¡Descarga Tresguerres capitulo 59 y más Esquemas y mapas conceptuales en PDF de Nutrición solo en Docsity!
Capítulo 59
Fisiología de la bilis y de la vía biliar
José A. Solís y María Teresa Muñoz
� E STRUCTURA FUNCIONAL DEL HÍGADO
� MECANISMO DE FORMACIÓN DE LA BILIS
� COMPOSICIÓN QUÍMICA DE LA BILIS
� FUNCIONES DE LA BILIS
� MOTILIDAD DE LA VÍA BILIAR
� B IBLIOGRAFÍA
ESTRUCTURA FUNCIONAL DEL
HÍGADO
El estudio de la estructura del hígado muestra que este órgano está preparado para facilitar los intercambios meta- bólicos entre la sangre de los sinusoides y las células hepá- ticas. Éstas están dispuestas de forma que gran parte de su superficie está en estrecho contacto con la sangre (Fig. 59.1 A ). Los hepatocitos se disponen ordenadamente for- mando tabiques (trabéculas) entre los cuales se sitúan los sinusoides. Dos de las seis caras que poseen las células hepáticas se orientan directamente hacia los sinusoides. Son las denominadas paredes basales (Fig. 59.1 B ). Entre esos tabiques celulares circula la sangre que lle- ga al hígado por la porta y por las arterias hepáticas. Para que las células hepáticas puedan tomar de la sangre todo lo que necesitan es preciso que entre ellas y la sangre se interpongan pocas barreras. Por ello, las paredes de los sinusoides son muy finas y carecen de membrana basal. Las células endoteliales que los recubren están reducidas a una fina lámina formada por sus membranas plasmáticas y en las que existen algunas fenestraciones. En lugar de por membrana basal, los sinusoides están rodeados sólo por algunas fibras aisladas de reticulina. Entre los sinusoides y las paredes de los hepatocitos se sitúa el espacio de Disse. Gracias a esas características estructurales, es posible el paso, no sólo de pequeñas moléculas, sino también de pro- teínas e incluso de quilomicrones. En las paredes basales de las células hepáticas y sus extensiones laterales intercelulares (paredes laterales) se sitúan numerosos receptores (de glucoproteínas, inmuno- globulina A, asialoproteína, insulina, glucagón, etc.) y pro- teínas transportadoras que son esenciales para que puedan pasar a las células los metabolitos que llegan con la sangre sinusoidal. En estas membranas hay una Na+/K+-ATPasa que extrae tres Na+^ de las células y los intercambia por dos K+^ de la sangre, que los envía al interior de los hepatocitos (Fig. 59.2). De esta forma se crea un potencial negativo intracelular de -40 mV y un gradiente de sodio (concentra- ción intracelular de sodio de 15 mM). Este gradiente fuer- za la difusión pasiva de otras sustancias al interior de las células. La función del transportador NTCP ( Na/taurocho- late co-transporting polypeptide , polipéptido cotransporta- dor de Na+/taurocolato) de la membrana basolateral se ve favorecida por el gradiente de sodio generado por esa ATPasa. Este transportador es esencial para el paso de sales biliares conjugadas a las células. La familia de transporta- dores OATP ( organic anion transporting polypeptides , polipéptidos transportadores de aniones orgánicos) [OATP, OATP2, OATP-B, OATP8]) interviene en el paso a las células de un gran número de aniones orgánicos (sales biliares no conjugadas, esteroides, eicosanoides, digoxina y otros fármacos, bilirrubina) con independencia del trans- porte de sodio. Este transportador se encuentra acoplado a la salida de los hepatocitos de CO 3 H-^ o de GSH. La entra- da en los hepatocitos de pequeños cationes orgánicos (fár- macos, colina, neurotransmisores monoamínicos, etc.) está
