¡Descarga Origen cel·lular descripción y más Apuntes en PDF de Fisiopatología solo en Docsity!
La célula: evolución y desarrollo
de los sistemas celulares
Jorge Joel Reyes Méndez
“El objetivo de la biología celular y molecular moderna es interpretar las propiedades de los organismos mediante la estructura de sus moléculas constituyentes.” FRANCOIS JACOB, 1973, en The logic of life.
Objetivos Introducción Desde las moléculas hasta la primera célula Formación de moléculas sencillas en las condiciones prebióticas Síntesis de los polinucleótidos Selección natural de las moléculas automultiplicadoras Moléculas de RNA como catalizadores Transferencia de información desde los polinucleótidos a los polipéptidos Formación de la membrana Primeras células De la célula procariota a la eucariota Células procariotas Reacciones metabólicas Fotosíntesis Respiración Glucólisis La célula eucariota Teoría actual de la endosimbiosis como origen de la célula eucariota
Estructura celular Transporte a través de membranas Transporte pasivo Difusión simple Transporte activo Citosol y citoesqueleto Microtúbulos Microfilamentos Filamentos intermedios Distribución de los filamentos del citoesqueleto en el citoplasma Retículo endoplásmico Aparato o complejo de Golgi Lisosomas Mitocondria Cloroplasto Fotosistemas Núcleo Cromatina y cromosomas El gen desnudo Reproducción de los virus Viroides Priones
Contenido
Capítulo^1
Objetivos
Al finalizar el capítulo, el lector podrá identificar las teorías actuales sobre el origen de los sistemas precelulares y de la vida; interpretar y conceptualizar los procesos evolutivos que llevaron a la aparición de los procariotas; señalar los funda- mentos de la teoría endosimbiótica del origen de las células eucariotas; identificar las diferencias fundamentales entre una célula procariota y una célula eucariota y describir el proceso de síntesis de proteínas en sus partes fundamentales.
Introducción
Hace aproximadamente 4 700 millones de años, según los cálculos más recientes, en uno de los brazos de la galaxia, una nube de gas y polvo se contrajo para formar un disco. Durante dicha contracción, esta nube aumentó su velocidad angular. Hubo un momento en el que el centro del disco fue tan denso, masivo y caliente que fue posible que se llevaran a cabo reacciones nucleares en su interior. Este hecho lo convirtió en una estrella, el Sol. Posteriormente, parte de las partículas de polvo y gas restantes se unieron para dar origen a los planetas. Setecientos millones de años después, es decir, hace cuatro mil millones de años, la Tierra completaba un giro alrededor de su eje en 18 horas. El joven Sol brillaba con sólo el 70% de su energía actual. La atmósfera no contenía oxígeno libre. Había pocos sonidos, salvo el ruido del viento, el de las erupciones volcánicas, el silbido de la lava al hacer contacto con el agua y las explosiones de los meteoritos al chocar contra la corteza terrestre. No había plantas, animales, bacterias, ni virus. A partir de este punto inicial, la opinión de los cientí- ficos varía ampliamente: no se sabe con exactitud cómo era el planeta Tierra cuando inició la vida. Las rocas que podrían brindar esta información se deformaron hace mucho tiempo a causa de los movimientos de la corteza terrestre. Stanley L. Miller afirma que “los compuestos orgánicos se organizaron en algo que podía automultiplicarse”. Son varios los escenarios posibles. A continuación se exponen los más recientes puntos de vista de científicos destacados, estudiosos del tema. Aunque los acontecimientos que originaron la vida en la Tierra aún son misteriosos, los científicos exploran desde hace algunos años pistas nuevas que se revelan prometedoras. El químico norteamericano Stanley Miller había efectuado en los años cincuenta del siglo XX los primeros experimentos
de reproducción de una sopa primitiva en la que se habrían sintetizado las moléculas que constituyen la materia viva. En la actualidad, esta hipótesis se considera improbable, y parece más verosímil que dichas moléculas se hayan formado en la superficie de determinados minerales. Según este punto de vista, uno de los problemas fundamentales es saber cómo surgieron las moléculas capaces de automultiplicarse en ausen- cia de enzimas, condición indispensable para que se pueda transmitir la información genética. Se sabe que todos los seres vivos están constituidos por cé- lulas (pequeños compartimientos rodeados con una membrana y rellenos de una solución de sustancias químicas). Las formas más simples de vida son células solitarias que se reproducen dividiéndose en dos. Los organismos restantes, incluso el ser humano, constan de agrupaciones de células que efectúan determinadas tareas y que se comunican con otros grupos de células mediante procedimientos de señales complejos. Todos los organismos y todas las células que los consti- tuyen proceden de un antecesor común, pero se desconocen los procesos intermedios. Uno de estos procesos, denomi- nado evolución , tiene lugar por medio de dos mecanismos bien definidos:
- la variación mediante la cual se produce en forma aleatoria un cambio en la información genética
- la selección en favor de los individuos que están mejor preparados para sobrevivir y reproducirse
Por otro lado, se sabe que más que una curiosidad bio- lógica, la endosimbiosis es uno de los motores más poderosos de la evolución del mundo vivo. La endosimbiosis crea con rapidez organismos quiméricos que engendran nuevos linajes, acerca a los socios y favorece transferencias masivas de genes que crean genomas también quiméricos: el genoma nuclear contiene, por consiguiente, genes eucariotas, pero también genes de origen bacteriano procedente de las mitocondrias o de los plastos, con los que coexiste. Estos acontecimientos explican tal vez los saltos evolutivos fundamentales que dieron origen a los grandes linajes del mundo viviente y moldearon la diversidad biológica actual. El mismo hombre puede con- siderarse como una comunidad simbiótica, que está formada por el citoplasma eucariota y las mitocondrias. El hombre es, en realidad, un conglomerado de bacterias, espiroquetas y virus diversos. El producto acabado de una industriosa colonia de bacterias simbióticas que lo armaron, de la misma forma que el hombre construye su hogar y sus ciudades. Este capítulo trata de la progresión desde las moléculas hasta los organismos multicelulares. En seguida se estudian las estructuras fundamentales de las células eucarióticas (cé- lulas que disponen de una anatomía intracelular estructurada a diferencia de las células procarióticas que carecen de estas estructuras).
