













Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: anatomia, Profesor: Cristina Monforte (Estructura y funció del cos humà., Carrera: Infermeria (Gimbernat), Universidad: UAB
Tipo: Apuntes
1 / 19

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Capas de la pared pericárdica
La pared cardiaca se divide en tres capas:
Epicardio : compuesto por dos planos tisulares. El más eterno es una lamina delgada y transparente que también se conoce como capa visceral del pericardio seroso y está formada por mesotelio. Debajo del mesotelio existe una capa variable de tejido fibroelástico y tejido adiposo. El epicardio contiene vasos sanguíneos, linfáticos y vasos que irrigan el miocardio.
Representa el 95% de la pared cardíaca. Las fibras del musculo cardíaco están organizadas en haces que se dirigen en sentido diagonal alrededor del corazón y generan la poderosa acción de bombeo. El musculo cardíaco es involuntario como el musculo liso.
sobre una capa delgada de tejido conectivo. Tapiza las cámaras cardiacas y recubre las válvulas cardiacas. Minimiza la superficie de fricción cuando la sangre pasa por el corazón y se continúa con el endotelio de los grandes vasos que llegan y salen del corazón.
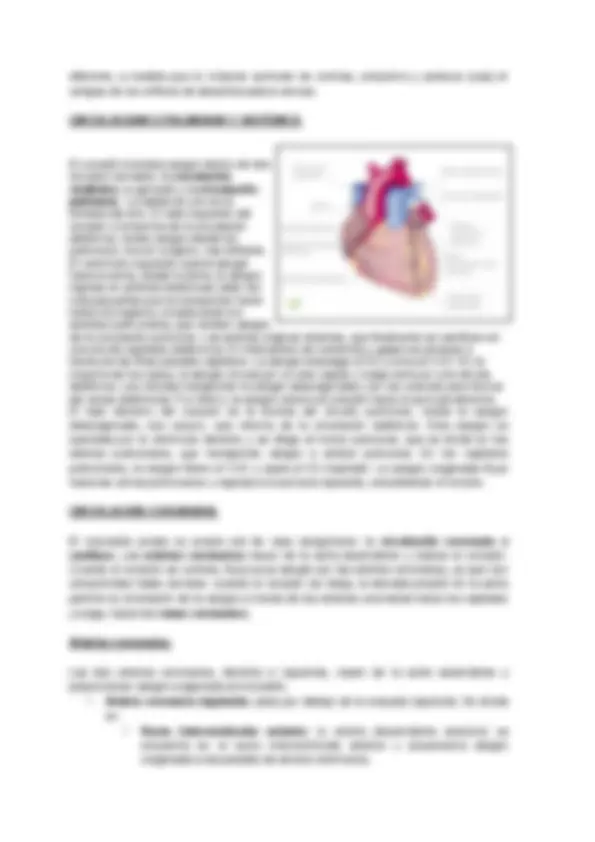
El corazón posee cuatro cámaras. Las dos cámaras superiores son las aurículas ( atrios ), y las dos inferiores los ventrículos. Las dos aurículas reciben sangre de los vasos que la traen de regreso al corazón, las venas, mientras que los ventrículos eyectan desde el corazón hacia los vasos que la distribuyen, las arterias. En la cara anterior de cada aurícula se encuentra una pequeña bolsa denominada orejuela. Cada orejuela aumenta ligeramente la capacidad de las aurículas, lo que permite a éstas recibir un volumen de sangre mayor. Además, en la superficie del corazón existe una serie de surcos que contiene vasos coronarios y una cantidad de variable de grasa. Cada surco marca el limite externo entre dos cámaras cardiacas. El surco coronario profundo rodea a casi todo el corazón y limita dos sectores: el sector auricular (superior) y el ventricular (inferior). El surco interventricular anterior es una hendidura poco profunda, ubicada en la cara anterior del corazón, que marca el limite entre el ventrículo derecho y el izquierdo. Se continúa en la cara posterior como surco interventricular posterior , delimitando ambos ventrículos en la parte posterior del corazón.