facilitada por el transportador OCT1 ( organic cation trans- porter , transportador de cationes orgánicos). Finalmente, los OAT ( organic anionic transporters, transportadores de aniones orgánicos ) transportan salicilatos, análogos nucle- ósidos y para-aminohipurato. Entre las caras laterales de las células hepáticas que en las trabéculas no están orientadas hacia los sinusoides, existe un espacio de calibre reducido (0.75-1.50 �m), denominado canalículo biliar , que representa el inicio de la vía biliar (Figs. 59.1 B y 59.1 C ). Sus paredes plegadas y con algunas vellosidades están formadas por las membra- nas de las células entre las que se sitúa. Este espacio está cerrado herméticamente y aislado del espacio intercelular y del espacio de Disse por una estructura compleja que se denomina complejo de unión. En él se pueden identificar cuatro componentes, pero el más importante son las unio- nes estrechas ( tight junctions, zonula occludens ) (Fig. 59.1 C ). En éstas se han identificados varias proteínas (ocludina, claudina) que se unen a las homólogas de las células vecinas y a otras presentes en el citoplasma hepa- tocitario (ZO-1, ZO-2, z one occludens ). Las uniones estre- chas son electronegativas, por lo que impiden el paso de aniones orgánicos y proteínas, pero permiten el del agua, pequeños cationes y parcialmente el de cationes orgánicos. La vasopresina, la epinefrina y la angiotensina II pueden aumentar su permeabilidad. Algunas de las sustancias captadas por las células hepáticas (electrólitos, colesterol, sales biliares, bilirrubi- na, fármacos, tóxicos, etc.), modificadas o no por estas células, son vertidas a los canalículos biliares y contribu- yen a generar la bilis hepática. En la excreción biliar de todas estas sustancias juegan un papel decisivo unos trans- portadores situados en la membrana canalicular de los hepatocitos. Todos ellos forman parte de la gran superfa- milia de transportadores ABC ( ATP binding cassette , case- te de unión a ATP), que son capaces de bombear aniones y cationes orgánicos anfifílicos a la bilis contra un gradiente de concentración de entre 100 y 1000 veces. La energía que consumen estas bombas se la aporta el ATP. A esta familia pertenece el transportador MDR1 ( multidrug resis- tance-1 P glycoprotein , glucoproteína de resistencia mul- tifarmacológica 1P), que participa en la excreción de cationes orgánicos, esteroides, fármacos y péptidos hidro- fóbicos. La MDR3 ( multidrug resistance-3 P glycoprotein , glucoproteína de resistencia multifarmacológica 3P) es esencial para la secreción biliar de lecitina, ya que la tras- loca desde la capa interna, citoplásmica, de la membrana canalicular a la externa ( flipasa ). De esta forma se crea una zona superficial rica en lecitina. En presencia de sales biliares, esta zona protruye hacia la luz canalicular hasta desprenderse en forma de vesícula unilamelar de lecitina (Fig. 59.3). A estas vesículas se incorpora el colesterol. Su deficiencia origina el tipo 3 de colestasis intrahepática familiar progresiva. La excreción de sales biliares la reali- za principalmente la BSEP ( bile salt excretory pump , bom- ba exportadora de sales biliares). Esta bomba es deficiente en el tipo 2 de colestasis intrahepática familiar progresiva. La MRP2 ( multidrug resistance-associated protein , prote-
716 F I S I O L O G Í A D E L S I S T E M A D I G E S T I V O
rrente benigna. Se supone que trasloca la fosfatidil-serina desde la capa externa de la membrana canalicular a la interna (Fig. 59.2). Además de los transportadores men- cionados se han identificado en la membrana canalicular intercambiadores cloro/bicarbonato (AE2), canales de clo- ro, Ca2+^ /Mg2+^ -ATPasa y varios transportadores de función aún no conocida (ABCG2, ABCG5, ABCG8).
MECANISMO DE FORMACIÓN
DE LA BILIS
Bilis canalicular
La bilis que fluye por los canalículos biliares es gene- rada por dos mecanismos diferentes. Uno es dependiente
de las sales biliares, mientras que el otro es independiente de éstas.
