en PDF de Anatomía Patológica solo en Docsity!
FACULTAD DE FARMACIA
UNIVERSIDAD COMPLUTENSE
TRABAJO FIN DE GRADO
Fiebre tifoidea y factores de virulencia de
Salmonella enterica serotipo Typhi
Autor: Elena Marín Rodríguez
Tutor: Concepción Pintado García
Convocatoria: Junio 2018
Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
Resumen Salmonella Typhi, la bacteria causante de fiebre tifoidea, infecta las células M del intestino a través de los sistemas de secreción de Tipo III codificados en las islas de patogenicidad 1 y 2. Después, alcanza las placas de Peyer donde infecta células fagocíticas que le van a permitir diseminarse por el organismo. Mediante dichos sistemas de secreción introduce una serie de factores que causan cambios en la célula hospedadora permitiéndole vivir de forma intracelular en ella. S. Typhi no solo hace uso de proteínas efectoras sino que, además, tiene la capacidad de producir la toxina tifoidea la cual procede de dos toxinas, CDT y la toxina pertussis, con una estructura novedosa que la hace característica de esta bacteria. La presencia de la toxina podría estar relacionada con una modulación de la respuesta inmune en el individuo lo que concuerda con el hecho de que haya una gran cantidad de portadores crónicos asintomáticos. Este estado de portador se debe a la formación de biofilms de S. Typhi en cálculos biliares de la vesícula biliar y podría corresponderse con una mayor predisposición a tener cáncer en dicho órgano. Abstract Salmonella Typhi, the typhoid fever causing agent, infects M cells from the gut through type III secretion systems codified by the pathogenic island 1 and 2. Subsequently, it reaches Peyer’s Patches where it infects phagocytic cells which allow bacteria to disseminate across the organism. Through these secretion systems, it introduces a range of factors that cause changes in host cells, allowing bacteria to live intracellularly. S. Typhi not only make use of effector proteins but also, has the capacity of developing typhoid toxin which comes from CDT and pertussis toxin, with a novel structure that makes it characteristic of this bacterium. Presence of the toxin could be associated to a modulation of the immune system response in the individual which agrees with the fact that there is a large number of asymptomatic chronic carriers. This carrier state is a consequence of S. Typhi biofilms in gallstones from the gallbladder and could correspond to a greater predisposition of developing gallbladder cancer. Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
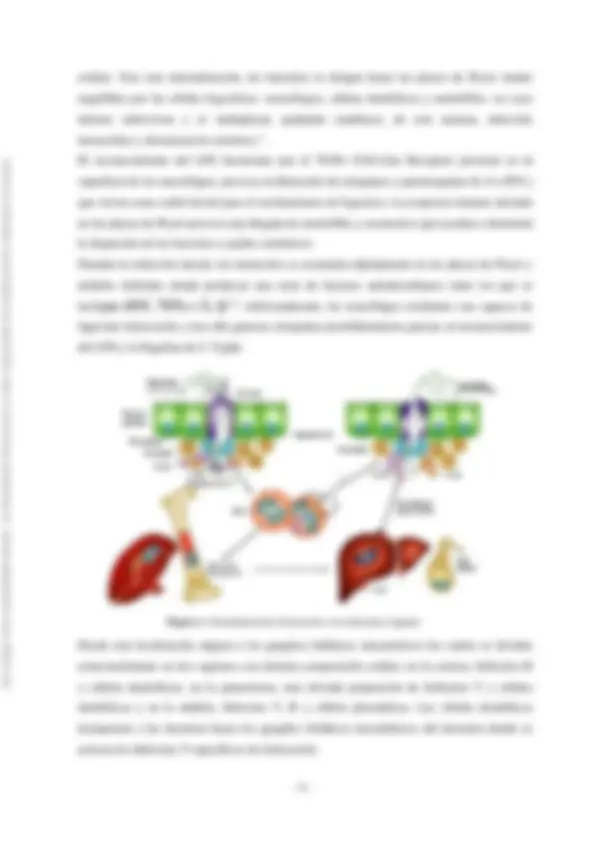
celular. Tras esta internalización, las bacterias se dirigen hacia las placas de Peyer siendo engullidas por las células fagocíticas - macrófagos, células dendríticas y neutrófilos- en cuyo interior sobreviven y se multiplican, pudiendo establecer, de esta manera, infección intracelular y diseminación sistémica^7. El reconocimiento del LPS bacteriano por el TLR4 (Toll-Like Receptor) presente en la superficie de los macrófagos, provoca la liberación de citoquinas y quimioquinas IL-6 e IFN-γ que sirven como señal inicial para el reclutamiento de fagocitos. La respuesta inmune iniciada en las placas de Peyer provoca una llegada de neutrófilos y monocitos que ayudan a disminuir la dispersión de las bacterias a tejidos sistémicos. Durante la infección inicial, los monocitos se acumulan rápidamente en las placas de Peyer y nódulos linfoides donde producen una serie de factores antimicrobianos entre los que se incluyen iNOS, TNFα e IL-1β^8. Adicionalmente, los macrófagos residentes son capaces de fagocitar Salmonella y tras ello generar citoquinas proinflamatorias gracias al reconocimiento del LPS y la flagelina de S. Typhi. Figura 1. Diseminación de Salmonella a los diferentes órganos Desde esta localización migran a los ganglios linfáticos mesentéricos los cuales se dividen estructuralmente en tres regiones con distinta composición celular; en la corteza, linfocitos B y células dendríticas; en la paracorteza, una elevada proporción de linfocitos T y células dendríticas y en la médula, linfocitos T, B y células plasmáticas. Las células dendríticas transportan a las bacterias hasta los ganglios linfáticos mesentéricos del intestino donde se activan los linfocitos T específicos de Salmonella. Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
