Baixe Apostila Bio Mol e outras Notas de estudo em PDF para Agronomia, somente na Docsity!
1. ESTRUTURA DO DNA.
A molécula de DNA é constituída por uma seqüência de nucleotídeos, que por sua vez é formado por três diferentes tipos de moléculas:
Um açúcar (pentose)
O açúcar que compõe o DNA é a pentose desoxirribose.
Já, no RNA a pentose é a ribose.
Um grupo fosfato
Uma base nitrogenada
As bases nitrogenadas são derivadas de compostos heterocíclicos que são as purinas ou pirimidinas.
No DNA existem quatro tipos de bases:
Adenina
Timina
Guanina
Citosina
- A ligação entre o grupo fosfato e a pentose:
Esta ligação é feita através de uma ligação fosfoéster com a hidroxila ligada ao carbono-5 da pentose.
Para a formação da molécula de DNA é necessário que ocorra a ligação entre os nucleotídeos.
Os nucleotídeos estão ligados covalentemente por ligações fosfodiéster formando entre si pontes de fosfato.
O grupo hidroxila do carbono-3 da pentose do primeiro nucleotídeo se liga ao grupo fosfato ligado à hidroxila do carbono-5 da pentose do segundo nucleotídeo através de uma ligação fosfodiéster.
Devido a esta formação a cadeia de DNA fica com uma direção determinada, isto é, em uma extremidade temos livre a hidroxila do carbono-5 da primeira pentose e na outra temos livre a hidroxila do carbono-3 da última pentose.
Isto determina que o crescimento do DNA se faça na direção de 5'para 3'.
Sabendo-se como são feitas as ligações entre os nucleotídeos, formando assim a fita de DNA, podemos analisar a estrutura tridimensional do DNA.
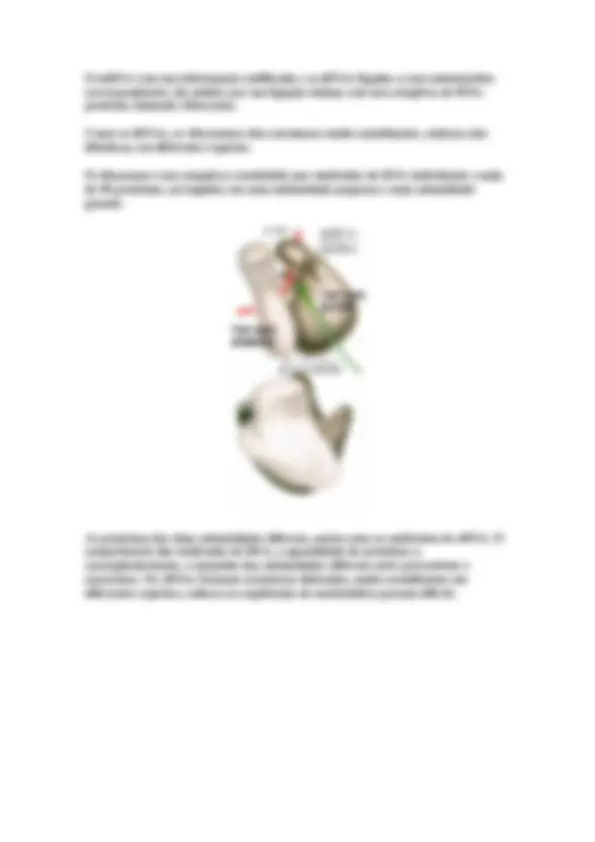
James Watson e Francis Crick postularam um modelo tridimensional para a estrutura do DNA baseando-se em estudos de difração de raios-X.
A relação espacial entre as duas fitas cria um sulco principal e um sulco secundário.
O pareamento das bases de cada fita se dá de maneira padronizada, sempre uma purina com uma pirimidina, especificamente: adenina com timina e citosina com guanina.
A proximidade destas bases possibilita a formação de pontes de hidrogênio, sendo que adenina forma duas pontes de hidrogênio com a timina e a citosina forma três pontes com a guanina.
