¡Descarga TEMA 6: SEÑALIZACIÓN CELULAR y más Apuntes en PDF de Biología Aplicada solo en Docsity!
TEMA 5 SEÑALIZACIÓN
CELULAR
La señalización celular es la capacidad para recibir señales desde el medio externo y responder. Procariotas y eucariotas. La naturaleza química de las moléculas señal es de naturaleza variable. Son moléculas que van desde gases como el NO, aminoácidos, nucleótidos, lípidos, péptidos o proteínas. La respuesta celular es muy variable, sobrevivir, dividirse, suicidarse, cambiar la formología de la célula o el metabolismo. Siempre se hace mediante moléculas señalizadoras (moléculas señal que van a actuar sobre una célula diana (target cell) que tiene un receptor específico para esa molécula señal. Esto es un área prioritaria de investigación, ya que muchos cánceres surgen por una alteración de las vías de señalización. Existen dos grandes grupos:
- Señalización directa por contacto : donde la molécula señal está en la membrana de la célula señalizadora y el receptor está en la membrana de la célula diana. Ej. Caderinas/cateninas. Vía de señalización de Wnt, integrinas, sindecanos.
Señalización mediante moléculas secretadas : donde la célula señalizadora sintetiza una molécula señal que recorre una cierta distancia hasta unirse con el receptor de la célula diana, no hay un contacto directo. En función de la distancia recorrida por la molécula señal: autocrina, endocrina y paracrina.
a. Autocrina : la molécula señalizadora tiene el propio receptor. Ej. Células tumorales, fabrican factores para inducir la proliferación.
b. Paracrina : la molécula señal
recorre una distancia algo mayor pero las células diana están relativamente cerca. Estas moléculas señal reciben el nombre de mediadores locales. Ej. Sinápsis química, neurotransmisores. c. Endocrina : las células señalizadora (células endocrinas) fabrican las
moléculas señal, hormonas,
que pasan al sistema circulatorio y pueden recorrer grandes distancias hasta que salen del torrente sanguíneo para llegar a la célula diana. Receptores diferentes pueden activar varias vías de señalización. Un receptor puede desencadenar una misma vía o diferentes vías de señalización. Estas vías están modificadas en las células tumorales. Otra idea es que lo receptores activados diferentes pueden activar distintas vías de señalización pero que se alguna forman influyen una en la otra. Una determinada molécula señal puede inducir respuestas diferentes en distintos tipos de células diana, ej. Acetilcolina, que cuando su receptor se localiza en el músculo cardiaco se produce una disminución en la frecuencia de contracción del músculo cardiaco, en cambio en una célula del músculo esquelético induce la contracción y en la glándula salivas provoca la secreción.
1.- Bases de la señalización celular
Etapas.
- Reconocimiento : recepción de la señal.
- Transducción : a través de vías de señalización que actúan en cascada diferentes proteínas y moléculas.
- Respuesta : donde haya activación y transcripción de determinados genes.
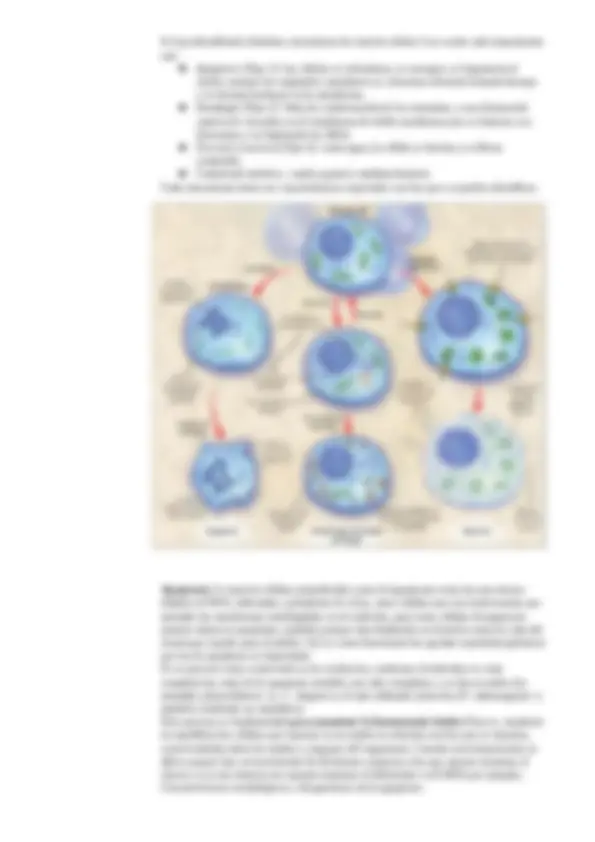
Tipos de receptores:
- Receptores intracelulares : donde la
molécula señal es pequeña e
hidrofóbica para que pueda atravesar
la membrana plasmática por difusión
