¡Descarga CÉLULAS DEL CONJUNTIVO y más Apuntes en PDF de Biología Celular solo en Docsity!
Tejido de origen mesodérmico. Se caracteriza por sus componentes: células, sustancia (matriz), fibras y sustancia fundamental. Funciones y características : soporte metabólico (para otros tejidos, a partir de este entran o salen cosas, nutrientes), soporte estructural (también para otros tejidos), está implicado en respuesta inmune, inflamación y renovación, altamente vascularizado, muy bien inervado y altamente heterogéneo. Hay fibras elásticas, colágenas o reticulares, etc. Fig. 2.1. Tejido conjuntivo. CÉLULAS DEL CONJUNTIVO
- Fijas : fibroblastos, células madre mesenquimáticas (MSCs), macrófagos, adipocitos.
- Móviles : macrófagos (unos son fijos y otros son móviles), eosinófilos, basófilos, neutrófilos, linfocitos, monocitos, células cebadas o mastocitos, células plasmáticas. A) Fibroblasto: “formador de fibras”. Cuando es una célula adulta se denomina fibrocito. Células alargadas con núcleos alargados al MO. Al ME son células alargadas con estructura glomerular, puede estar activo formando fibras (fase glomerular) pero si no está activo hay alguna mitocondria, algún Golgi, etc. (poca cosa). El RE está asociado a la formación de fibras. Alrededor del fibroblasto hay fibras de colágeno, entre ellas hay sustancia fundamental. En algunos casos, en determinados tejidos a los fibroblastos se les denomina células reticulares, que forman colágena tipo III. NO es un reticulocito (un reticulocito es un glóbulo rojo inmaduro). B) Célula madre mesenquimática (MSCs): en general, son células de aspecto “fibroblastoide”, es decir, que se parecen a los fibroblastos, pero son más pequeñas. Su desarrollo granular no es especial, tienen un poco de todo sin nada en especial. También tienen prolongaciones. Su característica más importante es que sirven de precursores, poseen una gran potencialidad, son capaces de generar diferentes estirpes celulares.
NO es correcto llamarlas células mesenquimáticas. Explicación: en el embrión hay un tejido que es el precursor del resto, el mesénquima. Aquí hay dos tipos de células: células mesenquimáticas y células madre mesenquimáticas. Se parecen mucho morfológicamente pero no son iguales en el carácter. En el adulto hay fibroblastos y células madre mesenquimáticas, es decir, que las células mesenquimáticas dan lugar entre otras, a los fibroblastos, pero las células madre mesenquimáticas siguen siendo las mismas. En el conjuntivo aparecen las células estromales, cuyo nombre correcto sería MSC también, pero cuando se habla de una estromal es en ciertos sitios definidos, es raro que se hable de ellas. C) Adipocitos: Siempre están en el tejido conjuntivo y suele ser en forma de células dispersas. Con ingestas abundantes proliferan y al final forman el tejido adiposo. Se explican luego en el tejido adiposo. D) Macrófagos: Célula muy heterogénea en cuanto a morfología (de alargado a redondeado), célula grande (20 μm). Es un gran fagocito. Es difícil distinguirlos al microscopio sin un marcaje especial. Son muy regulares con muchísimas digitaciones, pseudópodos, etc. y abundantes gránulos que son lisosomas (al ME). 2 funciones básicas : -‐ Recambio del conjuntivo: son capaces de fagocitar tanto células conjuntivas viejas y muertas como sustancia fundamental y restos del material de alrededor. -‐ Sistema inmune: fagocitan patógenos, son células presentadoras de antígenos (APC) y son capaces de secretar citoquinas (sustancias que actúan sobre la proliferación, quimiotaxis etc. del S.I.). Estas funciones hacen que se localicen normalmente próximos a vasos sanguíneos. Son una población un tanto heterogénea porque a veces se adaptan mucho a los tejidos y pueden recibir distintos nombres. Por ejemplo: la microglía es un macrófago, células de Kupffer (macrófagos hepáticos), osteoclastos (macrófagos del hueso). El macrófago aparece en el tejido conjuntivo en 2 estados: activo e inactivo. Cuando está activo se dice que es un macrófago móvil y cuando está inactivo es una célula fija. Dependiendo de esto también cambia su aspecto. Inactivos son más alargados, etc. y se asocian a la fibra donde están. A veces sufren transformaciones. Por ejemplo: en una situación de infección prolongada, los macrófagos proliferan, se juntan unos con otros y forman acumulaciones de macrófagos epiteloides (epiteloide porque se juntan unos con otros con poca sustancia intercelular). A veces llegan a fusionarse unos con otros. La célula de Langhans se genera por la fusión de diferentes macrófagos. Célula gigante de cuerpo extraño: cuando por ejemplo metes un material indigerible en un órgano, los macrófagos lo rodean y al final se fusionan (macrófago multinucleado con cuerpo extraño dentro), típico de pulmón. à Inciso: Intento de clasificación primero con el “sistema retículoendotelial” o SER: agrupa a cualquier célula que sea capaz de fagocitar sin tener en cuenta su origen. (1924) En 1 969 aparece el sistema fagocito-‐monocito o SFM: aquí están los fagocitos que derivan de monocitos. Por error aquí se metieron los macrófagos y las células dendríticas (DC) pensando que ambas tenían el mismo precursor. La célula dendrítica tiene otro origen. Incluye todos los macrófagos “profesionales”. Los macrófagos son la célula madura de monocito.