A dupla hélice é mantida unida por duas forças:
Por pontes de hidrogênio formadas pelas bases complementares
Por interações hidrofóbicas, que forçam as bases a se "esconderem" dentro da dupla hélice.
Estudos recentes mostram que existem duas formas de DNA com a hélice girando para a direita, chamadas A-DNA e B-DNA, e uma forma que gira para a esquerda chamada Z-DNA. A diferença entre as duas formas que giram para a direita está na distância necessária para fazer uma volta completa da hélice e no ângulo que as bases fazem com o eixo da hélice.
B-DNA: Tem a dupla hélice mais longa e mais fina. Para completar uma volta na hélice são necessários 10 pares de bases.
A-DNA: Tem a forma mais curta e mais grossa. Para completar uma volta na hélice são necessários 11 pares de bases.
- O estado superenrolado é menos estável que o estado relaxado. Para desfazer o "supercoil" é necessário uma quebra em uma das cadeias de DNA.
- Em Procariotos, o superenrolamento é feito por um grupo de enzimas denominadas "topoisomerases II", enquanto que o relaxamento é feito pela "topoisomerase I".
- Ambas topoisomerases catalizam a quebra e a religação de ligações fosfodiéster do DNA.
Propriedades Físicas e Químicas do DNA.
Soluções de DNA, em pH = 7,0 e temperatura ambiente, são altamente viscosas;
A altas temperaturas ou pH extremos o DNA sofre desnaturação, isto porque ocorre ruptura das pontes de hidrogênio entre os pares de bases. Esta desnaturação faz com que diminua a viscosidade da solução de DNA;
Durante a desnaturação nenhuma ligação covalente é desfeita, ficando portanto as duas fitas de DNA separadas;
Quando o pH e a temperatura voltam ao normal, as duas fitas de DNA espontaneamente se enrolam formando novamente o DNA dupla fita. Este processo envolve duas etapas:
A primeira é mais lenta pois envolve o encontro casual das fitas complementares de DNA, formando um curto segmento de dupla hélice.
A segunda etapa é mais rápida e envolve a formação das pontes de hidrogênio entre as bases complementares reconstruindo a conformação tridimensional.
2. REPLICAÇÃO DO DNA.
O DNA é a molécula que transmite a informação genética e tem a capacidade de se auto-duplicar. Uma cópia do DNA é passado para a célula-filha durante a divisão celular. A este processo de auto-duplicação do DNA deu-se o nome de replicação.
Replicação do DNA é o processo de auto-duplicação do material genético mantendo assim o padrão de herança ao longo das gerações.
Surgiram algumas dúvidas a respeito da replicação do DNA:
Quando a replicação se completa, tem-se a fita velha pareada com a fita nova ou a fita velha com a velha e a nova com a nova?
O DNA sofre replicação semi-conservativa.
Duas teorias tentaram explicar a replicação do DNA:
- Teoria conservativa: Cada fita do DNA sofre duplicação e as fitas formadas sofrem pareamento resultando num novo DNA dupla fita, sem a participação das fitas "parentais" (fita nova com fita nova formam uma dupla hélice e fita velha com fita velha formam a outra dupla fita).
- Teoria semi-conservativa: Cada fita do DNA é duplicada formando uma fita híbrida, isto é, a fita velha pareia com a fita nova formando um novo DNA.
Estas hipóteses foram testadas pelo experimento de Meselson-Stahl.
Como se dá à replicação propriamente dita?
Experimento:
Células de E.coli crescidas por várias gerações, num meio onde a única fonte de nitrogênio era cloreto de amônio (NH4Cl) com a substituição do nitrogênio normal (14N ) por nitrogênio "pesado" (15N ).
Desta forma, todos os componentes nitrogenados das células que cresceram neste meio, incluindo as bases do DNA, estariam enriquecidos por nitrogênio "pesado". A densidade dos componentes das células crescidas em 15N é maior do que a dos componentes das células crescidas em meio "normal" (14N) , no caso o DNA das células "parentais".
