


Atlas de Histología Animal y Vegetal
MEMBRANA CELULAR
Manuel Megías, Pilar Molist, Manuel A. Pombal
DEPARTAMENTO DE BIOLOGÍA FUNCIONAL Y CIENCIAS DE LA SALUD.
FACULTAD DE BIOLOGÍA. UNIVERSIDAD DE VIGO.
(VERSIÓN: JULIO 2017)























Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Biología Celular, Profesor: , Carrera: Biología, Universidad: USC
Tipo: Apuntes
1 / 31

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Este documento es una edición en pdf del sitio http://webs.uvigo.es/mmegias/inicio.html
y
ha sido creado con el programa Scribus
(http://www.scribus.net/)
Todo el contenido de este documento se distribuye bajo la licencia Creative Commons del tipo BY NC SA (Esta licencia permite modificar, ampliar, distribuir y usar sin restricción siempre que no se use para fines comerciales, que el resultado tenga la misma licencia y que se nombre a los autores).
Tras dejar el espacio pericelular en nuestro viaje hacia la célula tropezamos con la membrana plasmática de la célula. Ésta es una estructura vital. La rotura de la membrana plasmática durante más de unos pocos segundos lleva irremisiblemente a la muerte celular. Es una barrera física que separa el medio celular interno del externo. En las células eucariotas, y en algunas procariotas, también hay membranas intracelulares que delimitan a los orgánulos, separando el medio interno del orgánulo del citosol.
Composición y estructura. Las membranas celulares están formadas por lípidos, proteínas y, en menor medida, por glúcidos. La estructura y la organización de las membranas celulares, así como sus propiedades, están condicionadas fundamentalmente por los lípidos. Éstos son moléculas anfipáticas, con una parte hidrofílica y otra hidrofóbica, que se disponen formando una bicapa lipídica donde las partes hidrofóbicas se encuentran en el centro de la membrana y las hidrofílicas en contacto con el agua. Entre los lípidos se anclan las proteínas denominadas integrales. Las proteínas transmembrana son proteínas integrales que poseen secuencias de aminoácidos hidrofóbicos entre las cadenas de los ácidos grasos de los lípidos, y
dominios hidrofílicos que están en contacto con la solución acuosa intra y extracelula. También hay proteínas asociadas a una u otra superficie de la bicapa lipídica y que pueden disociarse de ella. Los glúcidos no aparecen en todas las membranas, por ejemplo en algunas intracelulares, pero son abundantes en la que delimita la célula con el medio externo, la membrana plasmática. Se localizan en la superficie extracelular. Los glúcidos se encuentran unidos covalentemente a los lípidos o a las proteínas.
Por tanto las membranas son como láminas extensas que cuando se observan en secciones transversales, perpendiculares a sus superficies, con el microscopio electrónico presentan un aspecto trilaminar: dos franjas oscuras que corresponden con las partes hifrofílicas de los lípidos y una franja clara más ancha entre ellas que son sus cadenas de ácidos grasos. A esto se denomina unidad de membrana y es así para todas las membranas celulares. El espesor de las membranas varía entre los 6 y los 10 nm, lo cual indica que no todas las membranas son exactamente iguales.
Las propiedades fisiológicas y estructurales de las membranas dependen de la proporción y del tipo de
Esquema de la organización de una membrana plasmática según la visión actual del modelo de mosaico fluido de Singer y Nicolson (1972). Es una bicapa fluida estructurada por los lípidos pero heterogénea en su organización. Determinados lípidos se asocian entre sí para formar agrupaciones más densas denominadas balsas de lípidos, en la cuales se sitúan ciertas proteínas por afinidad eléctrica. El colesterol se localiza entre las cadenas de ácidos grasos, cerca de la zona hidrofílica ("cabezas" de los lípidos). Las proteínas transmembrana comunican el exterior (arriba) con el interior (abajo) de la célula. Los glúcidos se localizan en la parte extracelular formando el glucocálix. En este esquema no se muestran las interacciones con la matriz extracelular ni con las moléculas del citoesqueleto. (Modificado de Edidin, 2003 y Nicolson 2014)
moléculas que las componen: lípidos, proteínas y glúcidos. Así, la membrana de los eritrocitos de rata contiene un 50 % de lípidos, un 40 % de proteínas y un 10 % de glúcidos. Una proporción similar a ésta es la más común entre las membranas plasmáticas de todas las células animales, con algunas excepciones. Por ejemplo, la mielina formada por las membranas plasmáticas de las células de Schwan, que rodean a los axones situados fuera del sistema nervioso central, contienen un 80 % de lípidos y un 20 % de proteínas. Las membranas intracelulares suelen contener una mayor proporción de proteínas que la membrana plasmática. La mayor diferencia la encontramos en las mitocondrias donde el porcentaje de proteínas de su membrana interna llega hasta el 80 %. Por supuesto, lípidos, proteínas y glúcidos son grupos heterogéneos de moléculas y también las membranas celulares se diferencian en la composición y en la proporción de distintos tipos de lípidos, de proteínas y de glúcidos. Además, como dijimos anteriormente, las membranas están en una constante renovación que permite a la célula cambiar su composición.
