¡Descarga transcripcion y más Transcripciones en PDF de Bioquímica solo en Docsity!
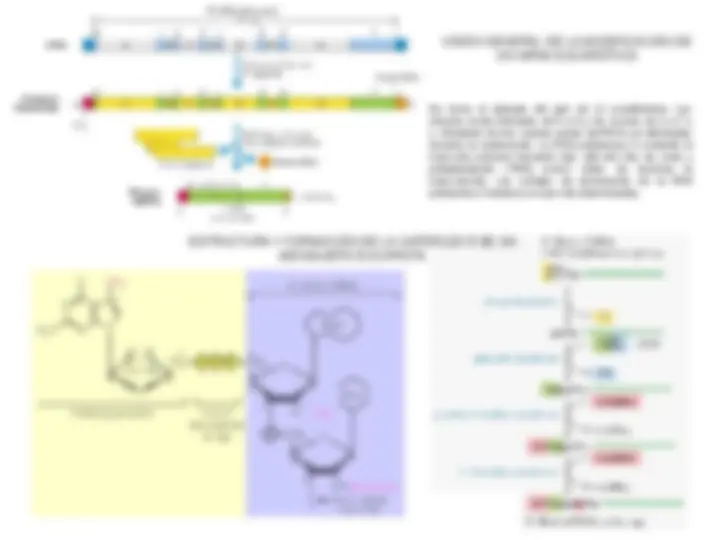
REACCIÓN DE LA POLINUCLEÓTIDO FOSFORILASA REACCIÓN DE UNA RNA POLIMERASA DIRIGIDA POR DNA
COMPOSICIÓN DE SUBUNIDADES DE LA RNA POLIMERASA DE E.coli
ALGUNOS INHIBIDORES DE LA TRANSCRIPCIÓN
La rifampicina es un inhibidor de la iniciación de la transcripción en las bacterias y la -amanitina es un inhibidor de las RNA polimerasas eucariotas. La cordicepina es un terminador de la transcripción ya que la posición 3’ del azúcar está ocupada por H (en rojo), en vez de por un grupo hidroxilo. El sistema de anillo tetracíclico del a actinomicina (en azul) se intercala entre pares de bases G-C adyacentes en el DNA. Los grupos R de la molécula (en rojo) son polipéptidos cíclicos y ocupan los surcos estrechos de la hélice.
n rNDP ⇄ (rNMP)n + nPi
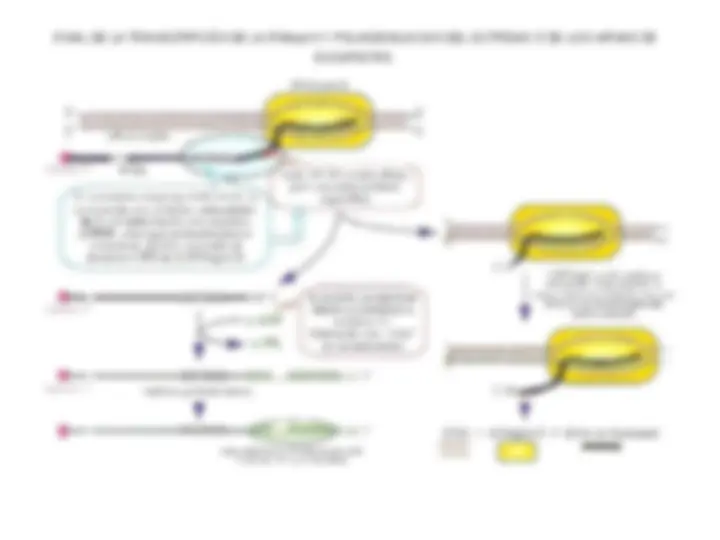
PASOS DE INICIACIÓN Y ELONGACIÓN DE LA
TRANSCRIPCIÓN POR LA RNA POLIMERASA BACTERIANA
BURBUJA DE TRANSCRIPCIÓN
Se han calculado las longitudes del DNA desenrollado y del híbrido DNA-RNA a partir de las reactividades de los complejos de transcripción con reactivos como el KMnO 4 , que oxida las bases en los ácidos nucleicos de cadena simple. Mediante la técnica de huellas dactilares, se determina la longitud del DNA en contacto con la enzima. Seis o siete nucleótidos del RNA detrás del DNA híbrido están protegidos del ataque de la ribonucleasa mediante su unión a la enzima. (nt = nucleótido).