A continuación, las bacterias pasan a sangre desde el sistema linfático a través del conducto torácico produciendo una bacteriemia primaria transitoria siendo de nuevo captadas por los macrófagos que recubren los sinusoides del hígado, bazo, médula ósea y vesícula biliar, donde las bacterias se siguen multiplicando. Desde estos órganos regresan a la sangre, causando una bacteriemia secundaria comenzando en ese momento las manifestaciones clínicas (incremento de la temperatura corporal, síntomas pseudogripales e incluso problemas neurológicos). Las bacterias vuelven a ser eliminadas de la sangre por los macrófagos y vía hígado, alcanzan la vesícula biliar donde son capaces de establecer biofilms si se dan las condiciones adecuadas, persistiendo adheridas a cálculos biliares; esto permite la reinfección del tracto intestinal estableciéndose en las placas de Peyer del íleon distal donde causan inflamación, ulceración y necrosis. Las bacterias se excretan en heces y unas tres semanas después pueden aparecer hemorragias en la zona ulcerada e incluso perforación, la cual daría lugar a una septicemia que conlleva la muerte por fiebre tifoidea en la mayoría de los casos^1. Aproximadamente un 90% del genoma de S. Typhi está relacionado con secuencias que se presentan en otros serovares de Salmonella , lo que significa que hay un genoma central de Salmonella que facilita la colonización, infección y transmisión. El genoma específico de S. Typhi presenta unos 300-400 genes que están asociados a fagos específicos o a Islas de Patogenicidad de Salmonella (SPIs); hasta ahora se han descubierto 15 SPIs en S. Typhi, dos de las cuales (SPI-1 y SPI-2) codifican sistemas de secreción de proteínas muy importantes en la patogenia. Estas SPIs se encuentran flanqueadas por secuencias repetidas y tienden a poseer una composición variada de G/C comparada con las regiones a su alrededor^9. S. Typhi produce un repertorio de productos que contribuyen a la patogénesis, algunos son compartidos por el resto de serovares o serotipos pero otros son únicos como es el caso de la toxina tifoidea , la cual es altamente inmunogénica y su expresión se encuentra altamente regulada cuando S. Typhi infecta células humanas^4. La supervivencia de S. Typhi en el interior de los macrófagos se debe fundamentalmente a una resistencia al estrés oxidativo, por ejemplo mediante la reducción de intermediarios reactivos de nitrógeno, la actuación de la arginasa así como enzimas que degradan peróxidos (3 catalasas y 2 alquil hidroperóxido reductasas)^6. El resultado de la infección de Salmonella Typhi viene determinado por factores relacionados tanto con el hospedador como con la bacteria. Esto incluye la virulencia de S. Typhi, la habilidad del hospedador para ejercer una respuesta inmune adecuada y finalmente con su capacidad para destruir el patógeno^7. Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