- Flujo biliar dependiente de los ácidos biliares. El 50% del flujo biliar canalicular y el 33% del flujo biliar total son dependientes de las sales biliares. Se trata de la bilis que se forma como consecuencia del gradiente osmó- tico creado por la secreción activa de las sales biliares. Este gradiente arrastra agua al interior de los canalículos a través de las uniones estrechas. Las sales biliares responsables de este flujo proceden en su mayor parte (95%) de la sangre portal; las sales biliares de la sangre portal proceden, a su vez, del intesti- no. Las sales biliares se encuentran circulando por el lla- mado circuito enterohepático , formado por el hígado, vías biliares, intestino, sangre portal y nuevamente el hígado. Este recorrido lo realizan las sales biliares varias veces al día. A lo largo de cada uno de ellos pueden perderse
718 F I S I O L O G Í A D E L S I S T E M A D I G E S T I V O
Aniones orgánicos Bilirrubina Cationes orgánicos
Sales biliares conjugadas
Unión estrecha
Agua Cationes
Fosfatidil serina
OATP OATP OATP-B
GSH CO 3 H- OCT ATPasa
NTCP
2K+
3Na+
FIC BSEP MDR Lecitina
MDR GSH MRP
ABCG ABCG ABCG
Figura 59.2. Proteínas transportadoras transmembrana en los hepatocitos. En la membrana sinusoidal se identifican: 1) el transportador NTCP de sales biliares conjugadas acoplado a una Na +/K+-ATPasa. 2) Los transportadores OATP, para el trans- porte de aniones orgánicos y la salida de glutatión (GSH) y bicarbonato (CO 3 H–). 3) El transportador OCT1, implicado en el paso a las células de los cationes orgánicos. En la membrana canalicular se localizan las proteínas transportadoras depen- dientes del ATP: 1) BSEP , que excreta a la bilis sales biliares. 2) MDR3 , o flipasa, que trasloca moléculas de lecitina de la capa interna a la capa externa de la membrana. 3) MRP1, que excre- ta los cationes orgánicos absorbidos de la sangre por la OCT1.
- La MRP2 , encargada de excretar los aniones orgánicos cap- tados por las OATP. Este proceso se encuentra facilitado por la excreción de GSH y CO 3 H–. 5) La proteína FIC1 , de función desconocida, se supone que trasloca la serina de la capa exter- na a la interna de la membrana canalicular. También se repre- senta la situación de las uniones estrechas sellando el espacio canalicular.
Transportador MDR
Sales biliares
BSEP
Translocación de lecitina desde membrana interna a externa Secreción de ác. biliares
Protrusión en la membrana canalicular
Sales biliares
Salida de colesterol
Sales biliares
Formación de microvesícula unilamelar
Figura 59.3. Representación esquemática de la secreción de colesterol. A) Traslocación de lecitina en la membrana canali- cular por la proteína MRD3 y excreción de sales biliares por la BSEP. B) Las sales biliares de la bilis fuerzan la formación de una protrusión en la membrana canalicular enriquecida en leci- tina. Sobre las zonas de membrana no enriquecidas en lecitinas, las sales biliares fuerzan la salida de colesterol y su paso a la zona protruyente. C) Microvesícula biliar unilamelar formada por lecitina en la que se han incorporado algunas moléculas de colesterol.
pequeñas cantidades de sales biliares (Fig. 59.4). A pesar de ello, la cantidad de sales biliares en este circuito per- manece estable, ya que éstas pérdidas son inmediatamente compensadas por el hígado, que las sintetiza a partir del colesterol. La captación hepática de sales biliares presentes en la sangre sinusoidal es sumamente eficaz, como lo prueba el que durante un solo paso de sangre por los sinusoides, los hepatocitos de la periferia lobulillar capten entre el 60- 90% de sales biliares que llegan y que sean muy pocas las que queden para ser captadas por los hepatocitos de la región centrolobulillar. Esta eficiente captación de sales biliares se produce gracias a que en la pared basolateral de los hepatocitos se localizan los transportadores NTCP y OATP. Como hemos mencionado más arriba, la NTCP transporta sales biliares conjugadas aprovechando el gra- diente electroquímico generado por la Na +/K+-ATPasa. Por el contrario, los OATP transportan sales biliares no conjugadas y otros aniones orgánicos acoplados a la salida
de CO 3 H-^ y de GSH. Gracias a la actividad de estas bom- bas de sales biliares, las concentraciones de éstas en las células hepáticas pueden llegar a ser hasta 10 veces supe- riores a las existentes en la sangre. La captación hepática de sales biliares es más eficaz (80-90%) para las más hidrofílicas (conjugadas y trihidroxiladas) que para las sales biliares menos hidrofílicas (50-60%) (no conjugadas, mono- o dihidroxiladas) (Fig. 59.5). La actividad de este sistema puede ser estimulada por la insulina, alfa-adrenér- gicos, glucagón, corticoides y otras hormonas y frenada por el etinil-estradiol y por fármacos tales como la clor- promazina. Una vez dentro de las células, las sales biliares son transportadas hacia el polo biliar por la glutatión-S-trans- ferasa y con la participación de microfilamentos y del apa- rato de Golgi. Durante su travesía intracelular, las sales biliares son conjugadas con la glicina o la taurina y a ellos se suman las sales biliares que están siendo sintetizadas a partir del colesterol.
