Genètica Epigenètica i sexe
1
EPIGENÈTICA
Al llarg del desenvolupament es modifica de forma reversible el material hereditari implicant a múltiples RNA, canvis
en el DNA, les histones i la cromatina. En cap cas és causat per una modificació de la informació genètica. Per tant, no
es modifica la informació (la seqüència del DNA), sinó l’expressió de la d’aquesta informació genètica. Aquests canvis
relacionats amb el control de l’expressió gènica. Son canvis epigenètics.
Al llarg del desenvolupament del conjunt de cèl·lules tenim el mateix material genètic, però s’expressen diferents gens
en diferents moments del desenvolupament segons la cèl·lula a la que ens trobem; aquesta es la raó de l’existència de
cèl·lules diferenciades. Les proteïnes reguladores i el RNA no codificat són els encarregats de dur a terme una
regulació de l’expressió del DNA, és a dir, de la realització del control de la informació.
Així doncs, la epigenètica esta oferint una explicació sobre la manera en que els canvis fora de la seqüència del DNA,
poden influir en el fenotip i com s’hereten aquest canvis.
1. Canvis epigenètics
Els canvis genètics venen donats per mutacions en al·lels nuls, al·lels hipomòrfics, al·lels hipermòrfics i al·lels
neomòrfics. Per tant, aquest canvis deguts a mutacions son canvis genètics, irreversibles; a no ser que una segona
mutació compenses la modificació causada pel primer.
En canvi, els canvis epigenètics modifiquen l’expressió dels gens a partir d’alteracions i conseqüentment són
reversibles. Formen part del genoma, està prescrit que aquestes modificacions es produiran.
Considerarem tres tipus de mecanismes moleculars que alteren l’estructura de la cromatina i son la base de
nombrosos fenotips epigenètics: canvis dels patrons de metilació del DNA, modificacions químiques de les histones i
molècules de RNA que afecten l’estructura de la cromatina i l’expressió gènica.
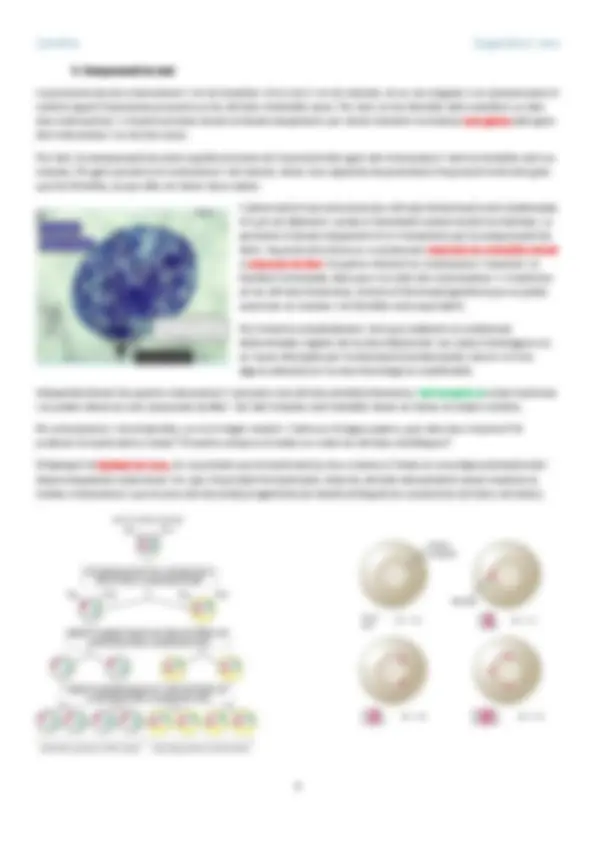
1.1 Metilació del DNA
El procés de metilació es defineix com
l’addició de grups metil a les bases
nucleotídiques, la més comuna, és la
metilació de la citosina per produir 5-
metilcitosina. Aquest canvi s’associa amb la
repressió de la transcripció.
Aquesta metilació del DNA es produeix en
bases de citosina que son adjacents a
nucleòtids de guanina, i passen a formar els
anomenats dinucleòtids CpG; els quals es
situen de manera diagonal entre si al llarg
de la doble cadena de DNA, de manera que
hi ha grups metil a ambdós cadenes de
DNA:
5’-𝐶𝑚𝐺-3’
3’-𝐺𝐶𝑚-5’