cava inferior y el seno coronario. Sus paredes tienen un espesor promedio de 2 a 3 mm. Las paredes anterior y posterior difieren mucho entre sí. La pared posterior es lisa; la pared anterior es trabeculada, debido a la presencia de crestas musculares denominadas músculos pectíneos , que también se extienden dentro de la orejuela. Entre la aurícula derecha y la izquierda se encuentra un tabique delgado denominado septum o tabique interauricular. Una formación anatómica oval remanece del foramen ovale, una comunicación interauricular en el corazón fetal que normalmente se cierra luego del nacimiento. La sangre pasa desde la aurícula derecha hacia el ventrículo derecho a través de la válvula tricúspide ,
Además de músculo, la pared cardíaca también contiene tejido conectivo denso que forma el esqueleto fibroso del corazón. Esta estructura consiste en cuatro anillos de tejido conectivo denso que rodean las válvulas cardíacas fusionándolas entre sí y uniéndolas al tabique interventricular. Al mismo tiempo, el esqueleto fibroso también evita el sobreestiramiento de las válvulas al pasar la sangre a través de ellas. Sirve como punto de inserción a los haces de fibras musculares cardíacas y como aislante eléctrico entre las aurículas y los ventrículos.
Cuando una cámara cardíaca se contrae eyecta un determinado volumen de sangre dentro del ventrículo o hacia una arteria. Las válvulas se abren y cierran en repuesta a los cambios de presión, a medida que el corazón se contrae y relaja. Cada una de las válvulas contribuye a establecer el flujo en un solo sentido, abriéndose para permitir el paso de la sangre y luego cerrándose para prevenir el reflujo.
Las válvulas mitral y tricúspide también reciben el nombre de válvulas aurculoventriculares o atrioventriculares (AV). Cuando una válvula AV está abierta, los extremos de las valvas se proyectan dentro del ventrículo. Cuando los ventrículos se encuentran relajados, los músculos papilares también lo están, las cuerdas tendinosas están flojas y la sangre se mueve desde un sitio de mayor presión, la aurícula, hacia otro de menor presión, el ventrículo, gracias a que las válvulas AV están abiertas. Cuando los ventrículos se contraen, la presión de la sangre empuja las valvas hacia arriba hasta que sus borde se juntan y cierran el orificio auriculoventricular. Al mismo tiempo, los músculos papilares se contraen estirando las cuerdas tendinosas. Esto evita que las cúspides valvulares reviertan y se abran a la cavidad auricular por acción de la elevada presión ventricular. Si las cuerdas tendinosas o las válvulas AV se dañan, la sangre puede refluir hacia las aurículas durante la contracción ventricular.
Las válvulas aórticas y pulmonares también se conocen como válvulas semilunares (SL) porque estén formadas por tres valvas con aspecto de medialuna. Cada valva se une a la pared arterial en su borde convexo externo. Las válvulas SL permiten la eyección de la sangre desde el corazón hasta las arterias, pero evitan el reflujo de la sangre hacia los ventrículos. Los borde libres de las valvas se proyectan hacia la luz de la arteria. Cuando en ventrículo se contrae, la presión aumenta dentro de las cámaras. Las válvulas SL se abren cuando la presión ventricular excede la tensión arterial permitiendo así la eyección de la sangre desde los ventrículos hacia el tronco pulmonar y la aorta. A medida que los ventrículos se relajan, la sangre comienza a empujar las cúspides valvulares, haciendo que las válvulas semilunares de cierren. No hay válvulas que reguarden los orificios de desembocadura de las venas cava superior e inferior de la aurícula derecha o los de las venas pulmonares, en la aurícula izquierda. Cuando las aurículas se contraen, una pequeña cantidad de sangre refluye desde las aurícula hacia dichos vasos. El reflujo se minimiza por medio de un mecanismo
diferente; a medida que el músculo auricular se contrae, comprime y produce (casi) el colapso de los orificios de desembocadura venoso.