F I S I O L O G Í A D E L A B I L I S Y D E L A V Í A B I L I A R 719
Sangre
Ácido biliar Via biliar
Colesterol
Vena porta
terminalÍleon Pérdida fecal
Figura 59.4. Circuito enterohepático de las sales biliares. Los áci- dos biliares secretados con la bilis son reabsorbidos en su mayor parte a lo largo del intestino delgado y en especial en el íleon ter- minal. Estos ácidos llegan de nuevo al hígado por la vena porta y, tras su conjugación, son nuevamente secretados para seguir repitiendo este ciclo. Sólo una pequeña cuantía del ácido circu- lante por ese sistema no se absorbe en el intestino y se pierde con las heces o no pasa al hígado y se distribuye por la circulación sistémica. La pérdida fecal es inmediatamente restituida por el hígado que lo sintetiza de novo a partir del colesterol. De esta manera, la cuantía total de ácidos biliares circulantes en ese sis- tema se mantiene constante (2.3 ± 0.43 mg.)
Sales biliares conjugadas
Glicina Taurina
GSH CO 3 H-
OATP
ADP
ATP
NTCP 2K + GSH S-TF
GSH S-TF BSEP H 2 O
H 2 O
Sales biliares no conjugadas
3 Na + Na+/k+-ATPasa -40mV Na +^ 15mM
Figura 59.5. Secreción biliar dependiente de las sales biliares. Las sales biliares pasan a las células para compensar el vacío dejado por el sodio al ser bombeado por la Na+/K+-ATPasa. La actividad de esta enzima crea un gradiente electroquímico entre ambos lados de la membrana basolateral de las células. En el interior de las células la concentración de Na+^ es de 15 mM y el potencial eléctrico de –40 mV. Las sales biliares son trans- portadas por el citoplasma unidas principalmente a la glutatión- S-transferasa (GSH-S-TF). Los transportadores de aniones orgánicos, OATP , captan de la sangre sales biliares no conjuga- das y las envían al citoplasma celular. En este lugar se conjugan con la glicina y la taurina. En la membrana canalicular, el trans- portador BSEP excreta estas sales biliares a la bilis consumien- do la energía aportada por el ATP. La concentración de sales biliares en el canalículo biliar llega a ser 1000 veces superior a la existente en la sangre. Esta diferencia osmolar fuerza el paso de agua desde la sangre a la luz del canalículo biliar, preferen- temente a través de las uniones estrechas.
bios permiten que la bilirrubina se fije a un portador MRP2 y que sea secretada contra un gradiente de concen- tración de hasta 50 veces. Como se ha mencionado más arriba, este transportador de membrana consume la ener- gía aportada por la hidrólisis del ATP. Este portador no es específico de la bilirrubina, sino que ésta lo tiene que com- partir con otros aniones orgánicos y todos ellos compiten entre sí por su uso. La enfermedad de Dubin-Johnson se debe a mutaciones en el gen MRP2 que anulan su función. Las sales biliares facilitan la secreción de bilirrubina, tan- to la conjugada como la no conjugada, probablemente tras incorporarla a micelas mixtas. Esto último es obligado en el caso de la bilirrubina no conjugada, pero no lo es en el caso de la conjugada. Sin embargo, por razones no bien conocidas, ambos tipos de bilirrubina se asocian en la bilis con micelas (véase Fig. 59.7). Se estima que el 60% de la bilirrubina presente en la bilis se excreta incluida en mice- las mixtas. En ese porcentaje figura la totalidad de bilirru- bina no conjugada (1 a 3% de la bilirrubina biliar).
