¡Descarga practica y más Ejercicios en PDF de Biología solo en Docsity!
Borja Figueirido, Paul Palmqvist y Juan A. Pérez-Claros
Introducción al Análisis Discriminante:
Evolución morfológica craneal en Ursidae
(Mammalia, Carnivora)
EEEEn esta práctica de la asignatura Paleontología y Evolución se ofrece
una introducción al Análisis Canónico Discriminante como una de las herramientas de clasificación –propias de la estadística multivariable– más utilizadas en el ámbito de la Paleobiología. Para el desarrollo de la práctica se hará uso de una base de datos craneométrica que incluye representantes de los diferentes géneros y especies de la familia Ursidae. Cuenta con un total de unas cuarenta variables métricas craneodentales tomadas en diferentes ejemplares de todas las especies vivientes de Ursidae: oso panda (Ailuropoda melanoleuca), oso polar (Ursus maritimus), oso perezoso (Ursus ursinus) oso andino (Tremarctos ornatus), oso pardo (Ursus arctos), oso negro americano (Ursus americanus), de oso negro asiático (Ursus thibetanus) y de oso malayo (Ursus malayanus). Asimismo, se dispone de estas variables en diferentes especímenes del oso de cara corta del Pleistoceno de Norteamérica, Arctodus simus. El objetivo principal es familiarizar al alumnado con la base teórica del análisis canónico discriminante, así como con su ejecución para la aplicación práctica. Para ello se ha elegido el programa SPSS v.19 – disponible en los recursos informáticos de la UMA– pues este software es probablemente el más usado para la práctica de la estadística multivariable en el ámbito de la Biología. Asimismo, se ilustrará el potencial del análisis discriminante para descifrar la paleoecología de las especies pretéritas. En consecuencia, el alumno tendrá que ser capaz de realizar e interpretar los análisis canónicos discriminantes y emanar conclusiones a discutir sobre la dieta de la especie extinta incluida en nuestra base de datos: el úrsido gigante del Pleistoceno de Norteamérica. Este último punto servirá para reforzar los contenidos de las clases teóricas sobre los métodos ecomorfológicos para descifrar la paleoautoecología de las especies extintas como herramienta de reconstrucción de la historia de la vida en la tierra.
El esquema a seguir en la práctica será el que se detalla a continuación:
- Introducción a la familia Ursidae y al orden Carnivora.
- Toma de variables en un espécimen mediante calibre digital.
- Breve descripción de la base de datos.
- Introducción teórica al análisis canónico discriminante (ACD).
- Interpretación del ACD: funciones y clasificación de grupos.
- Ejecución del ACD mediante SPSS v.19.
- Discusión y conclusiones.
Introducción al Análisis Discriminante:
Evolución morfológica craneal en Ursidae
(Mammalia, Carnivora)
Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
Borja Figueirido, Paul Palmqvist y Juan A. Pérez-Claros
La familia Ursidae, es una familia de mamíferos perteneciente al orden Carnivora (véase columna de la derecha) que evolucionó, a finales del Oligoceno e inicios del Mioceno, a partir de un grupo de cánidos hesperocioninos poco derivados, hace unos 20-25 Ma. Ahora bien, pese a que la historia evolutiva de los osos sea comparativamente más corta que la de otras familias de carnívoros, no es menos importante desde un punto de vista morfológico.
Sobre la familia Ursidae
La clasificación taxonómica de los úrsidos ha suscitado persistentes debates científicos.
Actualmente, diversos investigadores dividen la familia en cinco subfamilias: Hemicyoninae (�), Agriotheriinae (�), Tremarctinae, Ursinae y Ailuropodinae (véase cuadro 3.1).
Hoy día la familia Ursidae comprende únicamente ocho especies, distribuidas en cinco géneros que habitan en Norteamérica, Sudamérica, Europa y Asia (véase columna de la derecha de la página siguiente).