Escape del H Gases volcánicos
Acumulación de oxígeno
Capa de ozono
Oxígeno actual
Animales y vegetales modernos
Enfriamiento de corteza y atmósfera
Océanos Acumulación de materia orgánica
Desaparición de las moléculas orgánicas del ambiente
Algas
Código genético Glucólisis
Respiración Multicelularidad
Protocélulas
Fotosíntesis
4 500
4 000
3 000
2 000
1 000
500
Formación del sistema solar
Formación de la Tierra
Aparición de cúmulos globulares
Formación de galaxia
Big Bang
Evolución prebiótica
Evolución biológica
500
400
300
200
100
(^05)
4 500
10 000
Hace 1 500 millones de años Vida en tierra firme
Aparición de los dinosaurios
Aparición de los mamiferos
Extinción de dinosaurios
Hombre primitivo
Atmósfera Geosfera Biosfera
Evolución química
Figura 1-2. Escala del tiempo geológico y algunos de los principales hechos del origen de la vida.
CH 4 NH 3 CO 2 H 2
ñ
Agua
Formaldehído Ácido fórmico Cianuro de hidrógeno Ácido acético Ácido láctico Glicina Alanina Urea Ácido aspártico Calor
Cromatografía sobre papel
Trampa
Descarga eléctrica
Figura 1-3. Experimento de Miller.
Los polímeros primitivos pueden haberse formado de varias maneras, por ejemplo, al calentarse los constituyentes orgánicos en seco o al catalizar la reacción altas concentra- ciones de polifosfatos inorgánicos. En condiciones de labo- ratorio, el resultado son polímeros de longitud variable, cuyos componentes se acomodan en forma aleatoria (fig. 1-5). Pero una vez que se ha formado un polímero, éste puede influir en la formación de uno nuevo, ya que puede actuar como “molde” en una nueva reacción de polimerización. En efecto, los nucleótidos tienen la propiedad de emparejarse mediante la formación de enlaces de hidrógeno, de tal manera que un polímero (p. ej., el poli U) induce la formación del polímero complementario (poli A) (fig. 1-6). Probablemente el emparejamiento específico de los nucléotidos (bases) desempeñó un papel decisivo en el sur- gimiento de la vida. Considérese, por ejemplo, un fragmento de RNA que contiene las cuatro bases: adenina , A, uracilo , U, citosina , C, y guanina , G. Gracias a la complementariedad de las bases, si este RNA se añade a una mezcla de nucleótidos activados en condiciones que favorezcan la polimerización, se creará un RNA complementario del primero. Es decir, las muevas moléculas son como el vaciado del molde
original en donde A ha sido sustituido por U, C por G y así sucesivamente. La información contenida en la secuencia original se preserva, de algún modo, en las copias, ya que en una segunda vuelta de polimerización, éstas generan RNA idéntico al original. De esta forma, la información genética contenida en la secuencia de nucleótidos pasa de generación en generación mediante este proceso de emparejamiento de bases complementarias. Los mecanismos de duplicación requieren la presencia de un catalizador. Sin éste, la reacción es lenta y poco eficien- te, y otras reacciones secundarias podrían interferir, lo cual impediría la formación de réplicas exactas. En la actualidad, una proteína cataliza esta reacción, la polimerasa de DNA inexistente en la “sopa prebiótica”. En los tiempos primitivos, iones metálicos y minerales como la arcilla podrían haber cumplido la función de catálisis. Pero, lo que es más impor- tante, el propio RNA pudo haber actuado como catalizador. En efecto, el RNA, además de funcionar como molde para la duplicación, tiene la posibilidad de formar plegamientos intramoleculares que originan superficies complejas, las cuales catalizan reacciones específicas. Posteriormente se tratan los mecanismos propuestos a este respecto.
A
A
A
A
A
A
A
A
G
G
G
G
G
G
G
G
G G
G C
C
C
C
C
C
C
C
C C
C
C
C
C^
U
U^ U
U U
U
U U U U
H 2 O
A
A
A
A
A A
A
G
G
G
G
G G
G
G
G
G C
C
C
C
C
C
C
C
C C
U
U
U
U
U
U
A A A A
U U
H 2 O
U U U U U U U U U U U U U U U
Figura 1-4. Los cuatro tipos de nucleótidos pueden experimentar una polimerización espontánea con pérdida de agua. El resultado es una mezcla de polinucleótidos de longitud y orden aleatorios.
Figura 1-5. Los nucleótidos muestran preferencias al formar parejas: A con U, C con G, mediante la formación de puentes de hidrógeno. De esta forma, un polinucleótido sirve de molde para un polinu-
laces covalentes y enlaces entre nucleótidos. Por ejemplo, un RNA especializado puede catalizar el corte de otro RNA o de sí mismo en una secuencia determinada. Cada una de las reacciones químicas que cataliza el RNA catalítico depende de una conformación o agrupamiento específico de los átomos que constituyen su superficie, lo cual hace que uno o varios nucleótidos sean altamente reactivos. Estas reacciones catalíticas pudieron tener una importan- cia fundamental en la “sopa prebiótica”. En efecto, supóngase que una molécula de RNA es capaz de catalizar el proceso de polimerización a partir de una molécula cualquiera de RNA como molde. Esta molécula catalítica hará copias de sí misma, pero también de cualquier otro RNA que se encuen- tre en la proximidad, las cuales pueden ayudar, a su vez, a la reproducción o supervivencia de la primera. De esta manera, la especialización de diferentes RNA constituiría un sistema efi ciente capaz de copiarse y de sobrevivir (fig. 1-8).