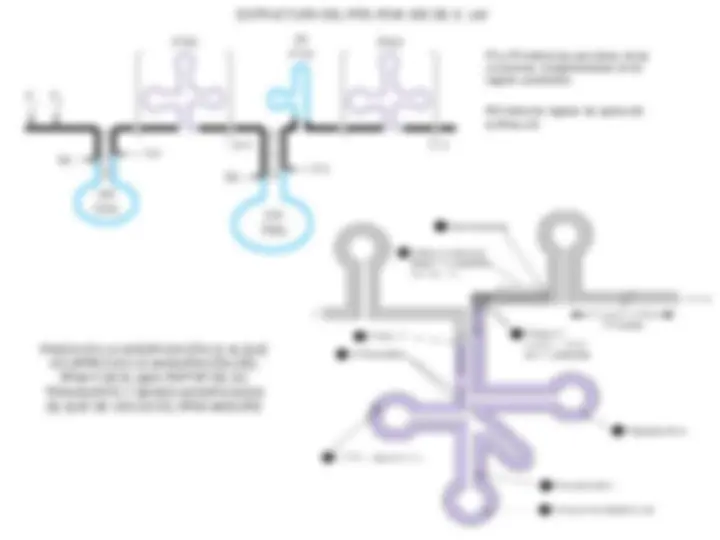
SECUENCIAS CONSERVADAS DE LOS
PROMOTORES RECONOCIDOS POR LA RNA
POLIMERASA DE E. coli
PROPIEDADES DE LAS RNA POLIMERASAS EUCARIOTAS
TRANSCRIPCIÓN POR LA RNA POLIMERASA I Y
PROCESAMIENTO DE LOS RNA RIBOSÓMICOS
PRINCIPALES EN LOS EUCARIOTAS
FACTORES IMPLICADOS EN LA TRANSCRIPCIÓN
DEL rRNA 5S POR LA RNA POLIMERASA III
Los genes se encuentran en copias en tándem, separados por espaciadores que no se transcriben. (a) Los transcritos 45 S producidos inicialmente se procesan mediante la eliminación de las porciones que se indican de color marrón, para dar los productos 18 S, 5.8 S y 28 S. A continuación, se ensamblan en subunidades ribosómicas, mediante la adición de proteínas.
Al menos los tres factores proteicos que se muestran junto con la RNA polimerasa III, deben ensamblarse en el gen antes de que pueda producirse la transcripción. El TFIIIA debe unirse al gen antes de que puedan unirse los factores TFIIIC y TFIIIB. Una vez formado el complejo II estable, se reciclará con pol III para producir muchas copias de RNA. Un exceso de rRNA 5 S formará un complejo con el TFIIIA, inhibiendo la ulterior transcripción.
MODELO DE LA FORMACIÓN DEL COMPLEJO MÍNIMO DE PREINICIACIÓN (PIC)
PARA LA RNA POLIMERASA II SOBRE UN PROMOTOR TATA
(a) El ensamblaje secuencial de un complejo consistente en TBP (a menudo con TFIIA), TFIIB más RNApol II, TFIIE, TFIIF y TFIIH da lugar a un complejo cerrado. TBP se une a menudo como una parte de un complejo mayor denominado TFIID. Algunas de las subunidades de TFIID juegan un papel en la regulación de la transcripción. El DNA es desenrollado en la región Inr por las actividades helicasa de TFIIH y tal vez de TFIIE, que crean el complejo abierto. El dominio carboxilo-terminal de la subunidad mayor de la RNApol II (CTD) es fosforilado por TFIIH (serie de puntos), y entonces la polimerasa abandona el promotor y empieza la transcripción. La elongación está acompañada de la liberación de muchos factores de transcripción y también es potenciada por factores de elongación. Después de la terminación, la RNApol II es liberada, desfosforilada y reciclada.