Rangos temporales para los géneros de osos actuales y extintos (barras horizontales rojas). Tomado de Krause et al. (2008; BMC Evol Biol, 8:
El orden Carnivora
CUADRO 3.1: Relaciones filogenéticas de Ursidae
<<
El orden Carnivora comprende 271 especies, que se reparten en 129 géneros y 13 familias. El carácter clave del orden es la presencia del llamado “par carnicero”, es decir, un cuarto premolar superior de cúspides cortantes que ocluye con el primer molar inferior (con cúspides también cortantes) en forma de tijera o cizalla. Los carnívoros presentan una amplia diversidad ecológica, con una distribución cosmopolita, se encuentran en hábitats muy diferentes (vg., desde desiertos o selvas hasta el Ártico o la alta montaña) y muestran una ecología trófica también muy variada, pues su dieta incluye desde altas cantidades de vegetales hasta insectos, carroña o carne de vertebrados.
Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
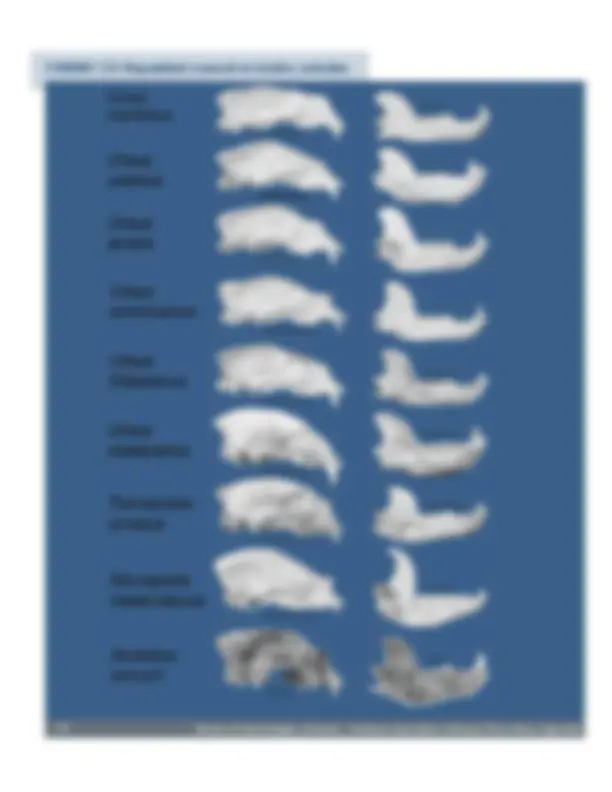
CUADRO 3.2: Disparidad craneal en úrsidos actuales
<<
Arctodus simus: el oso de cara corta y extremidades alargadas que nunca lo fue
Con una masa corporal estimada en 700-800 kg y una paleobiología un tanto enigmática, el oso de cara corta, Arctodus simus (Mammalia, Carnivora, Ursidae) ha sido el representante del orden Carnivora más grande que jamás ha hollado la tierra (véase columna de la derecha). Se han recuperado restos fósiles de este úrsido en más de cien localidades del Pleistoceno de Norteamérica, lo que evidencia que A. simus desempeñó probablemente un papel importante en los ecosistemas de esta época (véase columna de la derecha). Este oso se extinguió junto con la mayoría de la megafauna hace alrededor de unos12.000 años. La ecología trófica de A. simus ha sido objeto de debate y controversia por parte de la comunidad científica durante décadas. Así, por ejemplo, Kurtén (1967) sugirió que este oso fue un depredador activo, basando esta conclusión en algunos caracteres que, en su opinión, evidenciaban una anatomía del “tipo-félido” en comparación con otros osos actuales y extintos. Tales caracteres incluían un rostro corto y amplio, carniceras con una región cortante bien desarrollada, un cráneo muy profundo y de rostro acortado, mandíbulas poderosas y extremidades extremadamente alargadas. Siguiendo la hipótesis de Kurtén, muchos autores han asumido desde entonces que A. simus fue el carnívoro terrestre de hábitos depredadores con mayor porte que jamás ha existido. Sin embargo, otros autores intentaron desmitificar a A. simus basándose en las características craneodentales que esta especie comparte con su pariente filogenético actual, el oso de
anteojos de Sudamérica (Tremarctos ornatus ), especie cuya dieta es básicamente omnívora, aunque incorpora gran cantidad de materia vegetal en la misma. Estos autores sugirieron que A. simus fue básicamente un omnívoro, con una dieta que incorporaría grandes cantidades de recursos vegetales, aunque también enfatizaron su capacidad para fracturar hueso. No obstante, conviene reconocer que la similitud morfológica existente en el esqueleto craneodental de A. simus y T. ornatus probablemente refleje un patrón filogenético común y no una convergencia adaptativa. Subsecuentes avances en los análisis isotópicos del colágeno óseo y sus aplicaciones paleoecológicas, llevaron a otros autores a revitalizar la hipótesis que consideraba a Arctodus como la “gran fiera” del Pleistoceno de Norteamérica. Concretamente, basándose en sus altos valores de δ^15 N, sugirieron que se trataba de un carnívoro estricto, lo que corroboraría la hipótesis de Kurtén (1967). Por el contrario, Matheus (1995) concluyó, tras efectuar análisis biogeoquímicos que apuntaban en la misma dirección, que debido a la competencia con otros grandes predadores, A. simus fue probablemente un carroñero especializado más que un hipercarnívoro depredador (véase columna de la derecha; página siguiente). Bajo esta reconstrucción, las largas extremidades de A. simus le habrían permitido prospectar eficientemente una amplia área de campeo y su gran tamaño corporal podría haber sido utilizado como una estrategia disuasoria para otros carnívoros del Pleistoceno, facilitando así
Secuencia de
reconstrucción de
A. simus
Yacimientos con
restos del úrsido
gigante
<<4 Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
La base de datos a utilizar A. simus^ como predador.