Papel de las vías biliares en la
formación de la bilis
La bilis formada en los canalículos biliares pasa a un sistema de conductos de calibre progresivamente crecien- te que desemboca en la luz del duodeno. Mientras que los canalículos biliares carecen de paredes propias, la luz de los restantes conductos está limitada por células específi- cas que forman sus paredes. Los conductos que forman este sistema y que llevan la bilis hasta el duodeno reciben diferentes nombres (Fig. 59.9). Los más pequeños ( cana- les de Hering ) están formados por unas pocas células apla- nadas en contacto directo con las células hepáticas de la periferia lobulillar. Estos canales se continúan con los con- ductillos biliares o colangiolos (< 15 �m). Éstos se sitúan
F I S I O L O G Í A D E L A B I L I S Y D E L A V Í A B I L I A R 721
(1) Albúmina
OATP1 (2) GSH-S-TF (3) Bilirrubina monoconjugada
(4) Glucuronil transferasa Bilirrubina diconjugada MRP2 (5)
GSH
Bilirrubina diconjugada
Micelas mixtas
Figura 59.7. Secreción biliar de bilirrubina_._ La bilirrubina no conjugada, no soluble en el agua, circula por el plasma unida a la albúmina (1) y pasa a las células con la ayuda del transpor- tador OATP1 (2). En el citoplasma se una a la glutatión-S-trans- ferasa (GSH-S-TF), que lo conduce al retículo endoplásmico (3). En éste, los puentes de hidrógeno se rompen y su molécula se conjuga, en dos pasos sucesivos, con el ácido glucurónico (4). La bilirrubina diconjugada es hidrosolubre y es excretada a la bilis con la intervención de la MRP2. En la bilis puede incorpo- rarse a micelas mixtas.
A
B
C
CH 2
CO 2 H
COOH
HO
OH OH
OH
HO HO C C
COOH
CO 2 H
CH 3 CH
CH 2
CH 2 CH (^2) CH 2 CH 3 CH 3 CH
CH (^2) CH
CH 2 CH 3 CH 3 CH 2
CH 3 CH 2
CH 2 CH 2
CH 2 CH 3 CH 3 CH
N H
N H
N H
N H
N H
N H
N H
N H
0
0 0
0 0
0 0 0 0
0
Figura 59.8. La bilirrubina IXa presente en la bilis humana se forma tras la rotura del puente meteno alfa del Hem. La presen- cia de los radicales propiónicos en los dos anillos pirrólicos centrales permite la formación de seis puentes de H, cuya importancia funcional es decisiva. La molécula adopta una estructura enrollada que oculta los grupos moleculares hidrófi- los. Con ello la bilirrubina IXa pierde solubilidad en el agua. Su transporte por el plasma y a través de las membranas se realiza fijo a proteínas portadoras. En el interior de las células hepáti- cas los puentes de hidrógeno se rompren y los dos radicales propiónicos sirven para su unión a sendas moléculas de ácido glucurónico.
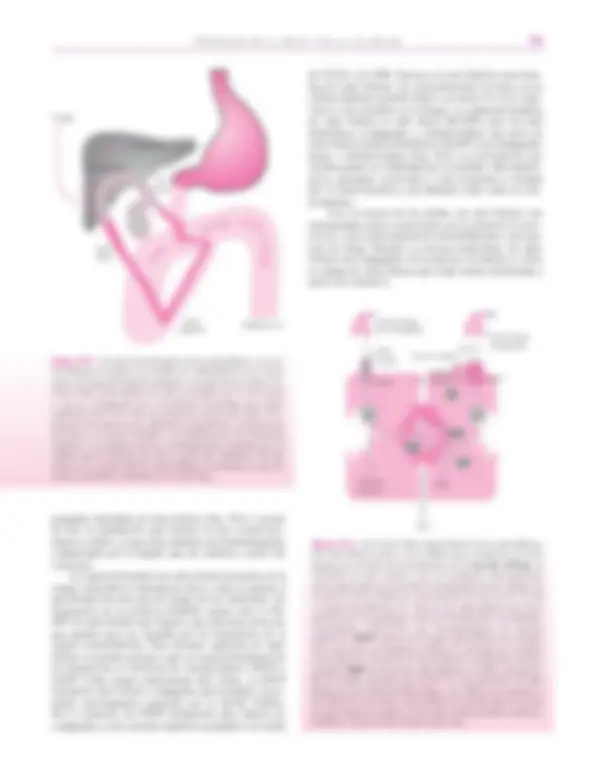
en la periferia de los espacios portales y están formados por 3 ó 4 células cuboides (< 10 �m de altura). En estos espacios y acompañados por una arteriola, se encuentran los conductos biliares interlobulillares (15-100 �m). Sus paredes están formadas por células más altas (10-14 �m), cuboides. La confluencia de varios de estos conductos interlobulillares da lugar a otros de mayor calibre (> 100 �m) ( conductos septales ), recubiertos por epitelio cilíndri- co (> 14 �m). La unión de varios de estos conductos va originando conductos de calibre progresivamente mayor, hasta llegar a la formación de los denominados, primero, conductos segmentarios (400-800 �m) y, más adelan- te, conductos hepáticos. El hígado posee dos conductos hepáticos y cada uno de ellos recoge la bilis procedente de un lóbulo hepático. Todos estos conductos poseen paredes formadas por tejido conjuntivo que están tapizadas por un epitelio cilíndrico que reposa sobre la membrana basal. Los conductos hepáticos confluyen en el conducto hepáti- co común , al cual le sigue el conducto colédoco. Estos conductos están revestidos también por un epitelio cilín- drico que asienta sobre una pared relativamente gruesa for- mada por tejido conjuntivo laxo, rico en fibras elásticas, y por fibras musculares longitudinales y oblicuas. Antes de desembocar en el duodeno, el conducto colédoco y el con- ducto pancreático se unen para drenar juntos en la papila duodenal mayor. El músculo liso de la pared del colédoco se hace más prominente a medida que se aproxima al duo- deno. Cuando el colédoco se interna en la pared del duodeno, su luz se estrecha por verse reforzadas sus capas
