Baixe Aula 03 - Núcleo interfásico e outras Notas de estudo em PDF para Biologia Celular, somente na Docsity!
Núcleo interfásico a 3
u
l^
a
OBJETIVOS (^) • Discutir a importância do aparecimento
de um núcleo individualizado.
- Listar os componentes estruturais do núcleo
durante a intérfase.
- Enumerar os diversos graus de organização
da cromatina.
- Correlacionar aparência, composição
e função do nucléolo.
Pré-requisitos
Compartimentalização celular
(Aula 15 de Biologia Celular I).
Controle do ciclo celular (Aula
1 de Biologia Celular II).
Divisão celular (Aula 2 de
Biologia Celular II).
BIOLOGIA CELULAR II | Núcleo interfásico
INTRODUÇÃO Durante a intérfase, quase todo o genoma de um eucarioto está dentro de um compartimento, o núcleo. O núcleo é o maior compartimento celular, ocupando em média 10% do volume total da célula. A maioria das células possui apenas um núcleo, com exceção de:
- eritrócitos de mamíferos, que perdem o núcleo durante a diferenciação;
- células derivadas da fusão de células precursoras, como as células musculares esqueléticas, que possuem muitos núcleos;
- certos protozoários, como os do gênero Giardia , que são binucleados ( Figura 3.1 ).
Figura 3.1 : (A) Célula muscular multinucleada; (B) o protozoário Giardia lamblia possui dois núcleos e quatro pares de flagelos.
O núcleo costuma ter formato arredondado e ocupar posição central nas células, mas pode ficar na periferia, como nas células musculares ( Figura 3.1 ) ou nos adipócitos (reveja a Figura 27.2 em Biologia Celular I) e não serem redondos, como os núcleos de leucócitos ( Figura 3.2 ).
IMPORTÂNCIA EVOLUTIVA
Você aprendeu em Biologia Celular I (Aula 15) que a compartimenta- lização foi um grande avanço evolutivo porque permitiu que as diferentes funções celulares pudessem ser distribuídas em diferentes locais, onde as moléculas envolvidas em determinada atividade celular estão próximas. Esse raciocínio se aplica perfeitamente ao compartimento nuclear, mas há outras vantagens que tornaram o aparecimento do núcleo evolutivamente tão importante.
Núcleos Miofibrila
Núcleos
Flagelos a b
BIOLOGIA CELULAR II | Núcleo interfásico
Para que você possa avaliar o impacto evolutivo da separação dos eventos de transcrição e tradução, basta lembrar que a quantidade de informação contida no genoma de procariotos é muito menor que a contida no genoma dos eucariotos. Essa diferença não se deve apenas à maior massa de DNA presente neles. Nos procariotos, os genes estão arranjados em seqüência linear. Se toda a diversidade de informação dos eucariotos tivesse de ser arranjada na forma de genes enfileirados, seria impossível acomodar o comprimento de seu genoma numa célula! Foi a oportunidade de processar o RNA resultante da transcrição que gerou a possibilidade de sobrepor informações. Ao ser processada, cada molécula de RNA ora perde uma parte, ora outra parte, transpõe regiões etc., podendo gerar vários mRNA, cada um resultando numa proteína diferente. Assim, as células eucarióticas puderam se tornar muito mais complexas, com uma variedade de proteínas muito maior do que a variedade genômica. Se você tinha aprendido que o aparecimento do núcleo foi evolutivamente importante porque protegeu o DNA, não se preocupe. Esse conceito não está errado. De fato, envolvido por um envelope tão bem estruturado (você já vai ver!), o genoma está protegido de todo o movimento do citoplasma. Mas vamos reconhecer que, se nada mais tivesse acontecido depois do aparecimento do núcleo, a diferença entre eucariotos e procariotos seria muito menor!