Transferencia de información desde
los polinucleótidos a los polipéptidos
Se supone que hace entre 3.5 y 4 mil millones de años co- menzó la evolución de las moléculas automultiplicadoras o autorreplicantes de RNA. De la misma manera que ahora los organismos compiten para sobrevivir, sistemas compuestos de varios tipos de RNA, como los de la figura 1-9, competían por los precursores que les servían para automultiplicarse. Su capacidad para sobrevivir dependía de la exactitud y eficiencia de la copia, así como de la estabilidad de la misma. Sin embargo, aunque las moléculas de RNA tienen la aptitud de codificar y mantener información con sólo cuatro nucleótidos o bases, no son muy capaces de catalizar reacciones complejas. Los polipéptidos, que están compuestos por más aminoácidos, son mucho más capaces de crear puntos activos catalíticos por medio del plegamiento de sus cadenas. Incluso algunos polipéptidos creados al azar uniendo ciertos aminoá- cidos pueden mostrar capacidades catalíticas superiores a las del mismo RNA para la duplicación de éste. Por consiguiente, cualquier sistema de RNA con un polipéptido catalítico habría tenido mayores probabilidades de sobrevivir y, de igual manera, un polinucleótido capaz de estimular la síntesis de un polipéptido catalítico habría pre- sentado grandes ventajas sobre los demás. ¿Cómo pudieron los polinucleótidos ejecutar tal tarea? ¿Cómo pudo la información codificada en sus secuencias especificar el orden de diferentes tipos de polímeros? Se sabe que los polinucleótidos pueden actuar como catalizadores para unir diferentes aminoácidos. En la actualidad, la síntesis de proteínas se lleva a cabo mediante un conjunto de sistemas en los que desempeñan un papel importante tipos distintos de RNA. Sin embargo, en el proceso intervienen también diferentes proteínas previamente sintetizadas.
En resumen, la síntesis de proteínas tiene lugar de la forma siguiente:
- una molécula de RNA contiene la información genética para sintetizar un determinado polipéptido, información que está codificada
- otras moléculas de RNA actúan como portadoras al unirse a un tipo determinado de aminoácido.
Estos dos tipos de moléculas de RNA tienen secuencias complementarias, por lo que las instrucciones que están contenidas en la cadena que posee el código se transfiere al RNA que añade el aminoácido a la cadena polipeptídica en formación. Probablemente los precursores de estos dos tipos de moléculas de RNA dirigieron la primera síntesis de pro- teínas sin la ayuda de otras proteínas. En la actualidad, estos procesos de ensamble de aminoá- cidos tienen lugar en la superficie de los ribosomas, partículas complejas constituidas por varias moléculas de RNA de un nuevo tipo y por lo menos de 50 proteínas. Este RNA tiene un papel catalítico fundamental y constituye el 60% del
RNA catalítico que une nucleótidos para formar:
RNA catalítico que une nucleótidos para formar una nueva molécula con la misma secuencia y forma
catálisis
replicación
una copia de sí mismo
una copia de otro RNA
una copia de otro RNA
Figura 1-8.
ribosoma. Esto destaca, al menos desde un punto de vista evolutivo, la importancia del RNA en la primitiva síntesis dirigida de proteínas. De esta forma, el RNA fue capaz de ir creando herra- mientas —en forma de proteínas— para lograr una síntesis cada vez más eficiente de sí mismo y de nuevas moléculas de polinucleótidos y de polipéptidos. La síntesis de proteínas bajo la dirección de un RNA requirió la evolución de un código, el código genético , por medio del cual determinadas secuencias de nucleótidos especifican el orden en que se debe acomodar los aminoácidos que forman
la proteína. Este código está formado por conjuntos de tres nucleótidos o “tripletes”, cada uno de los cuales corresponde a un determinado aminoácido. Este código parece haber sido seleccionado aleatoriamente y está presente en todas las formas de vida, lo que hace pensar en que todas las células actuales descienden de una misma línea celular primitiva que surgió del perfeccionamiento del mecanismo de la síntesis de proteínas. Una vez que la evolución de los ácidos nucleicos llegó al punto de disponer de estas moléculas proteicas catalíticas, llamadas enzimas, la proliferación del sistema replicante se
RNA catalítico
RNA catalítico
RNA codificado molde para una proteína
RNA transportador de aminoácidos
RNA transportador de aminoácidos
RNA codificado molde para una proteína
Proteí na naciente
una copia de otro RNA
una copia de otro RNA
Figura 1-9.
también se pueden copiar algunas secuencias mutadas, pero con una eficacia menor. Unos químicos de la Universidad de Friburgo crearon paralelamente otro sistema automultiplicador. El equipo de G. van Kiedrovski estudió la reproducción competitiva de cuatro pequeños ácidos nucleicos a partir de dos oligonucleótidos. Dos de estos ácidos nucleicos dirigen la formación de pro- ductos autocomplementarios; los otros dos, la de productos complementarios (véase el lado izquierdo del esquema (BA: verificar a qué figura corresponde)). Al analizar las velocidades relativas de síntesis de los cuatro ácidos nucleicos establecieron que su copia es un sistema autocatalítico, hasta en el caso más ordinario de una simple complementariedad de los fragmentos de ácidos nucleicos con la hebra modelo, es decir, independi- entemente de la secuencia que se duplica. ¿Resuelven estos dos modelos el problema de la du- plicación del RNA prebiótico? El primero propone un mecanismo sencillo para la multiplicación del DNA, y el segundo muestra que la duplicación del DNA es un proceso autocatalítico cualquiera que sea la secuencia de DNA. Hay que señalar que en los dos casos, los oligonucleótidos utilizados están activados, es decir, eran más reactivos que los nucleótidos naturales, y que se añadieron agentes acopladores en el momento apropiado: los químicos ayudan a la automul- tiplicación. Por otra parte, el ácido nucleico que se estudió en ambos casos es DNA, que es más fácil de sintetizar que el
RNA, aunque se supone que este último apareció primero. Además, se sintetizaron los oligonucleótidos y, luego, se añadieron al medio reactivo, aunque se habrían podido for- mar de modo natural gracias a la acción catalítica de algunas rocas como la montmorillonita. Por paradójico que parezca, no se tiene ninguna idea de la forma en que surgieron los nucleótidos mismos. Según J. Ferris, si procesos como los descritos aquí su- cedieron para dar origen a la vida en la Tierra, estos nuevos resultados prueban sobre todo que el problema de la duplicación de los ácidos nucleicos fuera de una célula tiene solución, pero también que estaban presentes otros catalizadores distintos del RNA. Sin embargo, es posible imaginar que fragmentos de ácidos nucleicos, formados por catálisis mineral, se condensaron a veces en fragmentos más grandes. De manera gradual habrían adquirido las características necesarias para hacer vivir el mundo del RNA. Las primeras células vivas habrían aparecido al final de esta evolución.