musculares por otra nueva capa de fibras musculares circulares. Esta zona es la que se conoce como esfínter de Oddi. En situación lateral al conducto colédoco y a manera de un gran divertículo se sitúa la vesícula biliar , en la cual se almacena la bilis durante los períodos interdigestivos (Fig. 59.9). Su pared está formada por una capa muscular plexiforme, una adventicia y una serosa. La mucosa está plegada con depresiones profundas que pueden penetrar en el espesor de la capa muscular ( criptas de Rokitansky- Aschoff ). Sus células son cilíndricas altas, con núcleo basal oval y citoplasma pálido. La bilis canalicular recién formada en el lobulillo hepático sufre diversas modificaciones a su paso por todo este sistema de conductos, incluida la vesícula biliar. En efecto, la bilis colangiolar aumenta su flujo, su alcalinidad y su contenido en bicarbonato pero pierde sales biliares. En este proceso juega un papel importante la secretina. El estímulo de la mucosa duodenal por el pH ácido, ácidos grasos o sales biliares provoca la liberación de secretina, la cual se une a receptores presentes en la membrana basola- teral de las células de los conductos biliares. Esta unión se sigue de la formación de AMP cíclico (AMPc) con la mediación de una proteína G y de una adenilciclasa. El AMPc tiene varios efectos (Fig. 59.10). Por un lado, acti- va la proteína quinasa A (PKA) y ésta abre unos canales de cloro (CFTR, cystic fibrosis transmembrane regulator , regulador transmembrana de la fibrosis quística) situados en el polo apical (biliar) de la célula. Consecuencia de ello es la salida de cloro a la luz biliar. Este canal no funciona correctamente en la fibrosis quística. La salida del cloro se encuentra acoplada con su intercambio por CO 3 H –. Es decir, el Cl–^ vuelve a entrar en las células intercambiándose por CO 3 H–, el cual sale a la bilis y la alcaliniza. La con- centración de éste aumenta desde 19-25 mM hasta 60 mM. Esta reentrada del Cl –^ la realiza el intercambiador Cl – /CO 3 H–, AE2. La segunda consecuencia de la forma- ción de AMPc es que la acuaporina-1, presente en vesícu- las citoplásmicas subapicales, se incorpora a la membrana apical de las células y contribuye a formar en ésta unos canales que permiten la salida de agua a la bilis. Todos estos canales y el intercambiador se expresan preferente- mente en las células de los conductos biliares de tamaño mediano y grande. En la membrana apical de las células de los conductos se sitúa también: 1) un cotransportador de sodio y glucosa (SGLT1, Na/glucose transporter , cotransportador de Na+^ y glucosa), 2) un transportador de sales biliares dependiente del sodio (iBAT, ISBT, Ileal sodium dependent bile salt transporter ) y 3) un canal de cloro sensible al calcio, no relacionado con el AMPc. El primero capta glucosa y sodio de la bilis, el segundo reabsorbe pequeñas cantidades de sales biliares y de sodio y el último permite la salida de clo- ro de las células, principalmente cuando falla el canal CFTR. En la membrana basolateral de las células de los conductos biliares también se han identificado numerosos transportadores. Los más significativos son: 1) Na+/K+- ATPasa, similar a la existente en los hepatocitos. Genera un
722 F I S I O L O G Í A D E L S I S T E M A D I G E S T I V O
Canalículo biliar Canal de Hering Conductillo biliar (colangiolo) Conducto interlobulillar Conducto septal
Conductos segmentarios
Conductos lobulares
Izquierdo
Conducto hepático común Conducto de Wirsung
Cístico Colédoco
Esfínter de Oddi
Vesícula biliar
Figura 59.9. Vías biliares intrahepáticas y extrahepáticas.