El corazón bombea sangre dentro de dos circuitos cerrados: la circulación sistémica (o general) y la circulación pulmonar. La salida de uno es la entrada del otro. El lado izquierdo del corazón e la bomba de la circulación sistémica; recibe sangre desde los pulmones, rica en oxígeno, roja brillante. El ventrículo izquierdo eyecta sangre hacia la aorta, desde la aorta, la sangre ingresa en arterias sistémicas cada vez más pequeñas que la transportan hacia todos los órganos, exceptuando los alvéolos pulmonares, que reciben sangre de la circulación pulmonar. Las arterias originan arteriola, que finalmente se ramifican en una red de capilares sistémicos. El intercambio de nutrientes y gases se produce a través de las finas paredes capilares. La sangre descarga el O2 y toma el CO2. En la mayoría de los casos, la sangre circula por un solo capilar y luego entra en una vénula sistémica. Las vénulas transportan la sangre desoxigenada y se van uniendo para formar las venas sistémicas. Por último, la sangre retorna al corazón hacia la aurícula derecha. El lado derecho del corazón es la bomba del circuito pulmonar; recibe la sangre desoxigenada, rojo oscuro, que retorna de la circulación sistémica. Esta sangre es eyectada por el ventrículo derecho y se dirige al tronco pulmonar, que se divide en las arterias pulmonares, que transportan sangre a ambos pulmones. En los capilares pulmonares, la sangre libera el CO2 y capta el O2 inspirado. La sangre oxigenada fluye hacia las venas pulmonares y regresa a la aurícula izquierda, completando el circuito.
El miocardio posee su propia red de vaso sanguíneos: la circulación coronaria o cardíaca. Las arterias coronarias nacen de la aorta ascendente y rodean el corazón. Cuando el corazón se contrae, fluye poca sangre por las arterias coronarias, ya que son comprimidas hasta cerrarse. cuando el corazón se relaja, la elevada presión en la aorta permite la circulación de la sangre a través de las arterias coronarias hacia los capilares y luego, hacia las venas coronarias.
Arterias coronarias
Las dos arterias coronarias, derecha e izquierda, nacen de la aorta ascendente y proporcionan sangre oxigenada al miocardio.
en:
encuentra en el surco interventricular anterior y proporciona sangre oxigenada a las paredes de ambos ventrículos.
directamente en la aurícula derecha.
Cuando la obstrucción de una arteria coronaria priva al músculo cardíaco del aporte de oxígeno, la reperfusión posterior (restablecimiento del flujo sanguíneo) puede generar aun mayor daño tisular. Este efecto se debe a la formación de radicales libres de oxígeno generados a partir del oxígeno reintroducido. Los radicales libres son moléculas eléctricamente cargadas que poseen un electrón desapareado. Estas moléculas reactivas y muy inestables, provocan reacciones en cadena que conducen al daño y a la muerte celular. Para contrarrestar los efectos de los radicales libres, las células producen enzimas que los convierten en sustancias menos reactivas. Además, ciertos nutrientes como las vitaminas C y E, los betacarotenos, el zinc y el selenio poseen funciones antioxidantes que les permiten remover los radicales libres generados por el oxígeno.
En comparación con las fibras musculares esqueléticas, las fibras musculares cardíacas son más cortas y menos circulares en sección transversa. Presenta ramificaciones, que les confieren la apariencia “en peldaños de escalera” característica de las fibras musculares cardíacas. Una fibra mide de 50 a 100 micrómetros de longitud y 14 de diámetro. Presenta un solo núcleo de localización central, aunque algunas células pueden presentar dos núcleos. Los extremos de las fibras se conectan con las fibras vecinas a través de engrosamientos transversales del sarcolema, denominado discos intercalares. Estos discos contienen desmosomas , que unen las fibras entre sí, y uniones en hendidura (gap) que permiten la conducción de los potenciales de acción desde una fibra muscular hasta las fibras vecinas. Las uniones gap permiten que todo el miocardio de las aurículas y de los ventrículos se contraiga como una única unidad coordinada. Las mitocondrias son más grandes y numerosas en las fibras musculares cardíacas que en las esqueléticas. En una fibra cardíaca ocupan el 25% del citosol, mientras que en una esquelética solo el 2%. Las fibras musculares cardíacas poseen la misma disposición de filamentos de actina y miosina. Las mismas bandas, zonas y discos Z que las fibras esqueléticas. Los túbulos transversos del miocardio son más anchos, pero más escasos que los del músculo esquelético; el único túbulo transverso por sarcómero se localiza en el disco Z. El retículo sarcoplasmático es algo más pequeño que el de las fibras esqueléticas. En consecuencia, el músculo cardíaco presenta menores reservas intercelulares de Ca2+.