Fig. 2.3. Activación y acción de los mastocitos. F) Células plasmáticas Gran tamaño, 20-‐ 30 μm o más, hasta 60 μm más o menos. Núcleo a veces lateralizado con cromatina en rueda de carro. Cuando es adulta o está totalmente diferenciada se ve RER, un poco de Golgi y poco más. RER más desarrollado que existe. Ahí se sintetizan los At. Deriva de linfocitos B, se producen de forma que el Ag provoca que el linfocito prolifere y algunos se diferencien a plasmáticas (los demás quedan como linfocitos B de memoria). La llegada de una segunda exposición al Ag provoca de nuevo la aparición de plasmáticas y linfocitos B de memoria, pero hay diferencias en la respuesta ya que la rapidez de respuesta es mucho mayor en la segunda por las células de memoria que en la primera. En la segunda respuesta las plasmáticas ya derivan de las de memoria. Plasmablasto: intermediario entre el linfocito B y la célula plasmática. Fig. 2.4. Activación célula plasmática. Marcadores para reconocer las plasmáticas: El CD78 y el CD138. También hay un CD127, en la transformación del linfocito B a plasmática, porque el linfocito B no lo presenta, la plasmática sí.
SUSTANCIA INTERCELULAR
Siempre hay células y vasos sanguíneos, y tiene que haber un continuo recambio entre la sangre y la célula, aunque no se toquen, pero pasan a través de la sustancia intercelular que puede ser de distintas formas. Hay dos tipos:
- Fibras También es fundamental, ya que permiten sobrevivir al tejido conjuntivo y lo mantienen en buenas condiciones. Las fibras soportan fuerzas de tracción. Puede que se deforme o no, dependiendo del tipo. Existen 3 tipos: colágenas, elásticas y reticulares. A. Colágenas: Al MO aparecen con formas fibrilares onduladas, unas más gruesas y otras más finas. Entre 0.3 μm y 30/50 μm. Es complicado establecer ese grosor porque a veces no se sabe si son dos pegadas o una. Al ME aparecen o cortadas longitudinalmente o transversalmente. Aparece una estriación en el corte longitudinal al acercarlo un poco. Bandas claras y oscuras. Resultan de la asociación de moléculas de colágena, que son cilindros formados por 3 cadenas, cada una es una cadena α y no tienen porque ser iguales entre sí. El tamaño de estas moléculas es de 300 nm de longitud y unos 1.5 nm de diámetro. Composición de aminoácidos: 30% de glicina, el más abundante. También tienen bastante prolina (10%), pero la mayoría hidroxiladas (hidroxiprolina e hidroxilisina). Hidroxiprolina e hidroxilisina permiten formar puentes entre las cadenas y las fibras. Mantienen la estructura. Pro suele ser de cadenas y Lys de moléculas. Fig. 2.5. Fibras de colágena. Cadenas α: unos 40 genes que las producen. Tipos de colágena unas 20.000, repartidas de forma heterogénea. La α-‐1 es el 90% del colágeno del organismo. El resto son las demás cadenas (muchos tipos pero en poca cantidad). Colágenas fibrilares: I,II,III. Son fibrillas. Colágenas de tipo laminar: IV. No fibrilar: Unas rodean a las células (forma moléculas pequeñas) y otras que son colágena de tipo facie que no forman fibrillas nunca, puentean entre las fibras y las moléculas. XVII, XVIII.
- RE. El ribosoma introduce el péptido en el interior del RE/Golgi: procadena. Cuando ya se sintetiza, se pierde el fragmento que lo transforma en procadena.
- Procesos de glucosilación. La presencia de aa con estos hidroxilos y demás, hacen que interaccionen las cadenas. Procolágena.
- Los extremos N y los C no interactúan. Esto sirve para que la fibra no crezca dentro del orgánulo al polimerizar y no rompa la célula.