Propiedades. Parte de las funciones de las membranas son debidas a sus propiedades físico químicas: a) es una estructura fluida que hace que sus moléculas tengan movilidad lateral, como si de una lámina de líquido viscoso se tratase; b) es semipermeable, por lo que puede actuar como una barrera selectiva frente a determinadas moléculas; c) posee la capacidad de romperse y repararse de nuevo sin perder su organización, es una estructura flexible y maleable que se adapta a las necesidades de la célula; d) está en permanente renovación, es decir, eliminación y adición de moléculas que permiten su adaptación a las necesidades fisiológicas de la célula.
Funciones. Cada tipo de membrana está especializada en una o varias funciones dependiendo
del compartimento celular del que forme parte. Entre las múltiples funciones necesarias para la célula que realizan las membranas están la creación y mantenimiento de gradientes iónicos, los cuales hacen sensible a la célula frente a estímulos externos, permiten la transmisión de información y la producción de ATP, son necesarios para la realización del transporte selectivo de moléculas , etcétera. Las membranas también hacen posible la creación de compartimentos intracelulares donde se realizan funciones imprescindibles o la envuelta nuclear que encierra al ADN. En las membranas se disponen múltiples receptores que permiten a la célula "sentir" la información que viaja en forma de moléculas por el medio extracelular. Por ejemplo, dan a las neuronas sus propiedades y capacidades, también a las musculares. Tambén poseen enzimas asociadas que realizan numerosas actividades metabólicas, como la síntesis de celulosa o de ácido hialurónico, fosforilaciones, producción de energía, síntesis de lípidos, etcétera. Como hemos visto en el apartado anterior, la adhesividad celular a la matriz extracelular o a otras células en los tejidos animales se debe a las moléculas presentes en la membrana plasmática.
En los siguientes apartados veremos los componentes moleculares, para después tratar las propiedades de las membranas celulares y algunas de sus funciones más importantes. En capítulos posteriores veremos que las membranas celulares de los orgánulos participan de forma determinante en sus funciones, en el trasiego de moléculas en el interior de la célula mediante el denominado tráfico vesicular, así como en la incorporación y liberación de macromoléculas entre el interior y el exterior celular en los procesos de endocitosis y exocitosis, respectivamente.
constituyen la parte hidrofóbica (fobia por el agua) de los glicerofosfolípidos y son los que constituyen la parte interna de las membranas.
El glicerol hace de puente entre los ácidos grasos y la parte hidrofílica (apetencia por el agua). Este componente hidrofílico está formado por un grupo fosfato al que se pueden unir una variedad de moléculas, tales como la etonalamina, colina, serina, glicerol, inositol, el inositol 4,5 bifosfato, etcétera. Estos componentes son los que dan nombre a los distintos tipos de glicerofosfolípidos. El tipo fosfatidilcolina representa más del 50 % de los fosfoglicéridos en las membranas eucariotas.
Esfingolípidos
Deben su nombre a que poseen una molécula de esfingosina, un alcohol nitrogenado con una cadena carbonada larga, a la cual se le une una cadena de ácido graso, formando la estructura básica denominada ceramida (ver figura ). A la ceramida se le une una parte hidrofílica que puede de ser de diversa naturaleza. Por tanto queda una estructura similar a la de los glicerofosfolípidos, dos cadenas hidrofóbicas unidas a una estructura hidrofílica. Los esfingolípidos constituyen la mayoría de los denominados glicolípidos de las membranas, es decir, lípidos que poseen uno o más azúcares unidos formando parte de su zona hidrofílica. Otro tipo de esfingolípidos son las esfingomielinas que poseen una etanolamina o una colina fosforiladas en sus zonas hidrofílicas. Los esfingolípidos son más abundantes en las mebranas plasmáticas que en las de los orgánulos, y se les propone como lo principales responsables, junto con el colesterol, de la segregación lateral de la membrana en dominios moleculares (balsas de lípidos).