A ORGANIZAÇÃO GERAL DO NÚCLEO INTERFÁSICO
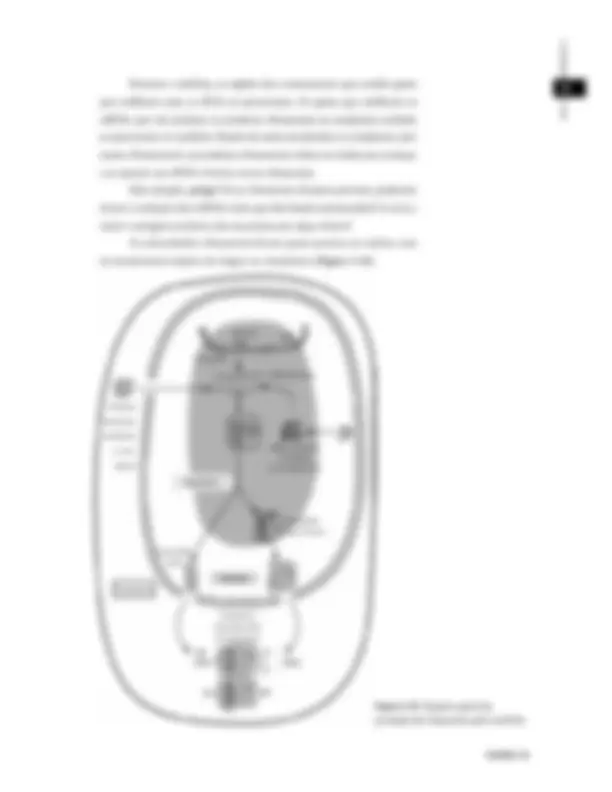
O núcleo é o compartimento que contém o DNA, organizado na forma de cromatina , e está separado do citoplasma por um envelope ou envoltório nuclear. O envoltório define um ambiente nuclear, cuja matriz é diferenciada, o nucleoplasma. O nucleoplasma e o citoplasma se comunicam diretamente através de aberturas no envoltório. Essas aberturas não são simples buracos, e sim poros muito bem estruturados, denominados complexos do poro. Partes diferentes da cromatina têm arranjos especiais, sendo o nucléolo o mais conhecido deles. O envoltório nuclear, formado por duas membranas, está sustentado tanto pelo lado citoplasmático quanto pelo lado nuclear, por filamentos do citoesqueleto ( Figuras 3.4 e 3.5 ). No lado nuclear, filamentos intermediários formam a lâmina nuclear.
AULA
3
No lado citoplasmático, outros tipos de filamentos intermediários, MÓDULO 1^ além de microtúbulos, colaboram com a sustentação. O próprio centro organizador de microtúbulos (centrossomo) se encontra bastante próximo do envoltório nuclear. Vamos dedicar atenção especial a cada uma dessas principais estruturas nucleares nesta e na próxima aula.
Figura 3.4 : Micrografia eletrônica da região nuclear. Note que a cromatina aderida fica excluída da área sob o complexo do poro. Foto: Larry Gerace.
Figura 3.5 : Esquema ilustrativo da organização geral do núcleo. Esse esquema se baseia em informações obtidas a partir de micrografias como a mostrada na Figura 3..
Envoltório nuclear
Complexo do poro
Lâmina nuclear
Cromatina aderida Matriz nuclear 1 Pm
Retículo endoplasmático
Filamentos intermediários
Poro
Cromatina
Núcleo
Centrossomo
Lâmina nuclear
Membrana externa 1 Pm (^) Membrana interna Envoltório nuclear
Microtúbulos
{
AULA
3
MÓDULO 1
Figura 3.8 : Procedimento para dissociar os nucleossomos e separá-los das histonas.
Existe ainda uma outra histona compactando o DNA, a histona H1, que não faz parte do nucleossomo. Ela fica por fora do arranjo de nucleossomos, ajudando a torná-lo mais frouxo ou mais compactado ( Figura 3.9 ).
DNA de ligação
Histonas do nucleossomo
DNA de ligação digerido por nucleares
200 pares de nucleotídeos
Dissociação com alta concentração de sal
Nucleossomos liberados
Octâmero de histomas
Dissociação
Duplas hélice de DNA
H 2 A H 2 B H^3 H 4
11nm
Figura 3.7 : Modelo do nucleos-^ a^ b somo visto de dois ângulos diferentes, com o DNA represen- tado pelo tubo helicoidal (A) ou pelas linhas paralelas (B) ao redor das histonas. Esquema de Evangelos Moudrianakis.
BIOLOGIA CELULAR II | Núcleo interfásico
Figura 3.10 : Micrografia eletrônica de um trecho de DNA no estado compactado natural (a) e depois descompactado (b). A molécula nativa (A) é chamada fibra de 30nm e o DNA no seu estado descompactado (B) é comparado a um colar de contas. Fotos: (a) de Barbara Hamkalo; (b) Victoria Foe.
Figura 3.11 : Nucleossomos enrolados sobre si próprios. Em (a), aspecto frontal de uma fibra, em (b), o aspecto lateral. Os primeiros seis nucleossomos estão numerados, para facilitar a compreensão.
A estrutura formada pelos nucleossomos intercalados por DNA de ligação assemelha-se a um colar de contas ( Figura 3.10.b ). Quando assume a configuração nativa, ela se arranja como a fibra de 30nm ( Figura 3.10.a ). A fibra de 30nm consiste no colar de nucleossomos enrolado sobre si próprio, encurtando muito a molécula de DNA ( Figura 3.11 ).
Figura 3.9 : A histona H1 liga-se por fora dos nucleossomos, ajudando a mantê-los mais próximos ou mais afastados.
Histona H
Nucleossomo
Octâmero de histomas
30nm
1
2
(^3 )
5
6 1
2
3
4
(^5 )
a
b
a b
50nm
BIOLOGIA CELULAR II | Núcleo interfásico
Se você reparar nas micrografias eletrônicas de núcleos interfásicos, sejam irregulares como os da Figura 3.2 , ou regulares, como os das Figuras 3.5 e 3.14 , vai perceber que parte da cromatina tem aspecto bastante eletrondenso.