A análise do experimento foi feita por ultra-centrifugação utilizando uma solução concentrada de cloreto de césio.
John Cairns evidenciou por auto-radiografias a presença de "olhos" ou "bolhas de replicação" nos cromossomos circulares de bactérias crescidas em meio de cultura contendo [3H] - timidina.
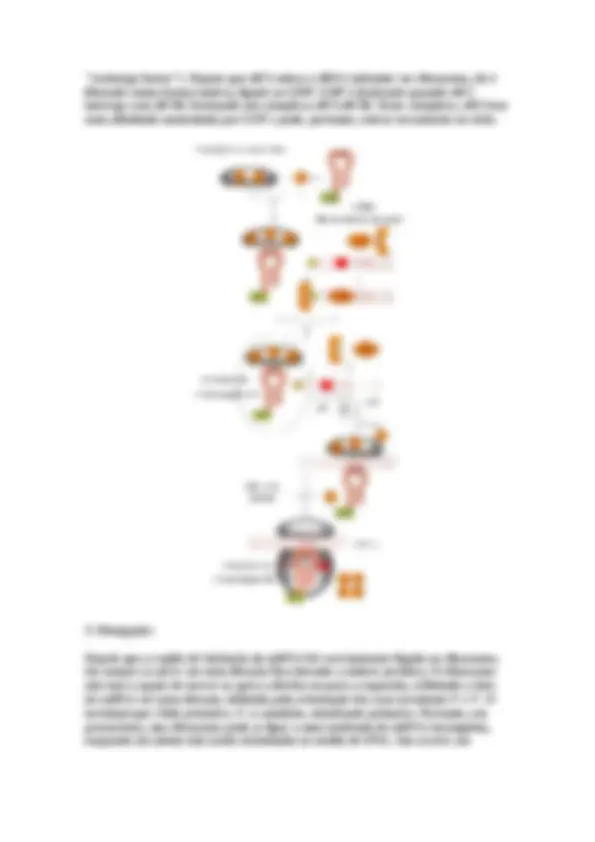
A análise indica que o DNA dupla hélice se replica por um processo progressivo de separação das duas fitas parentais, acompanhada pela síntese de uma nova cadeia complementar a este DNA (fita filha).
A extremidade onde está a "bolha" de replicação é chamada de forquilha de replicação.
Uma "bolha" de replicação pode ter uma ou duas forquilhas. Se tiver apenas uma forquilha significa que a replicação é unidirecional e se tiver duas forquilhas a replicação é bidirecional. Com base nos estudos auto-radiográficos, demonstrou-se que a replicação do DNA é bidirecional (uma "bolha" de replicação com duas forquilhas que se movimentam em sentidos opostos).
Em procariotos e bacteriófagos, o DNA tem apenas uma "bolha" de replicação, enquanto que em eucariotos existem várias "bolhas" de replicação que estão distribuídas ao longo do DNA.
A tensão formada pelo "andamento" da forquilha de replicação ao longo do DNA é solucionada pela ação de algumas enzimas.
DNA girase
À medida que a forquilha de replicação "anda" pela dupla fita de DNA, começa a aparecer uma tensão na porção logo à frente da forquilha, devido ao "supercoil" do DNA.
Para aliviar esta pressão do "supercoil" um grupo de enzimas, denominadas de topoisomerases, quebram momentaneamente o DNA, fazendo com que este sofra algumas rotações e em seguida este mesmo grupo de enzimas refazem a ligação "quebrada".
Em procariotos estas topoisomerases são conhecidas como DNA girase. Este processo de desenrolamento do DNA é extremamente importante em procariotos.
Antibióticos do tipo ovobiocina e ácido oxolínico inibem a ligação da DNA girase ao DNA. O emprego destes inibidores levam a formação de mutantes.
Como ocorre a replicação sendo as fitas são anti-paralelas?