La actividad cardíaca eléctrica intrínseca y rítmica permite que el corazón pueda latir toda la vida. La fuente de esta actividad es una red de fibras musculares cardíacas especializadas denominadas fibras cardíacas automáticas , debido a que son autoexcitables. Las fibras automáticas generan potenciales de acción en forma repetitiva que disparan las contracciones cardíacas. Continúan estimulando el corazón para que
lata, aún después de haber sido extirpado del cuerpo y de que todos sus nervios hayan sido cortados. Durante el desarrollo embrionario, solo el 1% de las fibras musculares cardíacas se diferencian en fibras automáticas; estas fibras cumplen dos funciones importantes:
causa la contracción cardíaca.
excitación cardíaca progrese a través del corazón. El sistema de conducción asegura que las cámaras cardíacas sean estimuladas para contraerse en forma coordinada, lo que hace del corazón una bomba efectiva. Los problemas con las fibras automáticas pueden causar arrítmias, donde el corazón late en forma anómala, demasiado rápido o demasiado despacio
Los potenciales de acción cardíacos se propagan con la siguiente secuencia:
localizado en la aurícula derecha, justo por debajo del orificio de desembocadura de la vena cava superior. Las células del nodo SA no tienen un potencial de reposo estable. En lugar del ello, se despolarizan en forma contínua y alcanzan espontáneamente el potencial umbral. Las despolarización espontánea es un potencial marcapasos. Cuando el potencial marcapasos alcanza el umbral se desencadena un potencial de acción. Cada potencial de acción del nodo SA se propaga a través de ambas aurículas, por medio de la uniones en hendidura presente en los discos intercalares de las fibras musculares auriculares. Siguiendo el potencial de acción, las aurículas se contraen.
potencial de acción llega al nodo auriculoventricular o atrioventricular (AV) , localizado en el tabique interauricular, delante del orificio de desembocadura del seno coronario.
auriculoventricular o atrioventricular (también conocido como haz de His ). Este es el único sitio por donde los potenciales de acción puede propagarse desde las aurículas hacia los ventrículos. (En el resto del corazón, el esqueleto fibroso aísla eléctricamente las aurículas de los vetrículos.)
izquierda que se extienden a través del tabique interventricular hacia el vértice cardíaco.
rápidamente el potencial de acción desde el vértice cardiaco hacia el reto del miocardio ventricular. Luego, los ventrículos se contraen y empujan la sangre hacia las válvulas semilunares.
Las fibras automáticas del nodo SA iniciarán por su cuenta un potencial de acción cada 0,6 segundos, o 100 veces por minuto. Así, el nódulo sinoauricular es el marcapasos natural. Esta frecuencia es mayor que la del resto de las fibras automáticas. Los impulsos nerviosos del sistema nervioso autónomo y de hormonas (como la adrenalina) modifican la frecuencia y la fuerza de cada latido cardíaco, pero no establecen el ritmo fundamental. Por ejemplo, en una persona en reposo, la acetilcolina
contracción cardíaca. La adrenalina, aumenta la fuerza de contracción mediante el aumento del flujo de entrada de Ca2+ al citosol. El período refractario (intervalo de tiempo durante el cual no puede desencadenarse una segunda contracción) de una fibra muscular cardíaca dura más que la de contracción. En consecuencia, no puede iniciarse una nueva contracción hasta que la fibra no se haya relajado correctamente. Por esta razón, la tetania (contracción sostenida) no se produce en el músculo cardíaco como lo hace en el esquelético. La función de bomba de los ventrículo depende de la alternancia de contracción (cuando eyectan sangre) y relajación (cuando se llenan). Si el corazón pudiera generar una contracción tetánica, el flujo sanguíneo cesaría.