Esteroles
El colesterol es el esterol más importante de las células animales y el tercer tipo de lípido más abundante en la membrana plasmática (hasta el 25 % del total de lípidos), mientras que aparece en pequeñas proporciones en las membranas de los orgánulos celulares (1 % en el retículo endoplasmático). El colesterol se localiza entre las cadenas de ácidos grasos de los otros lípidos. Es importante para la estructura de la membrana puesto que junto con los esfingolípidos parece contribuir a formar heterogeneidades en la distribución molecular y también participa en ciertos procesos metabólicos vitales como la síntesis de hormonas esteroideas o de sales biliares, entre otras. El colesterol no aparece en las membranas de las plantas, en algunas células eucariotas, ni en las bacterias, pero estas células tienen otro tipo de esteroles. Los estéroles son esenciales para la integridad y funcionamiento de las membranas eucariotas. Sirven para modular la rigidez, la fluidez y la permeabilidad.
La distribución de lípidos depende de la membrana
Aunque hay tres grandes grupos de lípidos de membrana (glicerolípidos, esfingolípidos y esteroles), hay miles de tipos moleculares diferentes de lípidos distribuidos por las membranas celulares. La composición de lípidos varía entre las membranas de los diferentes compartimentos membranosos de la célula. Se propone que la propia identidad de los orgánulos viene determinada por la composición de sus membranas, tanto proteínas como lípidos. Por ejemplo, la membrana plasmática tiene una composición lipídica diferente a la membrana del retículo endoplasmático o a la del aparato de Golgi. Estas diferencias se mantienen a pesar del flujo constante de lípidos desde sus compartimentos de síntesis, principalmemente el retículo endoplasmático, hasta otras Esfingolípidos.
Colesterol
membranas como la plasmática o los endosomas, estando este transporte mediado por vesículas, transportadores y contactos directos entre membranas.
La cantidad y tipo de lípidos varía entre membranas. Por ejemplo, aunque todas la membranas tienen fosfatidil colina, este glicerolípido es más abundante en las membranas del retículo endoplasmático. En la membranas post Golgi, es decir, membrana plasmática y endosomas, la concentración de esfingolípidos y colesterol es mayor que en el retículo y en las membranas del aparato de Golgi. Las mitocondrias tienen, aparte de otros más extendidos, lípidos de membrana propios como el fosfatidilglicerol y la cardiolipina, que sintetizan ellas mismas.
Esto significa que deben existir mecanismos de segregación por compartimentos membranosos de las diferentes especies de lípidos de membrana. Se han propuesto diversos mecanismos que contribuyen a esta distribución desigual de lípidos:
Síntesis diferencial. La concentración de determinadas especies de lípidos viene condicionada por su lugar de síntesis. Por ejemplo, los esfingolípidos se terminan de ensamblar en el aparato de Golgi y son los compartimentos post Golgi donde más abunda, pero están ausentes en el retículo endoplasmático. La fosfatidilcolina se sintetiza en el retículo endoplasmático y es ahí donde más abunda. Sin embargo, no siempre es cierto puesto que, por ejemplo, el colesterol se sintetiza en el retículo endoplasmático, pero es más abundante en membranas post Golgi. Esto es debido a que es transportado rápidamente a otras membranas. Además, durante los procesos de síntesis de unos lípidos se emplean a otros como donantes de partes moleculares por lo que a
Distribución de los lípidos más abundantes en diferentes compartimentos membranosos celulares. Las flechas azules delgadas indican trasiego de lípidos transportados por las vesículas, las rojas gruesas la transferencia de lípidos entre membranas muy próximas, las flechas rojas delgadas indican la transferencia de lípidos entre membranas mediante transportadores proteicos, los cuales actúan entre varios compartimentos (no indicado). En la imagen inferor se representa el incremento de la concentración de colesterol desde el retículo endoplasmático hasta la membrana plasmática. PC: fostatidil colina, PE: fosfatidil etanolamina, PI: fosfatidil inositol, PS: fosfatidil serina, SM: esfingomielina, ISL: esfingolípido inositol, CL: cardiolipina, MBP: bis monoacilglicerol fosfato (modificado de van Meer et al., 2008).