simple, y el receptor está en el interior
- Ensamblaje de un complejo de señalización sobre un receptor activado : varias moléculas son capaces de unirse fosforilando a una misma proteína por diferentes sitios, es una manera rápida de transmitir la señal. Ej. RTKs
- Complejos de señalización por unión a fosfolípidos hiperfosforilados : están en la monocapa interna de la bicapa lipídica. Se tiene un receptor inactivo y en la monocapa interna hay fosfolípidos de inositol que llevan dos fosfatos. Las proteínas de señalización están sueltas inactivas. Al llegar la molécula señal y activa el receptor, este añade un fosfato más al inositol. Esto hace que se unan las proteínas señalizadoras inactivadas.
La transducción de señales se basa en interacciones entre los denominados dominios de unión y los llamados motivos de reconocimiento. El dominio de unión es una zona de la molécula determinada que es capaz de reconocer y de unir un motivo determinado que haya en otra proteína. Los dominios y los motivos se dan entre proteínas excepto en la interacción de la proteína con el lípido.
INTERRUPTORES MOLECULARES EN LAS VÍAS DE SEÑALIZACIÓN
Las proteínas de señalización intracelular que actúan como interruptores moleculares ON/OFF. Dos grandes grupos:
- Señalización con fosforilación : quinasas que añades grupos fosfato para encender. Las fosfatasas apagan desfosforilando. Las quinasas siempre añaden grupos fosfato a partir de la hidrólisis del ATP por acción de una fosfatasa. En la mayoría de los casos, una proteína es inactiva que se activa por una quinasa y se inactiva por una fosfatasa. En algunos casos una proteína está activa cuando está desfosforilada e inactiva al estar fosforilada. La quinasa siempre actúa con una fosfatasa.
- Proteínas de unión a GTP. Cuando la proteína está unida a GDP está inactiva, al pasar a GTP se activa. Existen dos tipos de proteínas:
TIPOS DE RECEPTORES DE SUPERFICIE
Hay tres grandes grupos de receptores de superficie. a. Receptores acoplados a canales iónicos : El receptor es un canal regulado por ligando. Cuando la molécula señal se une al receptor se va a abrir el canal permitiendo el paso de un determinado ion o iones dando una respuesta celular. Ej. Es la unión neuromuscular, donde la molécula señal es la ACh que se une al receptor permitiendo que se abra y entren iones sodio.
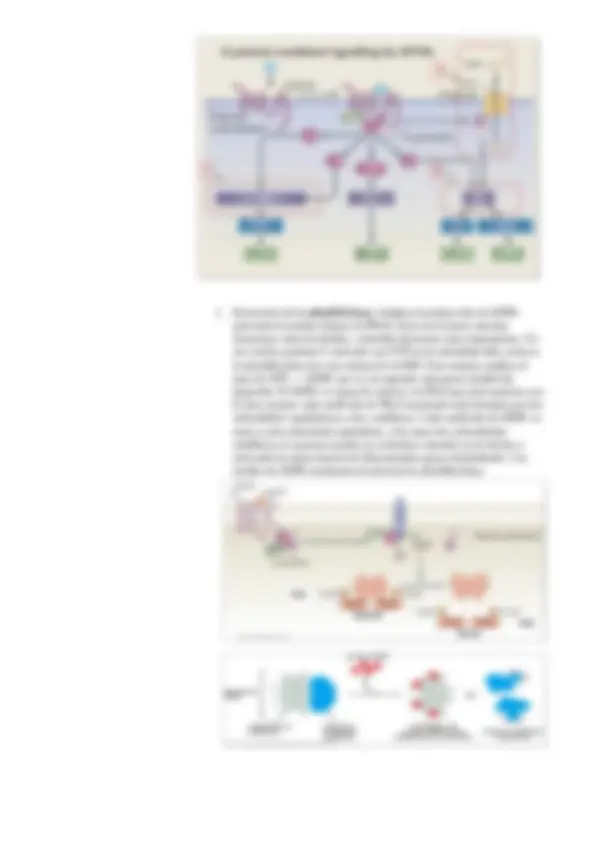
2.- Señalización mediante receptores de superficie celular acoplados a proteínas G y a enzimas
b. (^) Receptores acoplados a proteínas G ( G protein – coupled receptors, GPCR): Es uno de los grupos más diversos y mayores de proteínas codificadas por el genoma humano. Se calcula que tenemos unos 800 genes que codifican para estos receptores, y la molécula señal es variada: hormonas, neurotransmisores, estímulos sensoriales 8luz, sonido…). Estos receptores controlas funciones fisiológicas muy diversas: neurotransmisión, liberación de hormonas, respuesta inmune, participan en la contracción muscular, regulación de la presión sanguínea. Son tantas la funciones que controlan que se calcula que más del 60% de los fármacos van dirigidos a receptores acoplados a proteínas G que no funcionan adecuadamente.