Formación de la membrana
Es probable que uno de los acontecimientos cruciales en la evolución desde los sistemas automultiplicadores hasta la primera célula haya sido la formación de una membrana. La necesidad de mantener una alta concentración de proteínas
C C G C G G
CGG CCG
C C G
COMPLEMENTARIEDAD CRUZADA
AUTOCOMPLEMENTARIEDAD
REPLICACIÓN BASADA EN UNA CATÁLISIS CRUZADA DE OLIGONUCLEÓTIDOS, DIRIGIDA POR UNA HEBRA MODELO
C G G
C C G
C G G C G G C C G C C G C G G C G G
C G G C G G C C G C C G
C C G C C G
C C G C C G C G G C G G
C C G C G G C C G C G G C G G C C G
C G G C C G C C G C C G
C C G C G G
CICLO DE AMPLIFICACIÓN POR UNA TRIPLE HÉLICE doble hélice inicial (Crick y Watson)
(emparejamiento de Hoogsteen) 2
triple hélice agente de condensación
disociación
2
copia de la doblehélice inicial agente de condensación
C G G C C G C G G C C G
C GG
Figura 1-10B.
en las proximidades de un sistema automultiplicador de RNA y de evitar que otros sistemas vecinos se beneficiaran con las propiedades catalíticas de estas proteínas (fig. 1-11) originó el paso siguiente de la evolución: aislar el sistema de RNA automultiplicador junto con sus proteínas catalíticas mediante una membrana para que sólo el RNA que las generaba las pudiera utilizar. Existen unas moléculas, llamadas anfipáticas , que con- tienen una parte hidrófila (soluble en agua) y una parte lipó- fila (insoluble en agua) que se prestan especialmente para la formación de tales membranas. Estas moléculas se alinean en la interfaz aceite—agua para formar una vesícula monocapa (fig. 1-12 A ); cuando se colocan en agua se unen entre sí de manera espontánea y forman unas pequeñas vesículas cerradas bicapa por lo que el contenido acuoso queda aislado del medio exterior (fig. 1-12 B ). A las células de hoy las rodean membranas que están formadas, casi del todo, por fosfolípidos que adoptan dicha configuración de bicapas, cuyo ancho mide unos 5 nm y que se ven a la perfección con ayuda del microscopio electrónico.
Estas membranas aíslan las células del medio externo hostil y limitan el espacio ocupado por la célula. Se ignora en qué momento de la evolución se formó la primera célula. Puede haberse originado en el momento en que las primeras moléculas de fosfolípidos rodearon un
SIN MEMBRANA
Enzima primitiva
En una mezcla de RNA autoreplicantes, un RNA capaz de producir una proteína útil comparte esta proteína con otras especies de RNA competidoras
punto activado
ciclo replicante
CON MEMBRANA
Enzima primitiva membrana
Cuando el RNA y la proteína catalítica están encerrados en una membrana lipídica, el sistema aprovecha la enzima en exclusiva.
Figura 1-11.
Fosfolípidos
monocapa de fosfolípidos
AGUA
ACEITE
Figura 1-12 A****. Las moléculas anfipáticas como los fosfolípi- dos forman de modo espontáneo una monocapa en una interfaz aceite-agua.
AGUA
AGUA
AGUA
ACEITE
Figura 1-12B. Los fosfolípidos, cuando se dispersan en agua, forman espontáneamente vesículas monocapa en la interfaz aceite-agua o vesículas bicapa con las cabezas hidró- fi las enfrentando al agua.
espontánea y evolucione para adaptarse a un nuevo azúcar como fuente de carbohidratos, o bien, que se haga resistente a determinado antibiótico. Las bacterias se clasifican en dos grandes grupos de acuerdo con su hábitat (fig. 1-14):
- las eubacterias, que viven en los ambientes normales (suelo, agua, plantas y otros organismos superiores)
- las archaebacterias, que se han adaptado a vivir en condi- ciones sumamente adversas (fondo del mar, fuentes ter- males, zonas ricas en gases sulfurosos, etc.)
A pesar de su relativa simplicidad (o quizás por ello), las bacterias se alimentan con todo tipo de sustancias: azúcares sencillos o complejos, grasas, proteínas, hidrocarburos, poli- sacáridos, etc. Algunas incluso pueden aprovechar el CO (^2) como fuente de carbono y el N 2 como fuente de nitrógeno. En la actualidad son, sin duda alguna, los habitantes más abundantes del planeta Tierra, han sobrevivido durante más tiempo y probablemente son los que sobrevivirán más.