- Golgi: sigue el proceso de glucosilación (glucosilación de tipo N).
- Secreción en forma de procolágena.
- Llega la colágeno peptidasa y corta los profragmentos. Si falla esta enzima se produce el síndrome de Ehlers Danlos (piel que se estira mucho).
- Ahora ya sí que interactúan unas con otras. Autoensamblaje: formación fibrilla.
- Fibrillas de colágena por puentes (por las facies) se asocian y forman las fibras. Fig. 2.8. Ensamblaje de las fibras de colágena. à La vitamina C produce escorbuto por la deficiente glucosilación. à La colágena tiene unos 1000 aa. Bases por las que cada tejido permite tener su propia colágena:
- Sintetizar distintas cadenas α, dependiendo del tejido.
- Modificaciones post-‐traduccionales distintas.
- Estas fibras y fibrillas pueden variar su interacción con las moléculas de la matriz.
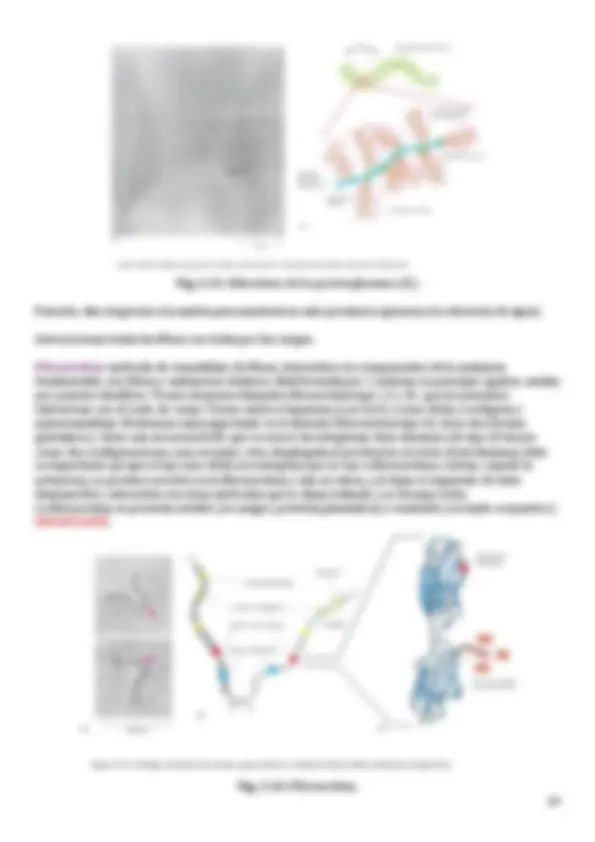
B. Fibras de elastina Son fibras bastante más delgadas. Aproximadamente 1-‐2 micras de diámetro. Al MO aparecen normalmente dispersas, pero en algunos casos se asocian y forman láminas, típicas de vasos sanguíneos. Al ME tienen el componente amorfo (90%) (elastina). Este componente amorfo está rodeado por el componente fibrilar (10%) (fibrilina la más importante). Si hay problemas con la fibrilina, se puede producir el síndrome de Marfan (extremidades más largas de lo normal). La fibra elástica impide que las estructuras crezcan más, como el hueso, etc. La elastina es un péptido o una cadena de unos 750 aa de longitud. Tiene bastante Pro y Lys pero casi sin hidroxilar, algo hay para las interacciones, pero mucho menos que la colágena. En algunas zonas también hay acumulación de aa no polares como Ala o Leu. La elastina se estructura para deformarse y luego recuperar la estructura original. Por ello se disponen en sitios que sufren modificaciones y que necesitan volver a su configuración original como la piel o los vasos sanguíneos. Esta función se permite por la disposición de la elastina. Forma en espiral al azar porque no sigue un patrón fijo. En el dibujo se ve una especie de gancho y otra zona alargada. En la alargada están los aa hidroxilados que forman puentes, y en el gancho están los aa no polares (como no son hidrosolubles, se pliegan). Lisil oxidasa: interacción entre unas Lys y otras. Al estirarse se despliega la fibra y al eliminar la fuerza vuelve a su configuración original. En las fibras interactúan 4 Lys y forman la isodesmosina, que es una estructura formada por la cola de 4 Lys como mínimo. Zona de interacción de unas elastinas con otras. à Latinismo: enfermedad por Lathyrus odoratus : almorta (gachas). La almorta tiene ODAP (oxalildiaminopropiónico), muy parecida al Glutamato, afecta a la lisil oxidasa por lo que hay problemas para formas colágenas y elásticas. Fig. 2.9. Fibras de elastina.