Estos receptores están en la membrana, atravesándo la membrana 7 veces en alfa – hélice, tienen el extremo C – terminal en el citosol y el N – terminal en la medio extracelular. La unión de ligandos inducen cambios
- Activación de la adenililciclasa : Implica la producción de AMPc activando la protein kinasa A (PKA). Esta vía la hacen muchas hormonas como la tiroides, controlan funciones muy importantes. Un vez está la proteína G activada con GTP en la subunidad alfa, activa a la adenililciclasa (es una enzima de la MP). Esta enzima cataliza el paso de ATP --> AMPc que es un segundo mensajero (molécula pequeña). El AMPc es capaz de activar a la PKA que está inactiva, eso lo hace porque cada molécula de PKA inactivada está formada por dos subunidades reguladoras y dos catalíticas. Cada molécula de AMPc se unen a cada subunidad reguladora, y las otras dos subunidades catalíticas se separan estando ya activadas entrando en el núcleo y activando la transcripción de determinados genes fosforilando. Los niveles de AMPc aumentan al activarse la adenililciclasa.
- Activación de la fosfolipasa Cβ (PLC β): Implica la formación de diacilglicerol (DAG), inositoltrifosfato (IP3) e iones Ca+2, los tres son segundos mensajeros. El fosfatidilinositol bifosfato en la monocapa interna, es hidrolizado por la fosfolipasa Cβ en DAG que se queda en la monocapa interna y en IP3. Esta es una molécula soluble que se mueve por el citosol hasta llegar a la membrana del retículo endoplasmático donde se une a un canal regulado por ligando que hay en la membrana del RE. En este caso el ligando es IP3. Si no está este no hay salida de iones Ca+2. Estos iones son segundos mensajeros muy importantes en las células, SIEMPRE que aumentan su concentración en el citosol hay una respuesta celular. Una de las respuestas es activar junto con DAG a la PKC. Esta PKC fosforilará activando a otras proteínas.
Apertura de canales, activación de canales iónicos.
La ACh activa a una proteína G, donde la subunidad βγ que va a abrir un canal iónico que permite la salida de ione K+, hiperpolariza la membrana dando lugar a una reducción de la frecuencia del ritmo cardiaco. Inactivación de los Receptores Acoplados a Proteínas G: La señalización por acoplamiento a proteínas G puede desactivarse cuando el receptor es fosforilado por un grupo de kinasas específico para fosforilar por el dominio del citosol al receptor. Este grupo de kinasas se llama GRK, que son kinasas de los receptores acoplados a proteínas G. Actúan estas GRK, y una vez el receptor está fosforilado, la proteína G está inactiva y a la cual se le une también la arrestina (sólo cuando está fosforilado) produciéndose una endocitosis mediada por clatrina que va a internalizar a ese receptor para degradarse en el lisosoma o reciclado para la membrana. El receptor y la proteína G dejan de funcionar al unirse estas kinasas y fosforila al receptor, la arrestina facilita la formación de las vesículas de clatrina. Este grupo de kinasas es un grupo de muchísima investigación.