Reacciones metabólicas
Una bacteria que crece en un medio salino glucosado y que es su única fuente de carbono lleva a cabo un enorme número de reacciones químicas porque utiliza la glucosa como fuente de energía, pero también construye a partir de este azúcar todas las sustancias químicas que necesita para su desarrollo y mul-
tiplicación. Todas estas reacciones químicas están vinculadas entre sí a lo largo de la llamada cadena metabólica. En la célula primitiva, esta cadena metabólica no tenía excesiva importancia, ya que la célula extraía las pocas molécu- las necesarias para su subsistencia de la “sopa prebiótica”. Sin embargo, al agotarse estos recursos naturales, los organismos que tenían ventaja eran los que habían perfeccionado sistemas enzimáticos capaces de crear las moléculas necesarias. Por eso existe la opinión de que la cadena metabólica aumentó en complejidad hasta llegar a la presente en los organismos actuales. Si se plantea la hipótesis de que la cadena metabólica evolucionó mediante sucesivos añadidos de pasos enzimáticos, está claro que las reacciones más antiguas deben estar más próximas al “centro del tronco” metabólico, que es donde se sitúan las reacciones más vitales, y, además, todas las células vivientes las deben compartir. Este tronco metabólico está ocupado por la glucólisis , proceso mediante el cual la glucosa se degrada en forma anaerobia (en ausencia de oxígeno, dado que la atmósfera primitiva no contenía oxígeno). La glucólisis se verifica virtualmente en todas las células y produce ATP (trifosfato de adenosina) , la fuente más versátil de energía que utilizan todas las células. Junto a estas reacciones de los fosfatos de azúcares se ejecutan cientos de otras reacciones: mediante algunas se sintetizan pequeñas moléculas que luego son utilizadas para generar moléculas más complejas, otras son empleadas para degradar grandes moléculas, que llegan por medio de los alimentos, en moléculas más pequeñas. Estas reacciones
PROCARIOTA ANCESTRAL
ARCHAEBACTERIA
EUBACTERIA
bacterias anaeróbicas capaces de vivir a altas temperaturas (p.ej., bacterias sulfurosas)
bacterias capaces de vivir con altas concentraciones de sal (bacterias halófilas)
bacterias que reducen el metano a CO2 (bacterias metanógenas)
bacterias grampositivas bacterias verdes fotosintéticas cianobacterias (algas azul-verdosas)
bacterias púrpura fotosintéticas bacterias gramnegativas espiroquetas
Figura 1-14. Evolución de las células procariotas.
metabólicas son comunes a todos los seres vivos. Evidente- mente, existen diferencias: algunas especies no son capaces de producir determinadas sustancias (p. ej., el hombre y otros animales superiores no pueden fabricar el aminoácido lisina, que sintetiza cualquier bacteria), pero en general existe una amplia concordancia de los procesos químicos de todos los seres vivos, prueba de una evolución a partir de una célula primitiva común.
Fotosíntesis
Si los primeros procesos metabólicos evolucionaron para llenar el vacío de compuestos orgánicos de la “sopa prebiótica”, cabe preguntarse qué pasó cuando se agotaron estos compuestos. Está claro que los organismos capaces de utilizar como fuente de carbono el CO 2 y como fuente de nitrógeno el N 2 atmos- férico tenían más ventajas de sobrevivir. Sin embargo, aunque son muy abundantes en la atmósfera, ambos compuestos son muy estables, por lo cual se requiere mucha energía y un número elevado de reacciones para convertirlos en productos sencillos, como azúcares o aminoácidos. En el caso de la captación del CO 2 se creó un mecanismo, la fotosíntesis, mediante el cual un pigmento, la clorofila, capta la energía radiante procedente del Sol. La célula utiliza posteriormente esta energía para sintetizar las moléculas que necesita. Una de las primeras reacciones que induce la luz solar fue, quizá, la fosforilación de los nucleótidos para originar el ATP, comodín energético de las células. Otra reacción probable fue la reducción del CO 2 y del N 2 para permitir su incorporación a moléculas orgánicas. La reducción —incor- poración de electrones— se efectuó al inicio a partir del SH 2 y luz solar, y después a partir de H 2 O y luz solar. Así empezó la producción en gran escala del O 2 , como producto secundario de la fotosíntesis. Las cianobacterias (también llamadas algas azules) son microorganismos actuales capaces de fijar el CO 2 y el N 2 de la atmósfera, y quizás sean los organismos más auto- suficientes que existan ya que, en principio, pueden vivir sólo de agua y luz solar. Junto con otros organismos que tienen estas propiedades, las cianobacterias, que se mantienen constantes desde hace miles de millones de año, crearon las condiciones idóneas para que otros organismos pudieran evolucionar con mayor facilidad.
Respiración
Por ser el oxígeno una sustancia altamente reactiva, debió resul- tar tóxica para los organismos primitivos, de la misma manera que actualmente es tóxica para los microorganismos anaero- bios. Por esta razón y para aprovechar su energía química, las células primitivas aprendieron a utilizar este elemento para
mejorar los procesos de degradación de las moléculas alimen- ticias. La glucosa, en ausencia de oxígeno, se descompone en ácido láctico o etanol (y produce dos moléculas de ATP), pero en presencia de oxígeno se degrada a CO 2 y H 2 O y produce 36 moléculas de ATP. La célula utiliza la energía producida en este proceso, denominado respiración, para su metabolismo y producción de nuevas moléculas de la misma manera que con la fotosíntesis se obtiene ATP en los organismos fotosinté- ticos. La mayor parte de los organismos, incluso casi todos los procariotas, emplean hoy el proceso de la respiración.
Glucólisis
La ruta glucolítica (o vía de Embden—Meyerhoff) consiste en la degradación anaeróbica de la glucosa a lactato. Para muchos tejidos, la glucólisis representa una ruta metabólica de urgen- cia, capaz de producir dos moles de ATP a partir de una mol de glucosa en ausencia de oxígeno molecular (fig. 1-15).