Proporción aproximada de los lípidos totales de un célula típica de mamífero (tomado de Vance 2015)
Las proteínas son las responsables de muchas de las funciones que tienen su base en las membranas celulares. El contenido de proteínas de las membranas respecto a los lípidos es varibale dependiendo del tipo de membrana, pero puede ser de cerca de 1:40 en número de moléculas, lo que indicaría que las membranas contienen bastantes proteínas. Incluso puede ser más alta en aquellas membranas con funciones netamente metabólicas como la membrana interna de las mitocondrias. Hay dos grandes grupos de proteínas relacionadas con las membranas: las integrales y las asociadas.
Proteínas integrales
Las proteínas integrales son aquellas que forman parte de la mambrana de manera permanente. Se dividen en tres grupos: las transmembrana, la integradas en una monocapa y las unidas covalentemente a moléculas que forman parte de la membrana.
Las proteínas transmembrana poseen tres tipos de dominios en sus secuencias de aminoácidos: uno
extracelular, uno intracelular y otro en el interior en la propia membrana. En el intracelular poseen secuencias de aminoácidos con radicales hidrofóbicos que se sitúan entre las cadenas de ácidos grasos de los lípidos de la membrana, mientras que los dominios intra y extracelular poseen secuencias de aminoácidos con radicales hidrofílicos. Estas proteínas son mayoritariamente producidas en el retículo endoplasmático y repartidas por otras membranas de la célula principalmente mediante el tráfico vesicular, como veremos en capítulos posteriores.
Existen proteínas transmembrana cuya cadena de aminoácidos cruza una sola vez la membrana mientras que otras pueden hacerlo hasta 7 veces. Muchas proteínas transmembrana realizan su función cuando se asocian con otros polipéptidos también integrales para formar estructuras oligoméricas (más de un elemento). Las funciones son muy variadas, pero destacan la adhesión llevada a cabo por las integrinas, cadherinas, selectinas y otras; el intercambio de iones, calcio, sodio o potasio, entre ambos lados de la membrana como hacen las bombas de iones y los canales iónicos, los cuales permiten gradientes que hacen posible, por ejemplo, la sínteis de ATP; permitir el cruce de moléculas como la glucosa por parte de los transportadores; algunas participan en la comunicación celular y actúan como receptores de señales como los que reconocen los factores de crecimiento, neurotransmisores, hormonas y otros. En este último caso, la organización en dominios extracelular e intracelular permite una comunicación entre ambos lados de la membrana, lo cual hace que una información extracelular, por ejemplo una hormona reconocida por el dominio extracelular, provoque un cambio conformacional en el dominio intracelular y esto desencadene una cascada de señales intracelulares que alteren la fisiología celular, incluso la expresión génica.
Hay proteínas integrales que forman parte de la membrana pero que no son transmembrana. Un tipo son aquellas que poseen parte de su secuencia de aminoácidos integrada entre los lípidos de una de las monocapas de la membrana, y por tanto tienen un dominio extramembranoso que puede ser citosólico o extracelular, según en que monocapa de la membrana
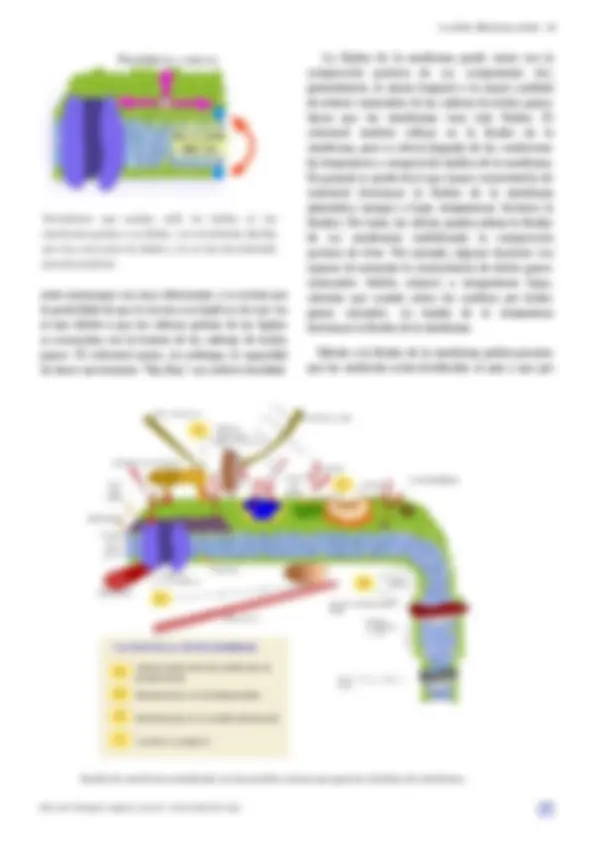
En este esquema se muestran los principales tipos de proteínas transmembrana. Las hay con un solo cruce como la glicoforina o con varios como algunos receptores. En ambos casos la secuencia o secuencias de aminoácidos localizadas entre las cadenas de ácidos grasos adoptan una conformación en alfa hélice. La aquaporina, un canal que cruza numerosas veces la membrana, posee secuencias de aminoácidos de la zona hidrofóbica que se disponen en hebras beta. (Modificado de Pollard et al., 2007)
estén integradas. Otro tipo de proteínas integrales, no transmembrana, son aquellas que se encuentran ancladas por enlaces covalentes a moléculas de la membrana, como puede ser un ácido graso. En este caso toda la secuencia de aminoácidos es extramembranosa, pudiendo ser citosólica o extracelular. En este tipo podemos también incluir a las proteínas que están unidas covalentemente a los glúcidos de los lípidos. A las proteínas que no son transmembrana se les puede llamar también periféricas puesto que sólo están relacionadas con una monocapa. Las proteínas periféricas Incluyen también a las proteínas asociadas (ver más abajo).