A diferencia del músculo esquelético, el músculo cardíaco produce poco del ATP que necesita por medio de respiración celular anaeróbica. Por el contrario, depende casi exclusivamente de la respiración celular aeróbica que se lleva a cabo en las numerosas mitocondrias. El oxígeno necesario difunde desde la sangre de la circulación coronaria y es liberado en el interior de las fibras musculares cardíacas desde la mioglobina allí presenta. Las fibras musculares cardíacas utilizan varias fuentes energéticas para producir ATP mitocondrial. En una persona en reposo, el ATP cardíaco proviene fundamentalmente de la oxidación de ácidos grasos (60%) y de glucosa (35%), con pequeñas contribuciones de la oxidación del ácido láctico, aminoácidos y cuerpos cetónicos. Durante el ejercicio, aumenta el uso de ácido láctico producido por la contracción activa de los músculos esqueléticos. Como en el esquelético, en el músculo cardíaco parte del ATP producido proviene de la fosfocreatina. Un signo que confirma la ocurrencia de un infarto de miocardio es la presencia en sangre de creatincinasa (CK), la enzima que cataliza la transferencia de un grupo fosfato desde la fosfocreatina al ADP para producir ATP. En condiciones normales, la CK y otras enzimas están confinadas dentro de las células. Las fibras musculares esqueléticas o cardíacas lesionadas y las que se están muriendo liberan CK hacia la circulación.
A medida que los potenciales de acción se propagan, generan corrientes eléctricas que pueden ser detectadas desde la superficie corporal. Un electrocardiograma es un registro de las señales eléctricas. El ECG es una representación de los potenciales de acción producidos por todas las fibras musculares cardíacas durante cada latido. En la práctica clínica, para realizar el ECG se colocan electrodos en los brazos y piernas (derivaciones de los miembros) y en seis
ubicaciones a nivel torácico (derivaciones precordiales). Cada electrodo detecta una
actividad eléctrica levemente diferente, según la posición que ocupa respecto al corazón.
Mediante la comparación de los trazaos entre sí, y con trazados normales, es posible determinar si:
En un trazado típico, aparecen en cada latido tres ondas claramente reconocibles.
la despolarización auricular , que se propaga desde el nodo SA, a través de las fibras contráctiles en ambas aurículas.
continúa con una importante onda triangular positiva y termina con una onda negativa. Este complejo representa la despolarización ventricular rápida , a medida que el potencial de acción progresa a través de las fibras ventriculares contráctiles.
repolarización ventricular y aparece justo cuando los ventrículos están comenzando a relajarse. La onda T es más pequeña y más ancha que el complejo QRS debido a que la repolarización se produce más lentamente que la despolarización. Durante la fase de la meseta de la despolarización sostenida, el trazado del ECG permanece plano.
Las ondas P grandes indican:
Una onda Q de mayor magnitud puede indicar:
Las ondas R grandes suelen indicar:
La onda T es más aplanada de lo normal cuando:
enfermedad coronaria. Una onda T puede estar más elevada en:
El análisis del ECG también incluye la medición de los espacios existentes entre las ondas, denominados intervalos o segmentos.
complejo QRS. Representa el tiempo de conducción desde el comienzo de la excitación auricular hasta el inicio de la despolarización ventricular. Dicho de otro modo, el intervalo P-Q es el tiempo requerido para que un potencial de acción viaje a través de la aurícula, el nodo AV y las fibras remanentes del sistema de conducción.
(diástole ventricular). A los 0,6 s se completa la repolarización ventricular y las fibras ventriculares contráctiles se encuentran relajadas.