La estructura de la base del complejo T3SS1, constituida por varios anillos conectados entre sí, se extiende desde la membrana interna hasta la membrana externa de Salmonella , y de ella sale una estructura en forma de aguja que interacciona con la célula hospedadora. En el interior de la base y de la aguja hay una varilla que forma el conducto entre el citoplasma bacteriano y la membrana plasmática de la célula hospedadora^13. En la región citoplasmática de T3SS1 hay un complejo de ATPasa que facilita el transporte de efectores moleculares a través del conducto hacia una estructura de translocasa en la membrana de la célula hospedadora^9 (Fig. 2). SPI-1 por tanto codifica una serie de proteínas efectoras que se translocan a través del T3SS1 y Figura 2. El T3SS está formado por un anillo (A), una maquinaria exportadora citoplásmica (B) y una estructura de anillo externa (C). que desempeñan diferentes funciones en la invasión de S. Typhi. Dichas funciones se resumen en la Tabla 1. Proteína efectora Efecto en la célula hospedadora SipA Reclutamiento de neutrófilos y reorganización del citoesqueleto SipB Nucleación de actina y translocación de otros efectores SipC Translocación de otros efectores SopA Reclutamiento de células inmunes y secreción de fluidos SopB Reorganización del citoesqueleto, reclutamiento de neutrófilos y secreción de fluidos SopC Reclutamiento de neutrófilos y secreción de fluidos SopD Reclutamiento de neutrófilos y secreción de fluidos SopE Reorganización del citoesqueleto SptP Reorganización del citoesqueleto Tabla 1. Proteínas efectoras del sistema de secreción de tipo III de la Isla de Patogenicidad- 1 Los efectores SipA y SipC ( Salmonella invasion proteins) afectan a la actina directamente durante los procesos de invasión, pero lo hacen a diferentes niveles. SipA puede unirse y estabilizar a la actina. SipC, que forma parte, junto con SipB, del poro de translocación de T3SS1, puede causar independientemente, el reordenamiento de la actina a través de distintos Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
dominios que desembocan en el ruffling de la membrana 14, 15 ,^^16 , es decir, una reorganización de la membrana plasmática y del citosol de la célula hospedadora con la consiguiente formación de extrusiones o pseudópodos que rodean e internalizan a la bacteria. Además de modular la actina, SipC interacciona directamente con Exo70 , un componente del complejo del exocisto, que interviene en el acoplamiento y fusión de las vesículas exocíticas con la membrana plasmática^17. Salmonella también altera el citoesqueleto de actina a través de la manipulación de fosfoinósidos. La membrana plasmática está íntimamente asociada con el citoesqueleto de actina y dicha interacción es a través del fosfatidil inositol 4-5 bifosfato (PtdIns (4-5) P2)^9. SopB/SigD ( Salmonella outer protein) es una inositol-fosfatasa producida por SPI-1 que provoca la desaparición del PtdIns (4-5) P2 de regiones invaginadas de la membrana durante la invasión de Salmonella^18 ; esto permite que la membrana plasmática sea más elástica y así se puede facilitar el remodelamiento para la entrada de S. Typhi. Los efectores SopE y SopE2 actúan en concreto como factores intercambiadores de guanina (GEFs) para las pequeñas GTPasas Cdc42 y Rac 19. SopE, además, puede también activar RalA , una GTPasa necesaria para la unión del exocisto^17. Tras la invasión, SptP, un efector implicado en la mediación de la recuperación del citoesqueleto del huésped después de la infección, actúa como una proteína activadora de GTPasa (GAP) para Cdc42 y Rac1 , inactivando dichas proteínas G y devolviendo la morfología celular a un estado normal relativo^20 (Fig. 3). Figura 3. Entrada de Salmonella en la célula hospedadora La principal proteína reguladora de T3SS1 es HilA , cuya expresión esta mediada por factores ambientales importantes para la supervivencia celular^14. Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