Proteínas asociadas
Las proteínas asociadas a las membranas plasmáticas son aquellas que no forman parte permanente de las membranas, es decir, no son proteínas integrales, y su unión a la membrana se produce por interacciones eléctricas con moléculas de la propia membrana (fuerzas de van der Waals). Estas asociaciones son más lábiles y las proteínas pueden abandonar la membrana con cierta facilidad. Son proteínas hidrosolubles.
Esquema de las principales formas de proteínas periféricas: integrales y asociadas. De izquierda a derecha: proteínas que tienen una parte de su secuencia de aminoácidos inserta en una de las monocapas de la membrana, proteínas que están unidas covalentemente a azúcares de los glucolípidos, proteínas que tienen ácidos grasos unidos covalente, lo que les permite insertarse en la zona hidrófoba de la membrana. Proteínas que interactúan eléctricamente con las cabezas de los lípidos; y proteínas que interactúan con los dominios extramembrana de proteínas transmembrana. (Modificado de Alberts et al., 2002).
La composición química de las membranas hace que posean unas propiedades esenciales para las funciones que desempeñan en la célula. Podemos agrupar dichas propiedades en cinco: semipermeabilidad, asimetría, fluidez, reparación y renovación.
Semipermeabilidad
Esta propiedad es consecuencia del ambiente hidrófobo interno de la membrana creado por las cadenas de ácidos grasos de los lípidos, difícil de cruzar por las moléculas con carga eléctrica neta. Esto permite a las membranas crear compartimentos intracelulares o mantener separados el medio intracelular del extracelular y, por tanto, impedir la libre difusión de diversos tipos de moléculas a su través. Sin embargo, la permeabilidad es selectiva. Las variables que más influyen en la difusión pasiva son la polaridad y el tamaño de la molécula. Así, moléculas pequeñas sin carga, por ejemplo el CO 2 , N 2 , O 2 , o moléculas con alta solubilidad en grasas como el etanol cruzan las membranas prácticamente sin oposición, por un proceso de difusión pasiva. La permeabilidad de la membrana es menor para aquellas moléculas con cargas pero globalmente neutras (el número de cargas negativas iguala al de cargas positivas) como el agua o el glicerol. Se podría pensar que el agua difunde libremente por las membranas pero no es así y por ello en determinadas membranas existen unas moléculas denominadas acuaporinas que facilitan el cruce de la membrana por parte del agua. Es menor aún la capacidad de atravesar la membrana para moléculas grandes neutras pero con cargas, como la glucosa. Sin embargo, es altamente impermeable a los iones y a las moléculas que
también carga neta. Algunos valores del coeficiente de permeabilidad a través de membranas por difusión pasiva son: O 2 : 2.3 cm/s, CO 2 : 0,35 cm/s, H 2 O: 0, cm/s, glicerol: 10 6 cm/s, sodio y potasio: 10 14 cm/s.
La desigual distribución de iones y moléculas entre ambos lados de la membrana es la base para la creación de los gradientes químicos y eléctricos. La medida de esa diferencia de concentración de cargas es lo que se llama potencial de membrana, que se usa para muchas funciones celulares, como por ejemplo la síntesis de ATP o la transmisión del impulso nervioso. La semipermeabilidad de la membrana también permite el fenómeno de la ósmosis, es decir, el flujo de agua hacia donde más concentración de solutos haya. Las células vegetales deben su crecimiento a este proceso. Como veremos más adelante, las moléculas que no cruzan las membranas libremente son interesantes para las células puesto que la variación de sus concentraciones a un lado u otro de la membrana puede actuar como señales o como herramientas. Por ello se han inventado proteínas integrales de membrana que permiten selectivamente el paso de estas sustancias de un lado al otro. Por ejemplo, la contracción muscular se debe a una rotura de ese gradiente eléctrico.