La célula eucariota
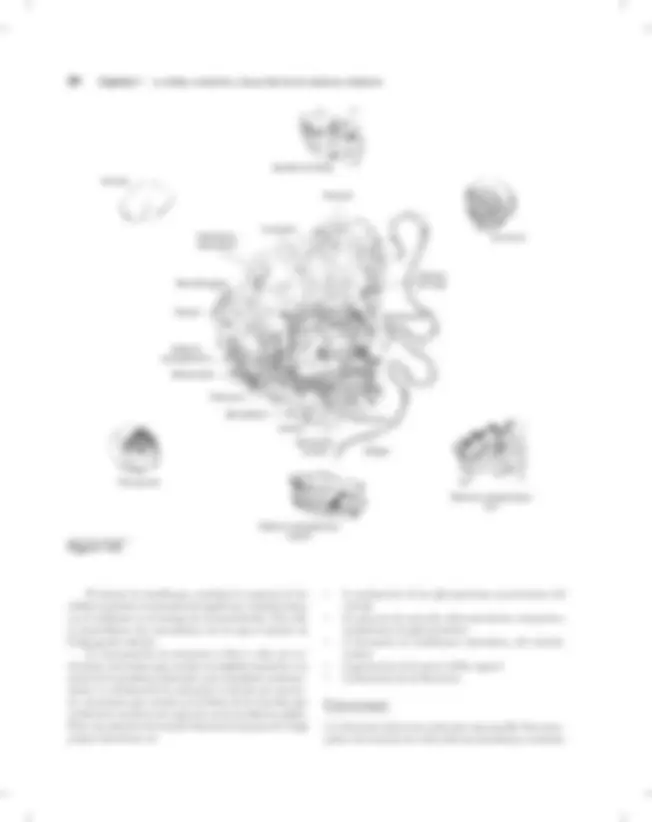
La célula eucariota tiene en la actualidad una estructura orga- nizada. A medida que la atmósfera se enriquecía con oxígeno, pereció una parte de las células primitivas que no pudieron adaptarse a estas nuevas condiciones. Otras desarrollaron una capacidad para respirar, o bien, tuvieron que ocultarse en lugares donde el oxígeno estaba ausente para conservar su condición de anaerobias. Sin embargo, una tercera clase descubrió que si se unía en simbiosis con una célula aerobia podía sobrevivir y desarrollarse de un forma mucho más rica. Ésta es la hipótesis más viable para la organización metabólica de las células eucariotas de hoy día que se estudia en detalle más adelante (cuadro 1-1). Por definición y en contraste con las células procari- otas, las células eucariotas tienen una estructura organizada y disponen de un cierto número de organelos. En particular, tienen un núcleo, separado del resto de la célula mediante
CH (^3)
CH 2 OH
H
O
O O C
2 OH OH
OH
OH OH
C
d-Glucosa-α l-Lactato
2ADP3–^ 2ADP4– +2Pi2–
Figura 1-15. Esquema simplifi cado de la glucólisis.
- mitocondrias y cloroplastos
- retículo endoplásmico liso (fino) y retículo endoplásmico rugoso (grueso)
- aparato de Golgi
- ribosomas
- lisosomas y peroxisomas
- citoesqueleto
- vacuolas
- flagelos
Una membrana más o menos organizada rodea la cé- lula eucariota, como las procariotas; dicha membrana está constituida fundamentalmente por fosfolípidos y proteínas específicas que tienen diferentes funciones. Al parecer, las mitocondrias evolucionaron a partir de algún organismo procariota que se unió en simbiosis con algún otro organismo primitivo anaerobio. En efecto, en muchos aspectos, las mitocondrias se parecen a los organismos procariotas:
- tienen un tamaño y forma parecidos
- se multiplican por división
- contienen su propio DNA
- el proceso de respiración se efectúa exclusivamente en ellas
- muchas bacterias actuales respiran igual que las mito- condrias
Mediante esta simbiosis, primitivos eucariotas anaerobios podrían haber sobrevivido en un ambiente cada vez más rico en oxígeno, utilizando la capacidad del procariota asociado para producir energía mediante el consumo de oxígeno at- mosférico. La adquisición de mitocondrias tuvo, sin duda, im- portantes repercusiones. Al ocuparse sólo de la respiración, permitió que la membrana se especializara en otras funciones, a diferencia de lo que ocurre en los procariotas; en éstos se debe conservar un gradiente importante de H+^ en la membrana plasmática para producir ATP. Por consiguiente, la membrana
eucariota pudo crear canales iónicos y otros organelos —re- ceptores— especializados en el intercambio de señales. Los cloroplastos llevan a cabo la fotosíntesis de la misma manera que las cianobacterias: absorben la luz solar con la clo- rofila que existe en sus membranas. También los cloroplastos manifi estan similitudes notables con las cianobacterias:
- son similares en forma y tamaño
- las membranas en las que se encuentra la clorofila están dispuestas en capas
- se reproducen por división
- contienen DNA con secuencias muy parecidas al DNA bacteriano
Todo ello apunta a que los cloroplastos evolucionaron a partir de las cianobacterias que se unieron en simbiosis a alguna célula eucariota primitiva. Este tipo de simbiosis es relativamente frecuente y, en la actualidad, se conocen células eucariotas que tienen auténticas cianobacterias en su interior. La teoría de la evolución de los eucariotas a partir de la hipó- tesis simbiótica se ilustra en la figura 1-17.
Teoría actual de la
endosimbiosis como origen
de la célula eucariota
La célula eucariota se distingue también por una red pro- teica, el citoesqueleto, que sitúa y desplaza los organelos en la célula. Todos los vegetales, los hongos y los animales son eucariotas. Su DNA está en uno de los organelos, el núcleo. Además del retículo endoplásmico y el complejo de Golgi, que intervienen en la síntesis de ciertas proteínas, los otros dos compartimientos que desempeñan un papel sustancial ya que suministran energía a la célula son las mitocondrias
Archaebacteria Otras eubacterias
Bacterias púrpuras sufurosas
Cianobacterias Plantas Hongos Animales
Procariota ancestral
Procariota ancestral anaerobio
Eucariota ancestral
Mitocondrias
Cloroplastos
Figura 1-17. Teoría simbiótica de la evolución de los eucariotas.