Durante los siguientes 0,2 s fibras contráctiles de las aurículas y ventrículos están relajadas. A los 0,8 s, la onda P aparece nuevamente en el ECG, la aurícula comienza a contraerse y el ciclo se repite.
EL CICLO CARDÍACO
Un ciclo cardíaco incluye todos los fenómenos asociados con un latido cardíaco. Por lo tanto, un ciclo consiste en: la sístole y la diástole de las aurículas y los ventrículos.
CAMBIOS DE PRESIÓN Y VOLUMEN DURANTE EL CICLO CARDÍACO
En cada ciclo cardíaco, las aurículas y ventrículos se contraen y se relajan en forma alternada, y transportan la sangre desde las área de mayor presión hacia las de menor presión. A medida que una cámara del corazón se contrae, la presión de la sangre que contiene aumenta. Cada ventrículo, sin embargo, eyecta el mismo volumen de sangre por latido, y ambas cámaras de bombeo siguen el mismo patrón. Cuando la frecuencia cardíaca es de 75 lpm, un ciclo cardíaco dura 0,8s.
Sístole auricular
Durante la sístole auricular , que dura aprox. 0,1 s, las aurículas se contraen. En ese momento, los ventrículos están relajados.
por la onda P del ECG.
se contrae, ejerce presión sobre la sangre contenida en su interior y la impulsa hacia los ventrículos a través de las válvulas AV abiertas.
existente en cada ventrículo (aprox. 105 mL). El final de la sístole auricular también es el final de la diástole ventricular (relajación). Por lo tanto, cada ventrículo contiene 130 mL de sangre, al final de su período de relajación (diástole). Este volumen se denomina volumen de fin de diástole (VFD).
Sístole ventricular
La sístole ventricular se extiende 0,3 s, durante los cuales los ventrículos se están contrayendo. Al mismo tiempo, las aurículas están relajadas, en la diástole auricular.
ventricular comienza, la presión en el interior de los ventrículos aumenta e impulsa la sangre contra las válvulas auriculoventriculares (AV), forzando su cierre. Durante aproximadamente 0,05 s, tanto las válvulas semilunares (SL) como las auriculoventriculares (AV) se encuentran cerradas. Éste es el período de contracción isovolumétrica.
dentro de dichas cámaras. Cuando la presión del ventrículo izquierdo sobrepasa la presión aórtica (80 mm Hg aprox.) y la presión del ventrículo derecho se eleva por encima de la presión del tronco pulmonar (20 mm Hg), ambas válvulas semilunares (SL) se abren. En este punto, comienza la eyección de la sangre desde el corazón. El período en el que las válvulas SL están abiertas es el de la eyección ventricular , que dura aprox. 0,25 s. La presión en el ventrículo izquierdo continúa aumentando hasta 120 mm Hg, mientras que la del ventrículo derecho llega a 25-30 mm Hg.
derecho eyecta el mismo volumen en el tronco pulmonar. El volumen remanente que queda en cada ventrículo al final de la sístole, de aprox. 60 mL, es el volumen de fin de sístole (VFS) o volumen residual. El volumen sistólico (VS o descarga sistólica), volumen eyectado en cada latido por cada ventrículo, es igual a la diferencia entre el volumen de fin de diástole y el volumen de fin de sístole:
En reposo el, el volumen sistólico es de 130 mL – 60 Ml = 70 mL.
Período de relajación
Durante el período de relajación , que dura 0,4 s, tanto las aurículas como los ventrículos están relajados. A medida que el corazón late más rápido, el período de relajación se hace más corto, mientras que la duración se la sístole auricular y ventricular solo se acorta levemente.
ventrículos se relajan, la presión dentro de las cámaras cae, y la sangre contenida en la aorta y en el tronco pulmonar comienza a retornar hacia las regiones de menor presión en los ventrículos. Este pequeño volumen de sangre que refluye produce el cierre de las válvulas SL. La válvula aórtica se cierra a una presión de 100 mm Hg. El choque de la sangre que refluye contra las valvas cerradas de la válvula aórtica produce la onda dícrota en la curva de presión aórtica. Después de que las válvulas SL se cierra, existe un pequeño intervalo en el que el volumen ventricular no varía debido a que todas las válvulas se encuentran cerradas. Este es el período de relajación isovolumétrica.