Fluidez y heterogeneidad
Es la capacidad de una molécula que forma parte de una membrana para desplazarse por ella. Las membranas son fluidas, prácticamente son láminas de grasa, donde las moléculas se encuentran en un estado de líquido viscoso. Esto implica que, en teoría, las moléculas podrían difundir y desplazarse por ella sin restricciones. Consideremos un glicerofosfolípido que está situado en la membrana plasmática en su monocapa externa. Tendría dos posibilidades de movimiento: uno lateral donde se desplazaría entre las moléculas contiguas, y otro en el que saltaría a la monocapa interna, movimiento denominado "flip flop". Los dos tipos de movimientos se han demostrado experimentalmente en membranas artificiales pero el primero es mucho más frecuente que el segundo. Una molécula lipídica puede recorrer 30 micras en unos 20 segundos por difusión pasiva lateral, es decir, podría dar la vuelta a una célula de tamaño medio en aproximadamente un minuto. Sin embargo, los saltos
El cruce de la membrana por parte de las moléculas depende de su tamaño y de sus características eléctricas (Modificado de Alberts et al., 2002).
entre monocapas son muy infrecuentes y se estima que la posibilidad de que le ocurra a un lípido es de una vez al mes debido a que las cabezas polares de los lípidos se encuentran con la barrera de las cadenas de ácidos grasos. El colesterol posee, sin embargo, la capacidad de hacer movimientos "flip flop" con relativa facilidad.
La fluidez de la membrana puede variar con la composición química de sus componentes. Así, generalmente, la menor longitud o la mayor cantidad de enlaces insaturados de las cadenas de ácidos grasos hacen que las membranas sean más fluidas. El colesterol también influye en la fluidez de la membrana, pero su efecto depende de las condiciones de temperatura y composición lipídica de la membrana. En general se puede decir que mayor concentración de colesterol disminuye la fluidez de la membrana plasmática (aunque a bajas temperaturas favorece la fluidez). Por tanto, las células pueden alterar la fluidez de sus membranas modificando la composición química de éstas. Por ejemplo, algunas bacterias son capaces de aumentar la concentración de ácidos grasos insaturados (dobles enlaces) a temperaturas bajas, mientras que cuando suben los cambian por ácidos grasos saturados. La bajada de la temperatura disminuye la fluidez de la membrana.
Debido a la fluidez de la membrana podría pensarse que las moléculas están distribuidas al azar y que por
Movimientos que pueden sufrir los lípidos en las membranas gracias a su fluidez. Los movimientos flip flop son muy raros para los lípidos y no se han documentado para las proteínas.
Modelo de membrana actualizado con las posibles causas que generan dominios de membrana.
concretos de la superficie celular. Las células tienen otros mecanismos para confinar proteínas a determinados dominios celulares. Por ejemplo, en las
células epiteliales del digestivo ciertos transportadores y enzimas están localizados sólo en la zona apical y otros en la basal gracias al cierre a modo de cinturón que realizan las uniones estrechas, como vimos en el capítulo dedicado a la matriz extracelular. Tal asimetría es esencial para el funcionamiento de la célula epitelial.
Curvar una membrana es otra manera de crear dominios, y esta curvatura puede ser el proceso inicial de la formación de una vesícula, la extensión de una expansión celular, el cambio o crecimiento de un orgánulo, o simplemente un pliegue que actúa como barrera a las difusión lateral de moléculas.
La maquinaria necesaria para curvar una membrana requiere a su vez un dominio de membrana para llevarlo a cabo. Determinadas composiciones lipídicas o zonas con diferente carga eléctrica son sitios de atracción para la maquinaria. Los fosfoinosítidos son lípidos que participan en este reclutamiento, particularmente PIP2 y PIP3. Son apropiados para esto puesto que su cabeza puede cambiar en carga y estructura mediante modificaciones químicas locales. Estos lípidos son a su vez mantenidos en el sitio por afinidad con las proteías reclutadas. La creación inicial de un dominio para curvar una membrana ocurre también con la fostatidil serina, la cual, cuando cambia de hemicapa por acción de las flipasas, es capaz de ayudar a generar curvatura, y es retenida por proteínas como la caveolina. Los filamentos de actina pueden también controlar la difusión de lípidos, además de proteínas, creando barreras a modo de vallas en superficie citosólica de la membrana, creando así dominios de lípidos sin la intervención del colesterol o los esfingolípidos.