(del griego mitos , filamento, y khondros , grano), las cuales se ocupan de la respiración celular, y los plastos de los vegetales, donde tiene lugar la fotosíntesis o conversión de la energía luminosa en energía química. Algunos organelos pueden desaparecer y volverse a formar. Es el caso del núcleo durante la división celular, del aparato de Golgi y del retículo. Pero las mitocondrias y los plastos proceden siempre de la división de mitocondrias y plastos preexistentes. A finales del siglo XIX, esta par- ticularidad hacía pensar ya en bacterias que se dividían por bipartición en el citoplasma. Las mitocondrias y los plastos eran organismos vivos, instalados en simbiosis en la célula. En 1883, el investigador alemán A. Schimper observó : “Si se establece definitivamente que los plastos nunca se forman de novo en la célula huevo, entonces su situación en la célula donde se en- cuentran recuerda la de los simbiontes. Es posible que una planta verde no sea otra cosa que la unión entre un organismo incoloro y un microbio que posee los pigmentos clorofílicos.” Otros autores le pisaron los talones a principios del siglo XX, pero toparon, lamentablemente, con la incredulidad de sus colegas. Ocurrió como con la deriva de los continentes de Alfred Wegener: los argumentos propuestos no bastaron para afianzar la intuición. En particular, fracasaron todos los intentos de aislamiento y cultivo in vitro del microbio verde. Varios autores propusieron para estos organelos un origen autógeno a partir de repliegues de la membrana plasmática. Los progresos de la biología iban a brindar otros argumentos a la idea según la cual las mito- condrias y los plastos derivan de bacterias. La microscopia electrónica demostró que los organelos de los eucariotas están limitados por una membrana lipídica simple, salvo tres de ellos: el núcleo, rodeado por membranas provistas de poros que lo ponen en continuidad con el citoplas- ma, y los plastos y las mitocondrias, separados del citoplasma por dos membranas continuas. Además, la organización de la estructura de estos dos últimos organelos evoca la de ciertas bacterias. Por ejemplo, las membranas fotosintéticas o tilac- oides de los plastos de ciertas algas (las algas rojas) evocan en gran medida a los tilacoides de bacterias fotosintéticas, las cianobacterias. Los adelantos de la bioquímica pusieron de manifiesto otros parecidos sorprendentes. Los lípidos de las membranas que rodean a las mitocondrias existen en ciertas bacterias, pero no en otras partes de la célula eucariota. En cuanto a los lípidos característicos de las membranas de los plastos, se les encuentra en las cianobacterias. Además, las mitocondrias y los plastos desempeñan ciertas funciones metabólicas que efectúan también ciertos procariotas libres. La fotosíntesis se consuma de manera casi idéntica en las ci- anobacterias y los plastos. Las reacciones de oxidorreducción ligadas a la actividad energética de las mitocondrias y de los plastos requieren la intervención de cadenas de transferencia de electrones membranosas parecidas a las de los procariotas: colaboran en ellas unas complejas proteínas homólogas, como los citocromos.
Pero el argumento principal fue el descubrimiento de un genoma en los plastos en 1962 y luego en las mitocondrias al año siguiente. Este DNA se parece al de los procariotas: no está aislado dentro del organelo y se presenta en forma de varias copias idénticas de una molécula circular. El DNA se duplica y se transfiere durante la bipartición del organelo como en el caso de los procariotas libres. En las mitocondrias y los plastos existen ribosomas que sintetizan proteínas y son más pequeños que los del citoplasma de la célula. De hecho, tienen el tamaño y la composición del RNA de los ribosomas de los procariotas. Por todas estas razones, la hipótesis del origen simbiótico de las mitocondrias y de los plastos halló adeptos en los años 1970, como Lynn Margulis, de la Universidad de Amherst (Massachusetts), y Sarah P. Gibbs, de la Universidad McGill en Montreal. La biología molecular subrayó todavía más el parentesco entre estos organelos y los procariotas. Es el caso, por ejemplo, de las zonas promotoras de los genes, es decir, de las secuencias reconocidas para proteínas que aseguran su transcripción en RNA. Se descubrieron actualmente genes homólogos (que tienen el mismo origen evolutivo) de genes bacterianos en el genoma de las mitocondrias y de los plas- tos. Sería sorprendente encontrar bacterias en el citoplasma de eucariotas. En realidad, semejante modo de vida no es raro entre los eucariotas: se le designa endosimbiosis. Este modo de vida es el de ciertos microorganismos patógenos, como la bacteria causante de la listerosis o de la fiebre tifoi- dea. Pero también, el de numerosos animales unicelulares, los protozoarios, que son colonizados por varias bacterias. Kwang W. Jeon, de la Universidad de Tennessee, describió la aparición de una endosimbiosis entre las amebas, a raíz de una infección que diezmó un cultivo de laboratorio en 1966. Algunas amebas sobrevivieron a la presencia de bacterias en su citoplasma: veinte años más tarde, todavía contienen unas 42 000 bacterias por célula. Pero hay más: esta asociación se ha vuelto forzosa. Las amebas ya no pueden sobrevivir si se da muerte a sus compañeros citoplasmáticos aumentando la temperatura. Otro ejemplo clásico: las plantas de la familia de las leguminosas (trébol, guisante) poseen en sus raíces unos nódulos cuyas células albergan bacterias del género Rhizobium. Estas bacterias, llegadas del suelo, ayudan a la planta a alimentarse fijando el nitrógeno atmosférico. Estos microorganismos unicelulares están rodeados siempre por dos membranas: la membrana interna, que es la de la bacteria, y la membrana externa procedente de la membrana plasmática del hospedero, que rodea a la bacteria durante su penetración en la célula. No obstante, el origen de la membrana externa de las mitocondrias y los plastos es objeto de controversia (en el caso de los plastos, contiene lípidos de tipo procariota y sería de origen mixto). ¿Cuántos sucesos de endosimbiosis han tenido lugar en el curso de la evolución? ¿Cuáles son los procariotas actuales más próximos a los que viven en endosimbiosis? La biología
con respecto a los plastos de algas rojas. En el transcurso de la evolución, el alga roja asimilada habría perdido sus pigmentos rojos y adquirido la clorofila c , con lo cual se convirtió en un plasto pardo característico. Otros organismos pudieron adquirir plastos por en- dosimbiosis secundaria. Esto resuelve la vieja paradoja de las Euglena. Los manuales presentan a estos protozoarios como mitad animales (ya que las Euglena son flagelados y pueden fagocitar presas o restos) y mitad vegetales (pues poseen plastos con los mismos pigmentos que los de los vegetales verdes). En realidad, como se demuestra al comparar los genes de los plastos (fig. 1-19), se trata efectivamente de plastos de vegetales verdes. No obstante, las Euglena son eucariotas primitivos muy cercanas a los tripanosomas. Pudieron adquirir sus plastos a partir de un alga verde, presa o endosimbionte. Numerosos ejemplos actuales ponen de manifiesto fases intermedias en las endosimbiosis, tanto primarias como se- cundarias. De hecho, el aprendizaje de la vida en común tiene numerosas exigencias. El encuentro de los participantes es la primera fase, que no necesariamente representa ventaja para los dos. Las mitocondrias derivan, tal vez, de bacterias pató- genas endosimbióticas, como las bacterias actuales causantes de la listerosis o de la fiebre tifoidea. Por lo que se refiere a las cianobacterias, fueron tal vez presas antes de convertirse en plastos. En una segunda fase, el vínculo puede volverse más duradero. Puede significar una ventaja para un eucariota cazador de cianobacterias aprovechar algún tiempo la foto- síntesis de su presa antes de digerirla, o bien, criarla en su citoplasma. Cuando la relación es permanente, es necesario un control recíproco sobre la multiplicación de cada socio
para la supervivencia del organismo quimera formado. Queda abierto entonces el camino para una evolución conjunta (o coevolución) de los socios, y pueden establecerse intercambios recíprocos. Para el endosimbionte, la vida en el interior de la célula es en extremo confortable; es el hospedero el que soporta las agresiones del medio externo. Este hábitat protegido explica la desaparición de las envolturas protectoras; en los plastos con dos membranas ya no quedan huellas de la pared de las cianobacterias. La regresión de la pared también ha sido descrita en algas eucariotas unicelulares endosimbióticas ac- tuales. Pero la evolución regresiva más curiosa es, en verdad, la reducción del genoma de los organelos con respecto a las formas libres en el transcurso de la evolución (véase cuadro 1-2). Ya desapareció el núcleo del alga roja que asimilaron las algas heterocontas de plastos pardos; el tamaño del genoma de tipo procariota que está contenido en los organelos es entre diez y cien veces más pequeño que el de los procariotas libres. Por supuesto, ya desaparecieron todos los genes necesarios para orientarse, desplazarse y protegerse en el medio exterior,
Figura 1-18.