A replicação do DNA é semi-descontínua.
Sabemos que as fitas do DNA são anti-paralelas (uma fita tem direção de 5’ para 3’ enquanto que a outra tem a direção de 3’ para 5’).
- atividade exonucleásica de 3´ para 5´: retira os nucleotídeos de DNA que possam estar errados.É importante para os mecanismos de reparo do DNA.
- atividade exonucleásica de 5´para 3´: retira os nucleotídeos que fazem parte dos "primers" de RNA.
DNA polimerase II:
Ainda não tem suas funções bem definidas.
DNA polimerase III:
Enzima que forma os fragmentos de Okazaki. Possui atividade polimerásica e exonucleásica. Ocorre em baixa concentração na célula.
A DNA polimerase III faz parte de um grande complexo enzimático constituído por sete subunidades: a, e, q, t, g, d e b.
A DNA polimerase III propriamente dita é formada por três destas subunidades: a, b e e.
- Subunidade a: somente esta subunidade tem ação polimerásica e exonucleásica de 5’para 3’.
- Subunidade b: liga a DNA polimerase III ao molde do "primer" e este processo requer a hidrólise de ATP.
- Subunidade e: tem ação exonucleásica de 3’ para 5’. As outras subunidades, do complexo enzimático, são todos necessários à função biológica da enzima.
Replicação da fita descontínua:
É importante sempre ter em mente que:
- A replicação da fita descontínua é feita pela DNA polimerase, portanto na direção de 5’para 3’.
- Todas as DNA polimerases requerem, para a produção da fita de DNA, a hidroxila da extremidade 3’ livre.
Com ênfase na fita descontínua, experimentos foram desenvolvidos a fim de se conhecer o início da síntese desta fita já que a porção livre é a hidroxila 5’.
A análise dos fragmentos de Okazaki revelou que a porção final deste fragmento consiste de um curto segmento de RNA ("primer" de RNA) e o restante do fragmento é composto por DNA.
A DNA polimerase I possui atividade exonucleásica e polimerásica. Em sua atividade exonucleásica, agindo na direção de 5’para 3’, remove nucleotídeo por nucleotídeo do "primer" de RNA e à medida que estes nucleotídeos vão sendo removidos a DNA polimerase I, em sua atividade polimerásica, vai adicionando desoxirribonucleotídeos completando o fragmento com moléculas de DNA.
Os segmentos formados, agora compostos apenas de moléculas de DNA, são ligados entre si covalentemente formando a nova fita de DNA.
Esta ligação é feita por uma enzima chamada de DNA ligase.
A DNA ligase forma uma ligação fosfodiéster entre o grupo OH - 3’ na extremidade de um segmento com o grupo OH - 5’ do próximo segmento, liberando pirofosfato.
DNA Ligase
É uma enzima que faz as ligações covalentes entre os fragmentos de Okazaki formando assim a fita descontínua.
Quadro – Enzimas de Replicação:
DNA polimerases
Existem três tipos de DNA polimerases em procariotos:
- DNA polimerase I
- DNA polimerase II
- DNA polimerase III
Procariotos I II III Atividade polimerásica 5’--3’ + + + Atividade exonucleásica 3’--5’ + + + Atividade exonucleásica 5’--3’ + - - Moléculas presentes por célula 400? 10-
Existem quatro tipos de DNA polimerases em eucariotos:
- DNA polimerase a
- DNA polimerase b
- DNA polimerase g
- DNA polimerase e
Eucariotos a b g d Atividade polimerásica 5’--3’ + + + + Atividade exonucleásica 3’--5’ + - - + Localização na célula núcleo núcleo mitocôndria núcleo Associado a DNA primase + - - -
Para que haja economia da enzima DNA polimerase I e de tempo a fita descontínua sofre uma torção, que chamamos de "splicing"
O "splicing" é formado de modo que uma única DNA polimerase I replique, simultaneamente, as duas fitas de DNA.