Los microdominios de
Los movimientos de las moléculas pueden estar restringidos por las interacciones directas con la matriz extracelular, con el citoesqueleto, aunque también se pueden limitar los movimientos por la inclusión en las balsas de lípidos o por la disposición del citoesqueleto (imagen de la derecha).
Algunos mecanismos moleculares que curvan las membranas de las células.
lípidos atraen a las proteínas que realmente curvan a las membranas de manera efectiva. Hay proteínas especializadas en crear curvatura de membranas. El domimio proteico BAR (Bin/amphyphysin/Rsv161) es uno de ellos. Lo hace de dos maneras: mediante la creación de un andamiaje curvado sobre el que descansa la membrana o mediante inserciones de secuencias de aminoácidos en la membrana a modo de cuñas. Otros ejemplos son las caveolinas que también generan curvatura para formar caveolas, las tetraspaninas que generan túbulos en las membrana, los ESCRT que son responsables en los endosomas de crear las vesículas de los cuerpos multivesiculares, etcétera. Por último, los filamentos de actina son unas auténticos curvadores de membranas mediante polimerización, mediante la cual empuja a la membrana plasmática hacia afuera creando
expansiones celulares. Muchas de las proteínas capaces de curvar la membrana son también unas activadoras de la polimerización de filamentos de actina.
Otros dominios físicos de las membranas, aunque creados por moléculas presentes en la propia membrana como las proteínas transmembrana, son regiones de diferente espesor o altura. Son formados por proteínas transmembrana que tienen del dominio hidrofóbico más largo de lo habitual y por tanto se acomodan mejor en la membrana cuando se rodean de lípidos con cadenas de ácidos grasos largos. Estas agrupaciones de lípidos y proteínas crean áreas de mayor grosor que excluyen a otras proteínas con secuencias de aminoácidos hidrofóbicos más cortas o a lípidos con cadenas de ácidos grasos con menos átomos de carbono.
Dominio de membrana creado por diferencias de espesor.
frecuente. Los glicerofosfolípidos de cabezas polares grandes pueden salvar la barrera hidrófoba de los ácidos grasos mediante unos transportadores específicos, o traslocasas, localizados en las membranas. Hay tres tipos: flipasas, flopasas y mezcladores ("scramblases"). Estas proteínas se encargan de transportar glicerofosfolípidos entre las dos hemicapas y generar asimetría. Las flipasas transportan lípidos hacia la hemicapa interna, las flopasas hacia la hemicapa externa y las mezcladoras en ambas direcciones. Las mezcladoras no necesitan ATP para llevara cabo su función. Esta desigual distribución de los lípidos se mantiene por la propia dificultad de los movimientos “flip flop”. Por ejemplo, lo esfingolípidos, que no son translocados, permanecen en la monocapa donde se sintetizan: la interna del aparato de Golgi, que se será posteriormente la externa de la membrana plasmática. Más del 80 % de los esfingolípidos de la membrana plasmática se localizan en la monocapa externa.
Estas proteinas se distribuyen desigualmente por las diferentes membranas de la célula para aportar a cada membrana sus asimetría lipídica característica. Incluso dentro de una misma familia puede haber subtipos con diferente regulación. Como se mencionó antes, la apoptosis provoca la rotura de la asimetría de la membrana plasmatica que desencadena la fagocitosis de la célula. Esta rotura se debe a una activación puntual de una proteína mezcladora que se encuentra en dicha membrana. Sin embargo, otra proteína mezcladora que actúa en el retículo endoplasmático está funcionando siempre, y por eso las dos hemicapas de este orgánulo son muy parecidas. Otro ejemplo, en la membrana del eritrocito hay proteínas mezcladoras que se activan con la entrada masiva de calcio y favorecen la coagulación sanguínea.
Glúcidos. La distribución de los glúcidos, localizados sobre todo en la hemicapa externa de la membrana plasmática, se produce durante su síntesis, primero en el retículo endoplasmático y posteriormente en el aparato de Golgi.
Proteínas. La asimetría de las proteínas se produce durante su síntesis en el retículo endoplasmático, aunque las proteínas asociadas a la cara citosólica se sintetizan en el citosol.