Mitocondrias de animales Mitocondrias de hongos Vaca^ Erizo^
Mitocondrias de plantas Cachliabalus Maíz Aspergillus
Padospora
Acanthamoeba (protozoos) Pylalello (alga parda)
Tabaco Anagra Marchantia Prototheca(hopatica) (alga verde) Chondrus (alga roja)
Rhodospirillum rubrum
Mitocondrias
Protobacterias
Escherichia coli
Synechocystis Anabaena
Antilharmnion (alga roja) Odontella (diatomea)
Euglena Marchantia (hepatica)
Guisante
Maíz
Cianobacterias
Plastos
Figura 1-19.
pero otros siguen existiendo: simplemente fueron transferidos al núcleo de la célula hospedera mediante un proceso que se empieza a entrever gracias al estudio reciente de Brennicke y colaboradores (cuadro 1-2). En el curso de la evolución, muchos genes procedentes de los plastos y las mitocondrias migraron hacia el núcleo de la célula que los albergaba; aunque son nucleares, estos genes tienen, entonces, un origen evolutivo procariota. Estas transferencias sellan la asociación y son la razón de que no se puedan cultivar de modo aislado los organelos, los cuales se volvieron por completo dependientes de su hospedero. La simbiosis se volvió forzosa y los socios forman un nuevo or- ganismo quimérico. Unas regulaciones acoplan la expresión de ambos genomas, y participan en el buen funcionamiento de la nueva unidad fisiológica. La célula eucariota esconde tal vez otras endosimbiosis. Es el caso de los peroxisomas, unos organelos que intervienen en ciertas reacciones metabólicas como la fotorrespi- ración y las peroxidaciones. Estos organelos están limitados por una simple membrana, carecen de DNA y aparecen siempre por bipartición. Algunos científicos los consideran como endosim- biontes cuyo genoma fue transferido del todo al núcleo. Por tanto, la endosimbiosis influyó profundamente en la evolución de los eucariotas y les facilitó la consecución de las potencialidades metabólicas de los procariotas. Este fenómeno acompañó varias veces la fundación de linajes evolutivos en los eucariotas (fig. 1-20). Se complementan las potenciali- dades morfogenéticas de los eucariotas (citoesqueleto, paso al estado pluricelular, entre otros) y la ingeniería enzimática (fijación del carbono o del nitrógeno, respiración, etc.) de los procariotas, los cuales, aunque a menudo asimilados, nunca albergan simbiontes. Los sucesos de endosimbiosis parecen haber sido bas- tante numerosos, en ocasiones sucesivos, y originaron los plastos de cuatro membranas. En la actualidad, ciertas algas unicelulares, las criptofitas y las heterocontas, cuyo plasto de cuatro membranas deriva, recuérdese, de una endosimbiosis secundaria, viven en simbiosis en el citoplasma de dinoflage- lados que perdieron sus propios plastos. Se trata entonces de tres endosimbiosis sucesivas.
Estructura celular
Las células son estructuras con muy buena organización en su interior; están constituidas por diferentes organelos, y cada uno de ellos cumple funciones diferentes. Sin embargo, todas las células eucariotas, que son las de todos los seres vivos con excepción de las bacterias, cuyas células son mucho más sencil- las, comparten un plan general de organización:
- Una membrana que determina su individualidad
- Un núcleo que contiene el material genético y ejerce el control de la célula
Cuadro 1-2. Reducción del endosimbionte
Organismo Número total de genes
Tamaño del genoma (en pares de bases)
Cianobacterias libres > 5 000 de 1 a 10 millones Porphyra Alga roja 210 183 000 Cryptomonas Criptofi ta
140 112 000
Oryza (arroz) 110 114 000 Nicotiana (tabaco) 113 131 000
Tamaño de los genomas procariotas contenidos en los plastos de distintos grupos de vegetales comparado con las cianobacterias libres. En el transcurso de la evolución, numerosos genes de los plastos se transfi rieron al núcleo del hospedero eucariota.
Adaptado de Douglas SE, et al. Nature, 1991;350:148.
protistos
clorobiontes
heterocontas
rodobiontes
alveolobiontes
plantas terrrestres
algas verdes
rizópodos criptófitas (núcleo) oomicetos (seudohongos)
criptófitas (nucleomorfo) algas rojas
diatomeas y algas pardas
esporozoos (plasmodium) dinoflagelados ciliados (paramecios) distintos grupos de protozoos (amebas) tripanosomas
Figura 1-20. Antepasados comunes con los procariotas.