Las vías de señalización de los receptores acoplados a proteína G están implicados en el mantenimiento del cáncer, en la tumorgénesis. Dado que tiene distintas rutas son muy importantes y complicadas.
c. Receptores acoplados a enzimas ( _importante):_* Estos receptores acoplados a enzimas, son receptores transmembrana que por su dominio citosólico poseen ellos mismos actividad enzimática o se asocian directamente a una enzima. Pueden activar algunas rutas que también activan los receptores acoplados a proteínas G, algunas son las mismas rutas pero no se sabe muy bien porqué.
Hay distintas clases, los más abundantes y frecuentes son los receptores Tirosina Kinasa (RTKs). La molécula señal para la mayoría son factores de crecimiento que se requieren a concentraciones muy bajas (10-9 a 19-11 M). La respuesta de la célula suele ser lenta, puede durar horas, variada como crecimiento, proliferación, diferenciación, cambios en el citoesqueleto, movimiento de la célula, supervivencia. Las alteraciones de estas están implicadas directamente en el cáncer.
Hay varias moléculas señal donde los más importantes son:
■ Factor de crecimiento epidérmico (EGF): estimula el crecimiento, supervivencia y proliferación de distintos tipos celulares. Hay cuatro receptores tirosina kinasa que reciben los nombres de EGFR (receptor de factor de crecimiento epidérmico) que también se conoce como ER1, luego tenemos a ER2, ER3 y ER4.
■ Factor de crecimiento del endotelio vascular (VEGF): es importante porque estimula la angiogénesis, es decir, la formación de nuevos vasos sanguíneos.
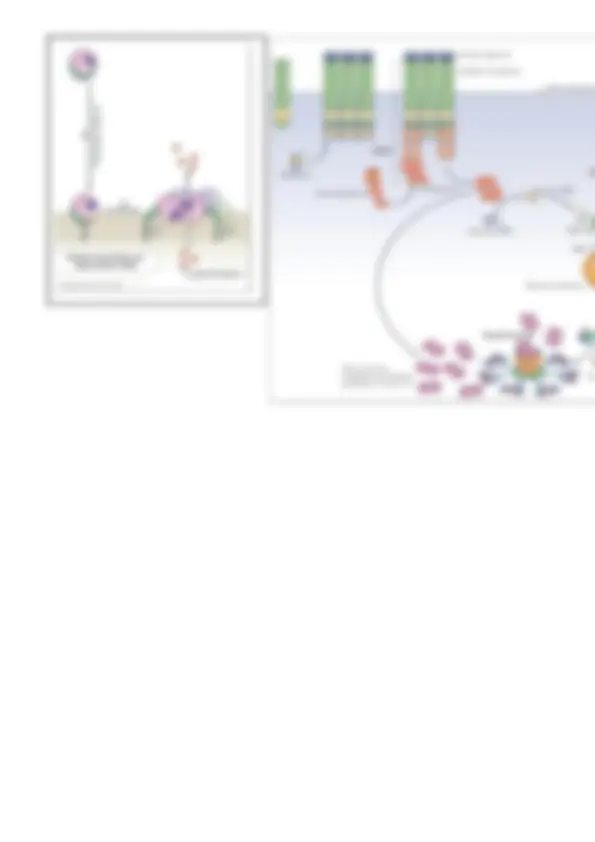
Existen 58 receptores tirosina kinasas en el genoma humano, la mayoría son proteínas transmembrana de paso único excepto el receptor de la insulina que es un dímero. Pero todos los demás, el receptor del factor de crecimiento epidérmico o derivado de las plaquetas por ejemplo, atraviesan solo una vez la bicapa lipídica. Activación de los RTKs ( importante) El receptor tirosina kinasa está en un modo inactivo pero cuando lleva la molécula señal, en general un factor de crecimiento, este factor suele dimerizar, se presenta como dos moléculas, y el receptor también lo hace.