REPRESENTACIÓN ESQUEMÁTICA DE CÓMO LOS BUCLES DE DNA PUEDEN PONER A LAS PROTEÍNAS
ACTIVADORAS (Act) O REPRESORAS UNIDAS AL POTENCIADOR (ENH) EN CONTACTO CON LAS TAF ASOCIADAS
CON EL COMPLEJO CENTRAL
ACETILACIÓN DE LAS HISTONAS CENTRALES
Estructura general de una histona central
Acetilación, en los lugares indicados en rojo, de los dominios N-terminal
La estructura general de cada una de las cuatro histonas centrales comporta un dominio helicoidal “plegado de histona” más un dominio desestructurado N- terminal muy básico. La acetilación en los núcleos se produce exclusivamente en los dominios N-terminal, en los lugares muy conservados que se indican en rojo.
FINAL DE LA TRANSCRIPCIÓN DE LA RNApol II Y POLIADENILACION DEL EXTREMO 3’ DE LOS mRNAS DE
EUCARIOTAS
VISIÓN GENERAL DE LA MODIFICACIÓN DE
UN mRNA EUCARIÓTICO
Se toma el ejemplo del gen de la ovoalbúmina. Los intrones están indicados de A a G y los exones de 1 a 7 y L. Alrededor de tres cuartas partes del RNA son eliminadas durante la maduración. La RNA polimerasa II extiende el transcrito primario bastante más allá del sitio de corte y poliadenilación (“RNA extra”) antes de terminar la transcripción. Las señales de terminación de la RNA polimerasa II todavía no han sido determinadas.
ESTRUCTURA Y FORMACIÓN DE LA CAPERUZA 5’ DE UN
MENSAJERO EUCARIOTA
MECANISMO QUÍMICO DEL CORTE Y EMPALME DEL mRNA
Los exones (E1 y E2) están indicados por líneas rojas , el intrón por una línea negra o la secuencia. El lugar de corte y empalme E1, posiblemente con la ayuda del snRNP U1, se aparea con una secuencia en la zona de ramificación para formar un bucle. El hidroxilo 2’ del lugar A de la rama reacciona con un residuo G (azul) en el extremo 3’ del exón 1 (el lugar de corte y empalme de E1). Esto libera la G adyacente (rojo) para transferirla al extremo 5’ del exón siguiente (E2). Los productos son un mensaje vuelto a empalmar y una estructura de lazo de un intrón en bucle que luego se degrada.
SECUENCIAS REPRESENTATIVAS EN LOS LUGARES DE
CORTE Y EMPALME
FUNCIONES DEL DOMINIO CTD DE LA RNA pol II EN LA MADURACIÓN DE LOS mRNAs
SÍNTESIS DE LA CAPERUZA 5’.
La síntesis de la caperuza 5’ es llevada a cabo por enzimas ancladas a la CTD de la RNA pol II. Tras finalizar la síntesis de la caperuza, ésta permanece atada al CTD a través de un complejo de unión de la caperuza (CBC).
SPLICING DEL MENSAJERO.
La coordinación del splicing y la transcripción proporciona un mecanismo atractivo para mantener próximos los dos extremos de corte. Cuando emerge el primer punto de corte (en 5’) este es unido por un espliceosoma anclado a la región CTD. El siguiente lugar de corte (en 3’) es luego capturado por este complejo cuando pasa, facilitando la yuxtaposición de ambos sitios de corte y el posterior proceso de corte y empalme.