Cuando la presión ventricular cae por debajo de la presión de las aurículas, las válvulas AV se abren y comienza el llenado ventricular. La mayor parte del llenado ventricular ocurre justo después de la apertura de las válvulas AV. En ese momento, la sangre que ha estado llegando a la aurícula durante la sístole ventricular, ingresa rápidamente a los ventrículos. Al final del período de relajación, los ventrículos han llegado a las tres cuartas partes de su volumen de fin de diástole. La onda P aparece en el ECG y señala el comienzo de otro ciclo cardíaco.
RUIDOS CARDÍACOS
sístole siguiente. En reposo, la descarga sistólica es el 50-60% del volumen de fin de diástole, debido a que un 40-50% de la sangre permanece en los ventrículos luego de cada contracción (volumen de fin de sístole). Tres factores regulan el volumen sistólico y aseguran que los ventrículos derecho e izquierdo bombeen el mismo volumen de sangre:
contraerse.
individuales.
sangre de los ventrículos pueda producirse.
Precarga: efecto del estiramiento
Una precarga mayor en las fibras musculares cardíacas antes de la contracción. Dentro de ciertos límites, cuanto más se llene de sangre el corazón durante la diástole, mayor fuerza de contracción ejercerá durante la diástole. Esta relación se conoce con el nombre de ley de Frank-Starling del corazón. La precarga es proporcional al volumen de fin de diástole (VFD). Normalmente, a mayor VFD, mayor es la fuerza que desarrollará el corazón en la siguiente contracción. Dos son los principales factores determinantes del VFD:
Cuando aumenta la frecuencia cardíaca, la diástole se acorta. Menor tiempo de llenado significa menor VFD, y los ventrículos se contraen antes de que estén adecuadamente llenos. Por el contrario, cuando aumenta el retorno venoso, llega un mayor volumen de sangre a los ventrículos, y aumenta el VFD. Cuando la FC es mayor de los 160 lpm, el VS suele descender debido al acortamiento de la fase de llenado. Las personas que presentan menor frecuencia cardíaca en reposo suelen tener mayor descarga sistólica, ya que su tiempo de llenado es más prolongado, y la precarga aumenta. La ley de Frank-Starling del corazón permite igualar la eyección de los ventrículos derecho e izquierdo y mantener el mismo volumen de sangre fluyendo en ambas circulaciones: sistémica y pulmonar. Si el lado izquierdo del corazón bombea más sangre que el lado derecho, el volumen de sangre que retorna al ventrículo derecho aumenta. El incremento del VFD provoca una contracción del ventrículo derecho más intensa en el latido siguiente, luego del cual se establece nuevamente el equilibrio entre ambos ventrículos.
Contractilidad
Los agentes que incrementan la contractilidad se denominan agentes inotrópicos positivos y aquellos que la disminuyen agentes inotrópicos negativos. Los agentes inotrópicos positivos promueven la entrada de Ca2+ a la fibra muscular cardíaca durante los potenciales de acción, lo que aumenta la fuerza en la próxima contracción. La estimulación de la división simpática del sistema nervioso autónomo por medio de adrenalina y noradrenalina, incrementa el nivel de Ca2+ en el líquido intersticial y los digitálicos poseen efecto inotrópico postivo. Por el contrario, la inhibición de la división simpática del SNA, la anoxia, la acidosis, algunos anestésicos y un aumento de los niveles de K+ en el líquido intersticial tienen efecto inotrópico negativo. Los bloqueantes de los canales de Ca2+ son fármacos que pueden tener efecto inotrópico negativo al reducir la entrada de Ca2+ y, por lo tanto, disminuir la fuerza del latido.