Figura 4. Estructura toxina tifoidea sus características virulentas y esto podría relacionarse con la pérdida de selectividad en sus hospedadores en estos serovares. Toxina tifoidea Salmonella Typhi posee un potente factor de patogenicidad que es la toxina tifoidea, la cual se ha descubierto recientemente. Dicha toxina parece haber surgido tras la combinación evolutiva de las actividades de dos exotoxinas antecesoras, como son CDT ( Cytolethal Distending Toxin, un tipo de toxinas heterotriméricas producidas por bacterias Gram negativas con actividad DNAsa) y la toxina pertussis^30. Se trata de una toxina de tipo AB con una estructura única A 2 B 5 sin precedentes. A diferencia de otras toxinas de tipo AB, como la toxina Shiga, la toxina colérica o la toxina pertussis en las que solo existe una subunidad A, la toxina tifoidea está constituida por dos subunidades A, CdtB y PltA, unidas covalentemente entre sí y asociadas no covalentemente a la subunidad B pentamérica PltB^31 (Fig.4). La existencia de un único residuo de cisteína en las dos subunidades A permite la formación de un puente disulfuro que las mantiene unidas covalentemente, no habiendo ningún otro tipo de interacción entre ambas. La subunidad PltA se asocia con el pentámero PltB a través de una hélice corta en el carboxilo terminal que se inserta en el lumen hidrofóbico del canal PltB. No existen interacciones entre CdtB y PltB. La subunidad CdtB se ancla al complejo PltA/PltB a través del puente disulfuro entre ambas subunidades A, siendo necesario ese puente para el ensamblaje de la toxina tifoidea en el periplasma de la bacteria. PltA es una ADP ribosil-transferasa, que posee una secuencia aminoacídica así como semejanzas estructurales a las de la toxina pertussis S1^31 , mientras que CdtB tiene una actividad desoxirribonucleasa que detiene el ciclo celular o causa muerte celular debido al daño que causa en el ADN de la célula intoxicada 32, 33. La subunidad PltB de la toxina tifoidea se une a un receptor que contiene una secuencia consenso de un sialoglicano específico. Se trata del ácido N-acetilneuramínico (Neu5Ac)- α2,3Galactosa (Gal)-β1,3/β1,4Glucosa/N-acetilglucosamina (GlcNAc), muy común entre las glicoproteínas superficiales y que se encuentra abundantemente en humanos^34. De esta secuencia trisacarídica, el azúcar terminal ácido N-acetilneuramínico es el más importante en Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
la unión ya que va a encajar en un bolsillo de PltB gracias a una serina. La toxina tifoidea está codificada en una isla de patogenicidad y se produce exclusivamente por la bacteria cuando se encuentra en células infectadas. Una vez sintetizada es excretada en el interior del lumen de la VCS por TtsA (typhoid toxin secretion protein A) la cual pertenece a una clase de endolisinas de bacteriófagos. En concreto, TtsA se trata de una N-acetil-β-D- muraminidasa. Tras su secreción en el lumen de la VCS, la toxina es empaquetada en transportadores vesiculares y transportada al espacio extracelular donde puede alcanzar células diana. Es importante destacar que la toxina no necesariamente afecta las células que contienen las bacterias, sino que puede intoxicar a cualquier célula localizada en un lugar remoto, tenga o no en su interior VCS, como neutrófilos, células endoteliales del cerebro, linfocitos, etc. 31, 33. La toxina tifoidea muestra una gran selectividad por los azúcares terminados en Neu5Ac (ácido N-acetilneuramínico), los cuales son muy comunes en las células humanas, al contrario que Neu5Gc (ácido N-glicolilneuramínico) que es más propio de otros mamíferos como los chimpancés, esta sería una de las razones esenciales por la especificidad de S. Typhi por los seres humanos. La presencia de este azúcar inusual en humanos se debe a la mutación del gen que codifica la enzima CMP-ácido N-acetilneuramínico hidroxilasa, CMAH , la cual está presente en otros mamíferos y convierte Neu5Ac en Neu5Gc^35 (Fig. 5); se cree que dicha mutación podría haber surgido después de que los homínidos se separasen de otros primates. Esta es la razón por la cual S. Typhi no causa fiebre tifoidea en chimpancés (ya que su toxina no tiene un receptor al que unirse), y es un serotipo restringido a los seres humanos. En el caso de los ratones, éstos presentan una CMAH funcional y su expresión es variable ya que tienen los dos tipos de azúcares terminales, por lo que la toxina tifoidea es capaz de inducir los síntomas de la fiebre tifoidea en ellos. Sin embargo, si se expresa constitutivamente la enzima en los ratones para que sólo expresen Neu5Gc, se ha demostrado que son totalmente resistentes a la toxina tifoidea^36. Figura 5. Conversión de azúcares por CMAH Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