La distribución y organización de las moléculas en las dos hemicapas de las membranas puede ser diferente. Esto es claro para los lípidos en la membrana plasmática (arriba), pero no tanto en la del retículo endoplasmático (abajo). Sin embargo, las proteínas se orientan y disponen de manera asimétrica tanto en las membranas del retículo como en la plasmática. Se indica la carga negativa de la fosfatidil serina para mostrar la diferente distribución de cargas entre ambas monocas de la membrana plasmática, pero no corre lo mismo en la membrana del retículo endoplasmático.
Proteínas encargadas de redistribuir los diferentes lípidos entre las dos monocapas de las membranas. La localización de estas proteínas varía entre orgánulos y entre tipos celulares. Flipasas, flopasas son en realizadad dos familias de proteínas cuyos miembros se distribuyen diferencialmente. (Modificado de Quazi y Molday, 2011).
Rotura y fusión de membranas
Una de las propiedades de las membranas más útiles para la célula es la capacidad de romperse y volver a ser fusionadas. Ello permite que los compartimentos intracelulares puedan ser tremendamente plásticos, es decir, crecer, dividirse, fusionarse, liberar fragmentos en forma de vesículas membranosas en un compartimento que viajan a otro con el que se fusionan, etcétera. Ésta es la base del transporte vesicular que veremos en los apartados siguientes. Esta característica de las membranas es también necesaria durante la etapa de la mitosis denominada citocinesis donde la membrana citoplasmática debe crecer en superficie, romperse y luego fusionarse para formar dos células hijas independientes. Estos procesos de rotura y de fusión de membranas están gobernados principlamente por las proteínas, entre las que se destacan las SNARE (soluble N ethylmaleimide sensitive factor attachment receptor), pero también por las propiedades fisicoquímicas de los lípidos que componen las membranas.
Reparación de membranas
Numerosos procesos naturales o la manipulación experimental de las células provocan la rotura de las membranas celulares. Por ejemplo, en los tejidos vivos sometidos a tensiones hay un proceso de rotura de la membrana plasmática, como ocurre frecuentemente en las células musculares. Pero también en los experimentos de clonación se necesita meter una pipeta, la captación de vectores o ADN supone a veces la poración de las membranas celulares, la propia manipulación supone roturas de membrana. La rotura de la membrana plasmática es letal para la célula si se prolonga más de unos cuantos segundos. Las células cuentan con mecanismos para reparar estos daños y mantener así las diferencias entre el medio interno y externo. Los tejidos que no son capaces de reemplazar a sus células como es el caso del sistema nervioso, estos mecanismos de reparación son especialmente interesantes.
Una rotura de la membrana plasmática rápidamente dispara la entrada masiva de calcio en la célula, lo que produce alteraciones celulares que, si se mantiene durante mucho tiempo, desencadenan la apoptosis (muerte celular programada). Per este incremento de calcio dispara también los mecanismos de reparación. El calcio dispara numerosas vías enzimáticas en
paralelo que confluyen en la generación de vesículas y fusión de estas para sellar la membrana. Curiosamente ni el cloro, ni el sodio, ni el potasio paracen participar en los mecanimos de reparación de las membranas.
Hay dos maneras de sellar la membrana según el tipo de daño que se produzca. Cuando los daños son pequeños (normalmente menores a 0.2 μm) las propiedades de los lípidos de la membrana son suficientes para repararlos. Ello es debido a que los lípidos en el borde de la membrana adoptan una disposición inestable que fuerza a dichos bordes a encontrarse y a sellarse. La rapidez con que este proceso ocurre depende de la tensión de la membrana, que depende a su vez de los puntos de anclaje, bien al citoesqueleto o a la matriz extracelular. Cuando se produce una rotura entra calcio a favor de gradiente de concentración, lo que hace que el citoesqueleto se desorganice parcialmente en la zona dañada y su efecto sobre la membrana disminuye, se rebaja así la tensión y aumenta la velocidad de resellado. Las proteínas ESCRT, que participan en los procesos membranosos de formación de vesículas internas en los cuerpos multivesiculares, también participan en el sellado de pequeñas roturas de membrana (<100 nm de anchura).
Cuando los daños son grandes (más de 0.2 0.5 μm) los bordes rotos libres de la membrana están demasiado lejos para se que puedan autosellar y se pone en funcionamiento un mecanismo de exocitosis masiva, es decir, fusión de vesículas con la membrana plasmática, aunque en este caso también incluye grandes
Cuando las roturas de la membrana son pequeñas, menores a unas 0.2 micras, las propiedades moleculares de los lípidos son suficientes para cerrar el hueco (Modificado de McNeil y Steinhardt, 2003)