FUNCIONES DE LA BILIS
La bilis posee varias funciones: 1) proporcionar al intestino ácidos biliares que faciliten la absorción de gra- sas y de vitaminas liposolubles, 2) eliminar sustancias endógenas o exógenas, tales como la bilirrubina, metales, fármacos, tóxicos y productos residuales, 3) homeostasis del colesterol y 4) proteger al intestino frente a infecciones mediante la secreción de IgA y citoquinas inflamatorias. De todas las funciones mencionadas, destaca la rela- cionada con la absorción intestinal de grasas y de vitami- nas liposolubles. Para que los lípidos de los alimentos puedan ser absorbidos por el intestino, deben sufrir un pro- ceso que los permita permanecer en solución acuosa y alcanzar la superficie de los enterocitos. Este proceso consta de varias fases: emulsificación, lipólisis, micela- ción, liberación de las micelas y paso a los enterocitos (Fig. 59.12). Los triglicéridos, fosfolípidos, colesterol, vitaminas liposolubles, etc. de los alimentos sufren una primera emulsificación durante la masticación y por las contrac- ciones del estómago. Para la formación de una emulsión fina sobre la que puedan actuar las enzimas pancreáticas (lipasa, fosfolipasa A 2 , colesterol-esterasa) son fundamen- tales los fosfolípidos de los alimentos y las sales biliares y fosfolípidos de la bilis. Todos estos forman una fina pelícu- la que rodea a cada gota de lípidos. Sobre esas microgotas
actúan la lipasa y las restantes enzimas lipolíticas del pán- creas. La fosfolipasa A 2 digiere los fosfolípidos de la superficie de las microgotas y, de esta forma, permite que la lipasa acceda a los triglicéridos de esas gotas. Para que las fosfolipasa A 2 ejerza su función en condiciones ópti- mas es necesaria la presencia de sales biliares y de calcio. La lipasa, con la colaboración de la colipasa pancreática, realiza la digestión parcial de las triglicéridos, liberando ácidos grasos y 2-monoglicéridos. Los productos de esta lipólisis, al igual que otros lípidos (colesterol, vitaminas A, E, D, K), son solubilizados en el agua al ser incorporados a micelas de sales biliares y fosfolípidos. Éstas se forman cuando la concentración de las sales biliares en la luz intestinal supera lo que se denomina “concentración mice- lar crítica” (3 mM). La capacidad de las micelas de sales biliares para solubilizar lípidos aumenta de forma conside- rable cuando en ellas existen también fosfolípidos biliares. Incorporados a estas micelas, los lípidos se aproximan a las células intestinales. La capa de agua que las recubre tiene un pH ligeramente ácido, suficiente para que los áci- dos grasos y 2-monoglicéridos pierdan su solubilidad en las micelas, se liberen de éstas, se protonicen y puedan pasar a las células intestinales. Aunque se ha considerado que este paso se producía por difusión pasiva, cada vez se cuenta con más pruebas de que al menos algunos lípidos (ácidos grasos) se absorben de forma activa con la media- ción de transportadores específicos.
724 F I S I O L O G Í A D E L S I S T E M A D I G E S T I V O
A (^) Glicina
Glicina
Taurina
Taurina
OH
OH
COOH
NI- 2 -CH 2 -COOH NI- 2 -CH 2 -CH 2 -SO 3 H HO Ácido biliar Ácido cólico Micela de ácidos biliares
B
Lecitina
Membrana bicapa
de lecitina Zona hidrófila
Zona hidrófoba
Zona hidrófila
c
HO Colesterol Micela mixta Vesícula biliar
Figura 59.11. Lípidos biliares. A) Representación esquemática de un ácido biliar (ácido cólico) aislado formando micelas, y su forma de conjugación con la glicina y la taurina. B) Fosfolípido (lecitina) y su representación esquemática en forma de membrana bicapa. C) Colesterol y su representación esquemática. Micela mixta constituida por sales biliares, lecitina y colesterol. Vesícula fosfolipídica con moléculas de colesterol en su espesor.