Hipótesis de Kurtén
A. simus como carroñero.
Hipótesis de Matheus
A. simus como herbívoro.
Hipótesis de Emslie
La base de datos craneodental comprende 122 individuos pertenecientes a las 8 especies de úrsidos actuales que hemos comentado anteriormente. Además, se han incluido especímenes fósiles de A. simus. Concretamente, se han incluido fósiles de A. simus procedentes de diferentes localidades de Norteamérica (véase cuadro 3.3), como un cráneo con mandíbula asociada de Hay Springs, Sheridan County (Nebraska), un cráneo de Potter Creek Cave, Shasta County (California), un cráneo de Upper Cleary River Beds (Alaska), un cráneo de Ester Creek (Alaska), tres cráneos de Rancho La Brea (California), un cráneo con mandíbula asociada de Fulton County (Indiana), una mandíbula aislada de Arkalon (Kansas), y cuatro mandíbulas de Rancho La Brea (California).
El objetivo será evaluar si A. simus muestra claras convergencias morfológicas con los úrsidos hipercarnívoros actuales, tal como propuso Kurtén (1967), o si por el contrario fue un omnívoro de gran porte.
Todos los datos métricos utilizados en este estudio proceden de ejemplares depositados en las colecciones osteológicas de diferentes museos internacionales, como el American Museum of Natural History (AMNH; Nueva York, USA), el Los Angeles County Museum (LACM; California; USA), el Museum für Naturkunde (MFN; Berlín, Alemania), el Muséum National d'Histoire Naturelle (MNHN; París, Francia), el Natural History Museum (NHM; Londres, UK) y el Naturhistoriska Riksmuseet (Estocolmo).
CUADRO 3.3: Algunos especímenes de Arctodus
Nebraska
Indiana
Alaska
California
<<
Variables métricas al concurso de la estadística multivariable
Numerosos estudios ecomorfológicos previos han mostrado que los análisis comparativos de la anatomía esquelética en los grandes carnívoros fisípedos aportan información relevante sobre sus preferencias tróficas y de hábitat, así como en relación a su modo de locomoción y comportamiento en la caza.
Basándonos en estos estudios, se han elegido una serie de variables métricas en el esqueleto craneodental que permiten estimar los momentos de fuerza para la musculatura temporal y masetérica, la longitud de la fosa masetérica, la profundidad de la
rama horizontal, la longitud y anchura del hocico, la anchura del cráneo y las longitudes, tanto mesiodistales como bucolinguales, de cada elemento de la dentición superior e inferior (véase cuadro 3.4).
Tales variables medidas en la muestra de carnívoros actuales de dieta conocida y en los cráneos fósiles de A. simus serán las que se sometan al análisis canónico discriminante para descifrar la dieta del plantígrado extinto.
Téngase en cuenta que en esta práctica se espera que previamente al análisis discriminante se efectúe un análisis de componentes principales para explorar la ordenación de los especímenes en un espacio fenotípico.