- Activación de la PI3K se forma PIP3F 0 E 0 se activa Akt
- Activación de la fosfolipasa Cγ 1. ACTIVACIÓN DE RAS Ras es un conjunto de una súperfamilia de GTPasas monoméricas con distintos componentes y distintas funciones en la célula, una de esas familias participa en esta señalización de los RTKs en concreto participan: H-Ras, K-Ras y N- Ras. Ras es una GTPasa monomérica anclada a la monocapa interna y que en su forma inactiva lleva GDP, cuando salga éste y entre GTP Ras se activa, para esto hace falta que intervenga un factor intercambiador de nucleótidos de guanina, un GEF. Interesa que Ras deje de estar activo en un momento dado y para esto interviene un GAP para acelerar la hidrólisis.
Al RTK activado se le une la proteína adaptadora GRB2 que reconoce el motivo de reconocimiento, motivo fosfotirosina del receptor, se une gracias a que esta proteína adaptadora tiene un dominio SH2 de reconocimiento. Esta proteína es capaz también de unir a la proteína Sos (Ras-GEF), esta proteína Sos se une a los dominios SH3 de Grb2 los cuales reconocen los motivos ricos en prolinas de Sos. ( Importante).
Antes de producirse esto la Sos está libre en el citoplasma, la proteína adaptadora Grb2 ha conseguido que la Sos esté cerca de la membrana plasmática. Esto es importante porque Sos es la GEF de Ras, es el factor intercambiador de nucleótidos de guanina de Ras, Ras está en la membrana y
por eso necesita que GEF esté cerca para intercambiar GDP por GTP. Se ha conseguido que esté GEF cerca y se consigue el intercambio, Sos favores la salida de GDP en Ras y la entrada de GTP. Teniendo ya a Ras activada y provocando la cascada de señales.
Una de las rutas que puede seguir Ras está activado es desencadenar la ruta de las MAP quinasa. Es una cascada de señalización muy conservada en la evolución que incluye la fosforilación secuencial de 3 quinasas:
■ MAP quinasa quinasa quinasa
(MAPKKK): Ref
■ MAP quinasa quinasa (MAPKK):
MEK
■ MAP quinasa: ERK (extracelular
signal-regulated kinase)
ERK fosforilada se trasloca al núcleo donde fosforila y activa factores de transcripción que controlan la expresión de genes que se requieren para el crecimiento celular, la diferenciación y la supervivencia. Por lo tanto, una activación constante de esta ruta hará que haya un incremento de la proliferación, de la supervivencia y que se desarrolle un cáncer.
Un RTK activado también puede activar al enzima fosfatidil inositol 3 quinasa (PI3K) , añade grupos fosfatos aunque ahora es a un fosfolípido de la monocapa interna no a una proteína, al fofatidil inositol – bifosfato (PIP2), formando fofatidil inositol – trifosfato (PIP3). Ese PIP3 que se acaba de formar permite que se le una dos proteínas que son PDK1 y Akt , estas se unen porque ambas tienen un dominio de homología con la plecstrina (dominio PH), éste reconoce y une un motivo que es el PIP3. PDK1 junto con otro complejo proteico llamado mTOR en su forma complejo 2 mTORC2 se encargan de fosforilar a Akt. De esta forma, sólo cuando se ha fosforilado Akt pasa a un estado activo. Se separa de la membrana plasmática y activa múltiples vías de señalización. (La Akt también se conoce como PKB , aunque actualmente se utiliza más Akt).
Esta ruta de la PI3 quinasa, es inactivado por la proteína PTEN , que es una fosfatasa. PKB/Akt media numerosas funciones celulares como: ■ Angiogénesis
■ Metabolismo
■ Crecimiento
■ Proliferación
■ Supervivencia
■ Síntesis de proteínas
■ Transcripción
■ Apoptosis (inhibición de la apoptosis) Todas estas características recuerdan a una célula tumoral.
3. ACTIVACIÓN DE LA FOSFOLIPASA Cγ (PLCγ) La fosfolipasa Cγ se encarga de fosforilar el PIP2 en DAG que se queda en la membrana plasmática y en IP3. El IP3 difunde y llega a la membrana del retículo donde hay un canal de membrana regulado por ligando donde salen iones Ca+2. Tanto DAG como IP3 son segundos mensajeros que activan la PKC. Además los iones Ca+2 activan a la calmodulina. Ras también puede activar a la PI3K y a la fosfolipasa Cε (épsidon). Un receptor acoplado a una proteína G también puede activar la vía de señalización de Ras. Todo está entremezclado.