MOTILIDAD DE LA VÍA BILIAR
La llegada de alimentos al duodeno provoca la con- tracción de la vesícula, la relajación del esfínter de Oddi y el aumento del flujo biliar hepático. Estos efectos condu- cen a que la vesícula vacíe el 50-75% de su contenido en el duodeno. El control motor de la vesícula es doble: hor- monal y neural. La principal hormona implicada en la res- puesta vesicular a la ingesta de alimentos es la colecistoquinina (CCK). Se trata de un polipéptido de 33 aminoácidos, producido y liberado por células especializa- das del sistema APUD ( amine precursor uptake and decarboxilation , sistema de captación y descarboxilación de los precursores de las aminas), situadas en la mucosa duodenal. Es liberada en respuesta a la presencia de H +, grasas o aminoácidos en la luz del duodeno, pero su libe- ración se inhibe por la presencia en ese lugar de sales bilia- res o de tripsina. Como respuesta a los alimentos se liberan otras sustancias. Unas actúan como agonistas y otras como antagonistas de la contracción vesicular. Entre las prime- ras figuran la gastrina, la secretina, el estímulo vagal, el simpático �-adrenérgico y la distensión antral. Entre los factores que relajan la pared vesicular figuran la somatos- tatina, el polipéptido pancreático, el polipéptido intestinal vasoactivo (VIP) y el estímulo símpático -adrenérgico. Durante los períodos interdigestivos, se produce el llenado vesicular, lo cual está determinado por la tasa de
secreción biliar y por la resistencia al flujo biliar que opo- ne el esfínter de Oddi. En estas fases, este esfínter está contraído, con una presión de 4 a 15 mm Hg por encima de la presión duodenal, lo que repercute sobre la presión en el conducto colédoco (12 mm Hg). Como la presión en el conducto cístico es de unos 8 mm Hg y la de la vesícu- la de 10 mm Hg, la bilis fluye hacia esta última. En contra de lo que se creía en otros tiempos, la vesícula tiene con- tracciones rítmicas durante las cuales vacía del 20 al 30% de su contenido. Estas contracciones coinciden con el paso
F I S I O L O G Í A D E L A B I L I S Y D E L A V Í A B I L I A R 725
Células intestinales
Capa de agua sobre la superficie intestinal
Emulsión de los lípidos en el estómago
Emulsión de los lípidos en el intestino. Microgotas recubiertas por una fina capa de fosfolípidos y sales biliares en su superficie
Fosfolipasa A 2. Digestión de la capa de fosfolípidos Lipólisis por la colipasa y lipasa pancreáticas
Liberación de ác. grasos y monoglicéridos
Incorporación de lípidos a micelas de sales biliares con o sin fosfolípidos
pH 5-
Figura 59.12. Papel de la bilis en la digestión intestinal de los lípidos.
250
150
(^50 )
150
mEq/L
Bilis vesicular
Bilis hépatica
Na+
Sales biliares
k+ CO 3 H-
Presión vesícula Reposo: 10 mm Hg Contracción: 25 mm Hg Na+ CI-
H 2 O
CO 3 H- CI- 3Na+^
2K+
10 mm Hg
mm Hg
200 150 100 50 Contracciones Colecistoquinina Morfina fásicas basales
Presión esfínter de Oddi 4-15 mm Hg
Figura 59.13. Función del sistema biliar de grandes conduc- tos. La resistencia del esfínter de Oddi permite la retención de la bilis en el conducto colédoco y el llenado vesicular. En ésta se produce la reabsorción de Na +^ , Cl – , K +^ y agua, por lo que el volumen de la bilis disminuye y sus componentes se concen- tran. Estos movimientos electrolíticos están determinados por la presencia en las células epiteliales de un cotransportador Na+/Cl –^ y de dos intercambiadores, uno Cl–/CO 3 H+^ y otro Na+/K +^. La presión en el conducto colédoco suele ser de unos 10 mm Hg, mientras que en el esfínter es de unos 4 a 15 mm Hg por encima de la existente en el duodeno. Sobre esa presión esfinteriana basal, se producen contracciones fásicas (3 a 7 minutos) de 4.3 ± 0.5 segundos de duración, durante las cuales la presión puede ascender entre 100 y 150 mm Hg. La fre- cuencia, amplitud y duración de estas ondas pueden verse influenciadas por diversos factores hormonales.