CUADRO 3.4: Variables métricas
Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
Las partes de la carnicera inferior
Cuales son estas partes en la carnicera de un felino
<<
Si pensamos en la generalizar este razonamiento a más de dos variables, se obtienen funciones multidimensionales con el mismo contenido conceptual: D = λ 1 X 1 + λ 2 X 2 +….+ λpXp
Así, esta optimización debe conseguir el mayor número de individuos de un mismo grupo con altos valores D, y el mayor número de individuos del otro con bajos valores. Por tanto D, servirá de discriminante entre ambos grupos mucho mejor que X 1 y X 2 de forma independiente. De este modo, a un nuevo individuo del que disponemos de los valores de las variables X 1 y X 2 , bastaría con aplicarle la función D y, con un grado de error que se puede calcular, asignarle el grupo de pertenencia más probable. Resulta intuitivo pensar que esta función D será tanto más eficaz cuanta mayor separación entre los centros de gravedad de los
CUADRO 3.6: Lógica del análisis discriminante II
dos grupos (i.e., puntos blancos en el Cuadro 3.5 y 3.6) consiga. Así, la máxima separación en sus proyecciones obligaría a la función discriminante a ser paralela a la recta que une estos dos centros de gravedad. Sin embargo, como se puede observar en el Cuadro 3.6 la separación entre ambos grupos mediante esta función discriminante (recta roja) dista de la perfección, pues existe una zona verde en la cual aún existen individuos entremezclados. Para solventar este problema, se puede buscar un eje no paralelo bajo el criterio de que se consiga hacer mínimas las dispersiones de las distribuciones proyectadas (elipses amarilla y azul en el ejemplo) reduciendo así la zona entremezclada y sacrificando el criterio de máxima separación de los dos centros de gravedad. Por tanto, la solución óptima ha de pasar por una combinación de ambos criterios que constituye un importante principio del Análisis Discriminante.
<<9 Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
Para ello, se llega al consenso de maximizar el cociente dispersión entre grupos (o dispersión factorial) / cociente dispersión intragrupos (dispersión residual). Por tanto, cuanto mayor sea la separación de los centros de gravedad, a costa de una menor dispersión en las proyecciones de los individuos de cada grupo la discriminación que ejerza D será mejor. Esto equivale a encontrar unos valores λ 1 ….. λ p tales que maximicen el cociente:
∆^2 = (DA / DB)^2 / S^2 r
Donde DA y DB son los valores medios de la función discriminante aplicada a todos los individuos de los grupos A y B, respectivamente, y S^2 r es una estimación conjunta de la varianza residual de D para ambos grupos. La solución matemática de la maximización de este cociente (primera derivada = 0; segunda derivada <0) permite hallar los valores (λ 1 ; λ 2 … λp) que, multiplicados por las variables originales (X 1 , X 2 … Xp), configuran la función D buscada.
La raíz cuadrada de ∆^2 se denomina como distancia generalizada de Mahalanobis. Así, a mayores valores de ∆ se obtiene una menor superposición en el valor de D para los casos pertenecientes a grupos distintos y una menor posibilidad de una clasificación incorrecta de los mismos.