Esto, sin embargo, no ocurre en células y tejidos de ratones deficientes en Rab32 o en su factor intercambiador de nucleótido BLOC- 3 40. Rab32 define, por lo tanto, un nuevo modelo de mecanismo de defensa basado en la entrega de un factor antimicrobiano a la VCS de S. Typhi^30. Además Rab32 es escindida específicamente por la actividad proteolítica de GtgE , una proteína efectora del T3SS1 del SPI-1, producida por S. Typhimurium pero no por S. Typhi. GtgE actúa junto con una segunda proteína efectora, SopD2 que, aunque no se trate de una proteasa, inactiva Rab32 funcionando como GAP (proteína que acelera el intercambio de GTP→GDP) para su GTPasa^37. Como se ha mencionado anteriormente, la presencia de la toxina en dosis bajas podría intervenir en una infección persistente lo que estaría directamente relacionado con un estado de portador crónico. En regiones donde S. Typhi es endémica, aproximadamente un 1-4% de los individuos infectados pasan a ser portadores crónicos asintomáticos los cuales representan un peligro para la salud local pública^41 ,^42. Dicha infección crónica normalmente se localiza en la vesícula biliar, lo que se relaciona con una excreción de la bacteria a largo plazo. Además, aproximadamente un 90% de esos portadores crónicos posee también cálculos biliares y esta asociación se relaciona con una mayor predisposición para desarrollar un cáncer de vesícula biliar^43 -^47. Se trata del sexto cáncer más común del tracto gastrointestinal y representa uno de los más malignos debido a que se extiende por todo el tracto biliar. Su malignidad ha sido asociada con factores genéticos y de estilo de vida, pero la infección por S. Typhi y la producción de cálculos biliares son los principales factores de riesgo. Recientemente, la técnica de PCR anidada reveló la presencia del gen de la flagelina de S. Typhi en un 67.3% de muestras hepatobiliares de pacientes con cáncer de vesícula mientras que, en pacientes con enfermedades benignas de la vesícula y población sana, este porcentaje era significantemente más bajo^45 , por lo que la detección temprana de S. Typhi podría usarse como estrategia de prevención de cáncer de vesícula. La producción de un biofilm podría ejercer un papel clave en la colonización y persistencia crónica de S. Typhi^48. Existen informes que documentan que la bilis, rica en lípidos y con actividad antimicrobiana, induce la producción de una matriz formada por un exopolisacárido de antígeno O que facilita la formación del biofilms de S. Typhi sobre la superficie de los cálculos biliares humanos De ser así, podría darse un ambiente propicio para la persistencia de la bacteria y que ésta pudiera ser liberada al intestino donde, a través de las heces, reinfectaría a nuevos Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
hospedadores; además el epitelio de la vesícula estaría en continua exposición a los factores de patogenicidad de la bacteria (propiedades carcinogénicas potenciales). Actividad carcinogénica potencial de productos de S****. Typhi Diferentes productos bacterianos poseen potencial carcinogénico entre los que se incluyen la glucuronidasa, que después de actuar sobre la bilis origina unos intermediarios de alta energía, y los compuestos nitrosos, generados a partir del nitrato por acción de enzimas bacterianas^49 , pero esencialmente la subunidad CdtB de la toxina tifoidea induce un daño en el DNA y provoca la detención del ciclo celular, así como apoptosis; en concreto, dicha subunidad es un homólogo de la DNAsa, una