Fig. 2.13. Estructura de los proteoglucanos (II). Función: dan turgencia a la matriz para mantenerse ante presiones (gracias a la retención de agua). Interaccionan todas las fibras con todas por las cargas. Fibronectina: molécula de ensamblaje de fibras. Interactúa con componentes de la sustancia fundamental, con fibras y sustancias celulares. Está formada por 2 cadenas en principio iguales, unidas por puentes disulfuro. Tienen dominios llamados fibronectina tipo I, II y III, que les permiten interactuar con el resto de cosas. Tienen unión a heparina (a un GAG), a una célula, a colágena y autoensamblaje. El dominio más importante es el dominio fibronectina tipo III, tiene dos formas globulares y tiene una secuencia RGD que reconoce las integrinas. Esos dominios de tipo III tienen como dos configuraciones, una cerrada y otra desplegada al producirse un tirón (beta-‐láminas). Esto es importante porque si hay una célula con integrina que se une a fibronectina y actina, cuando la actina tira, se produce un tirón en la fibronectina y esta se estira, y al dejar el segmento de beta-‐ láminas libre, interactúa con otras moléculas que lo dejan trabado y se forman redes. La fibronectina se presenta soluble (en sangre, proteína plasmática) e insoluble (en tejido conjuntivo). IMPORTANTE. Fig. 2.14. Fibronectina.
Recambio de la matriz extracelular Con respecto a las células: se cambian de forma normal. Las más habituales: fibroblastos + adipocitos, derivan de la célula madre mesenquimática. El resto son todo sanguíneas, derivan de médula ósea. Matriz: en el fondo son proteínas y azúcares. Así que se degrada y se sintetiza una nueva. Hay varias familias de enzimas que degradan materia (proteasas): metaloproteasas (necesitan un cofactor metálico) y serín proteasas. Dentro de las metaloproteasas hay varias familias, especialmente 3: MMP, ADAM y ADAMT (degradan matriz). MMP: metaloproteasa de matriz. ADAM: tiene carácter de desintegrina: bloquea integrinas, suelta a la célula de la matriz y tiene fibras sueltas. Elegantina (veneno de serpiente) antes se usaba en las operaciones de corazón porque bombeaban la sangre hacia otra parte y usaban estas moléculas para que no se pegaran a las paredes de la bomba. ADAMT: igual que las ADAM pero también con Trombospondina. Dentro de las serín proteasas: -‐ Colagenasas -‐ Elastasa -‐ Agrecanasa: rompe agrecano -‐ Estromelisina Hay 3 mecanismos de regulación de proteasas:
- Muchas de ellas se secretan en forma de propéptidos (proenzima) que sólo se activan al cortarse ese fragmento de más.
- Por ubicación en determinados lugares: confinamiento de receptores. Sobre todo sucede en casos de avance, la célula va eliminando la matriz de delante para poder moverse. Para esto posee unos receptores sobre los cuáles están las enzimas. Sólo son activas mientras están unidas al receptor.
- Secreción de inhibidores: TIMPs (inhibidores de tejido de metaloproteasa) y Serpinas (inhibidor de serín proteasa). Fig. 2.15. Regulación de proteasas.
Fig. 2.16. Lámina basal. Funciones : la presentan todos los epitelios y algunas células están totalmente rodeadas. Las células musculares están siempre rodeadas de lámina basal, igual que los adipocitos y la células de Schwann. Función de aislamiento, mecánica (un pequeño defecto en la lámina basal puede producir una distrofia, el movimiento tira del tendón, el tendón de la lámina basal y la lámina basal de la célula). Determinan polaridad (esa es la zona basal). Controlan proliferación y diferenciación. En el caso de las células madre, son muy importantes para la conformación del nicho molecular (la célula que toca la lámina basal y la que deriva de esta, toman caminos distintos). Filtración en el glomérulo renal. En algunas patologías son muy importantes: distrofias musculares y tumores (las células tumorales se vuelven malignas cuando son capaces de degradar las láminas basales y se pueden mover). Síntesis mixta : en principio la sintetiza la célula a la que rodea y por los fibroblastos próximos. Estructura : molecularmente complejas. Hay 4 moléculas que representan el 95%: -‐ Colágeno tipo IV: colágeno con partes que conectan entre sí y forman redes. -‐ Laminina: Heterotrímero formado por α-‐β y γ. Interaccionan con las otras. Dos extremos para autoensamblaje. Otros: interacción con integrinas, afinidad con distroglucano y con perlecano. -‐ Perlecan -‐ Nidógeno (entactina) Fig. 2.17. Estructura de la lámina basal. Tania Varela Chorén
-‐^2015