Si se calculan los valores medios que toma cada variable en cada uno de los grupos (A y B en nuestro caso), otra forma de expresar la distancia de Mahalanobis es:
Si disponemos de un nuevo individuo cuyo grupo de pertenencia desconocemos (por ejemplo, un ejemplar de una especie extinta de dieta desconocida) en el que se puede estimar el valor de las variables independientes (x 1 , x 2 … xp), al aplicarle la función D podremos atribuirle un grupo de pertenencia (esto es, efectuar una retrodicción sobre su
CUADRO 3.XXX: Lógica del análisis discriminante II
<<10 Práctica III, Paleontología y Evolución, : Profesores responsables: Palmqvist, Pérez-Claros y Figueirido
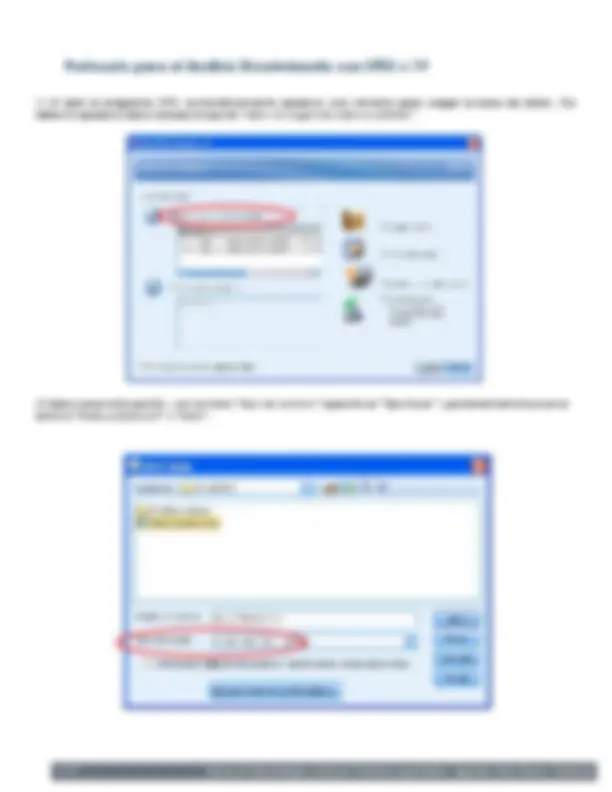
Al abrir el programa SPSS, automáticamente aparece una ventana para cargar la base de datos. Por defecto aparece seleccionada la opción “abrir un origen de datos existente”.
Seleccionar esta opción y en el menú “tipo de archivo” especificar “Tipo Excel” y posteriormente buscar el archivo “Datos práctica II” y “Abrir”.
Protocolo para el Análisis Discriminante con SPSS v.
<<12 Práctica III, Paleontología y Evolución. Profesores responsables: Figueirido, Pérez-Claros y Palmqvist
Seguidamente aparece una ventana de especificaciones del archivo Excel. Aquí, en nuestro caso, simplemente hay que “Aceptar” y automáticamente se abrirá el visor datos del SPSS donde podemos observar nuestro archivo Excel correctamente cargado.
Explora en la esquina izquierda el visor variables del SPSS donde aparecen una serie de especificaciones de las variables.
<<13 Práctica III, Paleontología y Evolución. Profesores responsables: Figueirido, Pérez-Claros y Palmqvist
En los siguientes menús, tenemos que seleccionar las siguientes opciones que serán explicados debidamente a lo largo del desarrollo de la práctica. ).
���� En “Clasificar” selecciona, la opción de “clasificación dejando una fuera”. Esto es para aplicar el
método de “leave one out” (véase texto para un mayor detalle).
1
���� En “Guardar” hay que seleccionar las tres casillas: (1) Grupo de pertenencia pronosticado; (2)
Puntuaciones discriminantes; (3) Probabilidades de pertenencia.
<<15 Práctica III, Paleontología y Evolución. Profesores responsables: Figueirido, Pérez-Claros y Palmqvist
Cuestionario
1) ¿Cuáles son las diferencias entre el ACP y el ACD?
2) ¿Cuántas funciones discriminantes se derivan en tu análisis?
3) ¿Puedes separar los grupos tróficos previamente preestablecidos en la muestra de
Ursidae?
4) ¿El ACD permite una buena discriminación entre los grupos?
5) ¿Qué características morfológicas poseen los miembros de cada grupo?
6) ¿Qué dieta poseería A. simus según el ACD?
7) ¿Qué limitaciones posee este tipo de análisis?
Bibliografía de referencia
Carrasco, J.L. & Hernán Huerta, M.A. 1993. Estadística multivariante para las Ciencias de la Vida.
Editorial Ciencia: Madrid.
Davis, J. C. 1973. Statistics and data analysis in geology. John Wiley Inc.
Mardia, K. V., Kent, J. T. and Bibby, J. M. 1979. Multivariate Analysis. Probability and Mathematical
Statistics. Academic Press: London.
Quinn, G. & Keough, M. 2002. Experimental Design and Data Analysis for Biologists. Cambridge
University Press.
Peña, D. 2002. Análisis de datos multivariantes. McGrawHill Ineteramericana: Madrid.
Zelditch, M. L., Swinderski, D. L., Sheets, H. D. and Fink, W. L. 2004. Geometric morphometrics for
biologist: a primer. Elsevier Academic Press: New York/London
<<18 Práctica III, Paleontología y Evolución. Profesores responsables: Figueirido, Pérez-Claros y Palmqvist