endonucleasa que causa la ruptura de la doble hebra de DNA. La toxina entra en la célula a través de endocitosis y es transportada al aparato de Golgi que la guía hacia el retículo endoplásmico; a continuación, la subunidad CdtB se transloca al núcleo celular para provocar el daño en el DNA^50. Este hecho activa una cascada de acontecimientos como es el reclutamiento del complejo sensor al daño de DNA llamado MRN que inicia la resección del extremo del DNA produciendo un extremo 3’ y además, la acumulación de la quinasa ATM en el sitio del daño^51. Esta quinasa promueve la fosforilación de la histona H2AX y la activación de una serie de puntos de control de daño del DNA entre los que se incluyen el supresor de tumores p53 y su efector p21 , que paran el ciclo celular en G1^52. Por otra parte, ATM activa el punto de control 2 (CHK2) que inactiva la fosfatasa de ciclo de división celular (CDC25) y cuando dicha fosfatasa no funciona, se acumula la forma hiperfosforilada de una ciclina (CDK1) que bloquea la proliferación celular en la fase G2/M del ciclo celular^53 (Fig. 7). Figura 7. Entrada de la toxina y daño en el núcleo celular Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
incubados con la bacteria en presencia de bilis. La bilis ejerce un efecto intenso en la regulación directa e indirecta de genes en S. Typhi mediante la regulación a la baja de la expresión de genes que se ven involucrados en la invasión celular, en la movilidad y proteínas de membrana externas^58. Una vez que se ha formado el biofilm, las células individuales que lo componen presentan una tolerancia mayor a agentes antimicrobianos y por tanto el tratamiento antibiótico pasa a ser inadecuado^48. En la matriz del biofilm, las células microbianas muestran una concentración mínima inhibitoria (CMI) entre 10 y 1.000 veces mayor comparadas con las mismas células en condiciones de crecimiento planctónico 59, 60. Por lo tanto, la concentración efectiva de antibióticos que habría que alcanzar para eliminar el biofilm en el organismo es imposible de conseguir debido a su toxicidad y a los efectos adversos en los pacientes. La rápida identificación de cepas de S. Typhi formadoras de biofilms permitiría reconocer pacientes con alto riesgo, y eso mejoraría la prevención de cáncer relacionado con infecciones bacterianas^48. Conclusiones ▪ Los factores de patogenicidad de Salmonella Typhi vienen determinados por las Islas de patogenicidad, entre las que destacan SPI- 1 (con genes de invasión esenciales para la infección intestinal) y SPI- 2 (necesaria para la supervivencia intracelular de la bacteria). SPI-1 y SPI-2 codifican dos Sistemas de secreción de Tipo III, que facilitan el transporte de efectores hacia la célula hospedadora. ▪ La toxina tifoidea, uno de los principales factores de virulencia, se forma exclusivamente cuando la bacteria es intracelular y presenta una arquitectura A 2 B 5 única. Uno de sus principales componentes, CdtB , es capaz de provocar un daño en el DNA y detener el ciclo celular, así como causar apoptosis. La toxina tiene como receptor una secuencia trisacarídica cuyo azúcar terminal solo se encuentra en humanos debido a la desaparición de la enzima CMAH. Esta enzima está presente en otros mamíferos y, por ello, S. Typhi no causa fiebre tifoidea en ellos. ▪ S. Typhi es capaz de formar biofilms en la superficie de cálculos de la vesícula biliar, hecho que podría suponer un factor clave para el desarrollo de la infección persistente en dicha localización y a la existencia de portadores crónicos asintomáticos, que excretan la bacteria y que diseminan la enfermedad. Esta situación aumenta la predisposición de desarrollar cáncer de vesícula biliar ya que la persistente liberación de mediadores de inflamación, toxinas y metabolitos, pueden ser potencialmente mutagénicos. Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
Bibliografía