Poscarga
Poscarga en VI equivale a la resistencia vascular periférica o sistémica. En VD equivale a resistencia vascular pulmonar.
La eyección de la sangre por el corazón comienza cuando la presión en el ventrículo derecho excede la presión del tronco pulmonar, y cuando la presión en el ventrículo izquierdo excede la de la aorta. En ese punto, la presión elevada de los ventrículos hace que la sangre empuje las válvulas semilunares y las abra. La presión que debe sobrepasarse para que una válvula semilunar puede abrirse se denomina poscarga. Cuando la poscarga aumenta, el corazón tiene más dificultad para impulsar la sangre y entonces el VS disminuye y permanece más sangre en el ventrículo al finalizar la sístole. Dentro de las condiciones que aumentan la poscarga, encontramos la hipertensión y la disminución del calibre de las arterias por arterosclerosis.
REGULACIÓN DE LA FRECUENCIA CARDÍACA
El gasto cardíaco o volumen minuto depende de la frecuencia cardíaca y del volumen sistólico. Los cambios en la frecuencia cardíaca son importantes en el control a corto plazo del volumen minuto y de la presión arterial. El nodo SA inicia la contracción y establecería una frecuencia cardíaca de 100 lpm. Sin embargo, los tejidos requieren diferente volumen de flujo sanguíneo, de acuerdo con las diferentes condiciones. Durante el ejercicio el gasto cardíaco aumenta para proporcionar a lo tejidos activos mayores niveles de oxígeno y nutrientes. El volumen sistólico puede disminuir, si el miocardio ventricular está dañado o si el volumen sanguíneo disminuye por hemorragia. En estos casos, los mecanismos homeostáticos se encargan de mantener un gasto cardíaco adecuado por medio de aumento de la FC y de la contractilidad. Entre los muchos factores que contribuyen a la regulación de la FC, el sistema nervioso autónomo y la hormonas liberadas hacia la circulación mediante la glándula suprarrenal (adrenalina y noradrenalina) son los más importantes.
Regulación autónoma de la frecuencia cardíaca
El centro cardiovascular del bulbo raquídeo es el principal sitio de regulación nerviosa de la actividad cardíaca. Esta región recibe aferencias de numerosos receptores sensoriales y centros cerebrales superiores, como el sistema límbico y la corteza cerebral. El centro cardiovascular regula la función cardíaca por medio del aumento o disminución de la frecuencia de descarga de impulsos nerviosos en las ramas simpática y parasimpática del SNA. Cuando la actividad física comienza, los proprioceptores que monitorizan la posición de los miembros y músculos, aumentan la frecuencia de los impulsos nerviosos enviados al centro cardiovascular. La aferencia proprioceptiva es uno de los principales estímulos que producen aumento de la frecuencia cardíaca al iniciar la actividad física. Otros receptores sensoriales que proveen aferencias al centro cardiovascular son los quimiorreceptores que controlan los cambios químicos en la sangre, y los barorreceptores , que se encargan de monitorizar el grado de estiramiento de las paredes de los grandes vasos producido por la presión del flujo sanguíneo. Barorreceptores importantes localizados en el arco aórtica y las carótidas detectan los cambios de la presión arterial y los informan al centro cardiovascular. Las neuronas simpáticas van desde el bulbo raquídeo hasta la médula espinal. Desde la región torácica de la médula espinal, los nervios simpáticos cardíacos estimuladores se dirigen hacia el nodo SA, el nodo AV y la mayor parte del miocardio. La estimulación de dichos nervios produce la liberación de noradrenalina que se une a receptores β 1 presentes en las fibras musculares cardíacas. Esta interacción produce 2 efectos distintos:
Durante una cirugía cardíaca, es útil disminuir la FC por medio de hipotermia , que enfría el cuerpo del paciente. Esto provoca un descenso del metabolismo y, por consiguiente, una reducción del consumo de oxígeno, lo que permite que el corazón y el cerebro puedan soportar cortos períodos de interrupción del flujo sanguíneo durante el procedimiento.