1. (^) Martínez Álvarez N. Virulencia, resistencia y elementos genéticos móviles en serotipos no prevalentes de Salmonella enterica. Memoria Grado de Doctor. 2007. 2. (^) Karkey A, Thompson CN, Tran Vu Thieu N, Dongol S, Le Thi Phuong T, et al. Differential epidemiology of Salmonella Typhi and Paratyphi A in Kathmandu, Nepal: a matched case control investigation in a highly endemic enteric fever setting. PLoS Negl. Trop. Dis. 2013; 7(8):e 3. (^) Pulickal AS, Gautam S, Clutterbuck EA, Thorson S, Basynat B, et al. Kinetics of the natural, humoral immune response to Salmonella enterica serovar Typhi in Kathmandu, Nepal. Clin. Vaccine Immunol. 2009. 16:1413– 1419 4. (^) Dougan G, Baker S. Salmonella enterica Serovar Typhi and the Pathogenesis of Typhoid Fever. Annu. Rev. Microbiol. 2014; 68: 317- 3 36. 5. WHO.int [Internet]. [actualizado Ene 2018; citado 5 abr 2018]. Disponible en: http://www.who.int/features/qa/typhoid-fever/es/ 6. (^) Kurtz JR, Goggins JA, McLachaln JB. Salmonella infection: Interplay between the bacteria and host immune system. Immunology Letters. 2017 (190): 42- 50 7. (^) Lahiri A, Lahiri A, Iyer N, Das P, Chakravortty D. Visiting the cell biology of Salmonella infection. Microbes and Infection. 2010; 12: 809- 818 8. (^) Kaiser P, Hardt WD. Salmonella Typhimurium diarrhea: switching the mucosal epithelium from homeostasis to defense, Curr. Opin. Immunol. 2011; 23: 456– 463 9. (^) Kaur J, Jain S. K. Role of antigens and virulence factors of Salmonella enterica serovar Typhi in its pathogenesis. Microbiological research. 2012; 167: 199- 210 10. (^) Hansen-Wester I, Hensel M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001; 3: 549– 559 11. (^) Hueck CJ. Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol Mol Biol Rev. 1998; 62: 379– 433 12. (^) Marlovits TC, Stebbins CE. Type III secretion systems shape up as they ship out. Curr Opin Microbiol. 2010; 13: 47– 52 13. (^) Galán JE, Wolf-Watz H. Protein delivery into eukaryotic cells by type III secretion machines. Nature. 2006; 444: 567– 573 14. (^) Lostroh CP, Lee CA. The Salmonella pathogenicity island-1 type III secretion system. Microbes Infect. 2001; 3: 1281– 1291 15. McGhie EJ, Hayward RD, Koronakis V. Cooperation between actin-binding proteins of invasive Salmonella : SipA potentiates SipC nucleation and bundling of actin. EMBO J. 2001; 20: 2131– 2139 16. (^) Myeni SK, Zhou D. The C terminus of SipC binds and bundles F-actin to promote Salmonella invasion. J Biol Chem. 2010; 285: 13357– 13363 17. (^) Nichols CD, Casanova JE. Salmonella - directed recruitment of new membrane to invasion foci via the host exocyst complex. Curr Biol. 2010; 20: 1316– 1320 Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
35. (^) Chou HH, Hayakawa T, Diaz S, Krings M, Indriati E, Leakey M, et al. Inactivation of CMP-N- acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution. Proc Natl Acad Sci U S A. 2002;99(18):11736- 11741 36. (^) Deng L, et al. Host adaptation of a bacterial toxin from the human pathogen Salmonella Typhi. Cell. 2014; 159(6):1290– 1299 37. (^) Song J, Willinger T, Rongvaux A, Eynon EE, Stevens S, Manz MG, et al. A mouse model for the human pathogen Salmonella Typhi. Cell Host Microbe. 2010;8(4):369- 3 76. 38. (^) Spanò S, Liu X, Galán JE. Proteolytic targeting of Rab29 by an effector protein distinguishes the intracellular compartments of human-adapted and broad-host Salmonella. Proc Natl Acad Sci USA. 2011, 108(45):18418– 18423 39. (^) Spanò S, Galán JE. A Rab32-dependent pathway contributes to Salmonella typhi host restriction. Science. 2012, 338(6109):960– 963 40. (^) Spanò S, Gao X, Hannemann S, Lara-Tejero M, Galán JE. A bacterial pathogen targets a host Rab- family GTPase defense pathway with a GAP. Cell Host Microbe. 2016, 19(2):216– 226 41. (^) Crump JA, Mintz ED. Global trends in typhoid and paratyphoid Fever. Clin. Infect. Dis. 2010, 50, 241 – 246. 42. (^) Mogasale V, Maskery B, Ochiai RL, Lee JS, Mogasale VV, Ramani E et al. Burden of typhoid fever in low-income and middle-income countries: A systematic, literature-based update with risk-factor adjustment. Lancet Glob. Health 2014, 2, e570–e 43. (^) Caygill CP, Hill MJ, Braddick M, Sharp JC. Cancer mortality in chronic typhoid and paratyphoid carriers. Lancet 1994, 343, 83–84. 44. (^) Dutta U, Garg PK, Kumar R, Tandon RK. Typhoid carriers among patients with gallstones are at increased risk for carcinoma of the gallbladder. Am. J. Gastroenterol. 2000, 95, 784–787. 45. (^) Nath G, Singh YK, Kumar K, Gulati AK, Shukla VK, Khanna AK et al. Association of carcinoma of the gallbladder with typhoid carriage in a typhoid endemic area using nested PCR. J. Infect. Dev. Ctries. 2008, 2, 302–307. 46. (^) González-Escobedo G, Marshall JM, Gunn JS. Chronic and acute infection of the gall bladder by Salmonella Typhi: Understanding the carrier state. Nat. Rev. Microbiol. 2011, 9, 9–14. 47. Gunn JS, Marshall JM, Baker S, Dongol S, Charles RC, Ryan ET. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014, 22, 648– 655. 48. (^) Di Domenico EG, Cavallo I, Pontone M, Toma L, Ensoli F. Biofilm Producing Salmonella Typhi: Chronic Colonization and Development of Gallbladder Cancer. Int. J. Mol. Sci. 2017; 18(1887): 1- 14 49. (^) Nath G, Gulati AK, Shukla VK. Role of bacteria in carcinogenesis, with special reference to carcinoma of the gallbladder. World J Gastroenterol 2010 November 21; 16(43): 5395- 5404 50. (^) Miller R., Wiedmann M. Dynamic Duo-The Salmonella Cytolethal Distending Toxin Combines ADP- Ribosyltransferase and Nuclease Activities in a Novel Form of the Cytolethal Distending Toxin. Toxins 2016, 8, 121 Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.
51. (^) Di Domenico EG, Romano E, del Porto P, Ascenzioni F. Multifunctional role of ATM/Tel1 kinase in genome stability: From the DNA damage response to telomere maintenance. BioMed Res. Int. 2014, 2014, 787404 52. (^) Grasso F, Frisan T. Bacterial Genotoxins: Merging the DNA Damage Response into Infection Biology. Biomolecules. 2015, 5: 1762– 1782 53. (^) Li L, Sharipo A, Chaves-Olarte E, Masucci MG, Levitsky V, Thelestam M et al. The Haemophilus ducreyi cytolethal distending toxin activates sensors of DNA damage and repair complexes in proliferating and non-proliferating cells. Cell Microbiol. 2002, 4, 87– 99 54. (^) Guerra L, Carr HS, Richter-Dahlfors A, Masucci MG, Thelestam M, Frost JA et al. A bacterial cytotoxin identifies the RhoA exchange factor Net1 as a key effector in the response to DNA damage. PLoS ONE 2008, 3, e 55. (^) Dongol S, Thompson CN, Clare S, Nga TV, Duy PT, Karkey A et al. The microbiological and clinical characteristics of invasive Salmonella in gallbladders from cholecystectomy patients in Kathmandu, Nepal. PLoS ONE 2012, 7, e 56. (^) Kalai Chelvam K, Chai LC, Thong KL. Variations in motility and biofilm formation of Salmonella enterica serovar Typhi. Gut. Pathog. 2014, 6, 2. 57. (^) Crawford RW, Rosales Reyes R, Ramirez-Aguilar Mde L, Chapa-Azuela O, Alpuche-Aranda C, Gunn JS. Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc. Natl. Acad. Sci. USA 2010, 107, 4353– 4358 58. (^) Crawford RW, Gibson DL, Kay WW, Gunn JS. Identification of a bile-induced exopolysaccharide required for Salmonella biofilm formation on gallstone surfaces. Infect. Immun. 2008, 76, 5341–5349. 59. (^) Høiby N, Ciofu O, Johansen HK, Song ZJ, Moser C, Jensen PØ et al. The clinical impact of bacterial biofilms. Int. J. Oral Sci. 2011, 3, 55–65. 60. (^) Hengzhuang W, Wu H, Ciofu O, Song Z, Høiby N. In vivo pharmacokinetics/pharmacodynamics of colistin and imipenem in Pseudomonas aeruginosa biofilm infection. Antimicrob. Agents Chemother. 2012, 56, 2683 – 2690 Este trabajo tiene una finalidad docente. La Facultad de Farmacia no se hace responsable de la información contenida en el mismo.