Figura 3.13 : Um cromossomo metafásico pode ir sendo desenrolado até chegar ao estado mais simples do DNA, a dupla hélice.
Cromossomo metafásico inteiro
Segmento de cromossomo na forma condensada
Segmento de cromossomo na forma estendida
Colar de contas de nucleossomos
Pequena região da dupla hélice de DNA
1400nm
700nm
300nm
30nm
11nm
2nm
Fibra de 30nm formada por nucleossomos compactados
AULA
3
MÓDULO 1
Figura 3.14 : Micrografi a eletrônica de um núcleo interfásico típico de células de mamífero. Foto: Daniel Friend.
O aspecto eletrondenso está, na maior parte das vezes, relacionado ao estado compactado da cromatina, a heterocromatina (nem sempre, você já vai conhecer as exceções). Poderíamos, assim, correlacionar as regiões eletrondensas do núcleo com regiões do genoma que não estão transcrevendo. Observando bem as fotos, você vai reparar que sempre existe heterocromatina na região mais periférica do núcleo, bem juntinho do envoltório nuclear. A identificação dessa parte da cromatina veio por acaso: testando o soro de pacientes de doenças auto-imunes por imunofluorescência em células em mitose e na intérfase, constatou-se que os anticorpos fluorescentes reconheciam os telômeros dos cromossomos mitóticos e também a heterocromatina aderida ao envoltório nuclear em células interfásicas. Examinando células de outras espécies, nem sempre encontramos os telômeros aderidos ao envoltório nuclear durante a intérfase. Mas esse dado foi importante porque deu início à idéia, cada vez mais aceita, de que durante a intérfase os cromossomos descondensados não estão embolados dentro do núcleo. Muito ao contrário, parece haver grande organização do genoma. Alguns autores supõem que, à semelhança do que ocorre no citoplasma, existe um nucleoesqueleto sustentando essa organização. De fato, se aplicarmos a núcleos isolados a mesma metodologia de extração do citoesqueleto, usando detergentes não-iônicos, um material protéico é isolado. Sob certas condições, esse material parece formar filamentos, mas sua organização não é semelhante à dos grupos de filamentos do citoesqueleto. Nenhuma das proteínas conhecidas do citoesqueleto faz
DOENÇAS
AUTO - MUNES
São causadas por falhas do sistema imune, que não elimina células produtoras de anticorpos que reconhecem as moléculas do próprio indivíduo ( self ). Os pacientes dessas doenças possuem na circulação anticorpos que conhecem e atacam suas próprias células.
5 Pm
AULA
3
Durante a intérfase, as regiões dos cromossomos que contêm genes MÓDULO 1^ que codificam para os RNA se aproximam. Os genes que codificam os mRNA que vão produzir as proteínas ribossomais no citoplasma também se aproximam no nucléolo. Depois de serem produzidas no citoplasma (por outros ribossomos!), as proteínas ribossomais voltam ao núcleo pra começar a se associar aos rRNA e formar novos ribossomos. Mas atenção, perigo! Se os ribossomos ficassem prontos, poderiam iniciar a tradução dos mRNA antes que eles fossem processados! Lá se ia a maior vantagem evolutiva dos eucariotos por água abaixo! As subunidades ribossomais ficam quase prontas no núcleo, mas só amadurecem depois de chegar ao citoplasma ( Figura 3.16 ).
Gene de rRNA DNA Transcrição rRNA precursor
Nucléolo
Subunidade maior imatura Subunidade menor
Proteínas ribossomais sintetizadas no cito- plasma
Transporte e maturação ativa dos ribossomas
RNA e proteínas envolvidas no processamento
Citoplasma
18S rRNA
40S
5.8S 2.8S 5S
60S
Nucléo
rRNA
Figura 3.16 : Esquema geral da produção de ribossomos pelo nucléolo.
BIOLOGIA CELULAR II | Núcleo interfásico
Figura 3.17 : Sub-regiões do nucléolo. Fotos: E. G Jordan e J. McGovern.
Heterocromatina periférica Envoltório nuclear Nucléolo
Área densa Área granular Centro fibrilar 2 Pm 1 Pm
Todo o processamento dos rRNA a partir do precursor é realizado no nucléolo por um conjunto de RNA e proteínas muito bem organizado. As proteínas vêm do citoplasma e se juntam ao nucléolo, facilitando o arranjo ao mesmo tempo em que os RNA são processados. Para que tudo funcione a contento, o próprio nucléolo precisa ser muito organizado. Em micrografias de grande aumento é possível perceber que o nucléolo é subcompartimentalizado, com regiões densas e regiões fibrilares ( Figura 3.17). Com tantos RNA e proteínas associados, não é de estranhar que o nucléolo seja eletrondenso, apesar de não ser formado por heterocromatina!
QUANTOS NUCLÉOLOS TEM UMA CÉLULA?
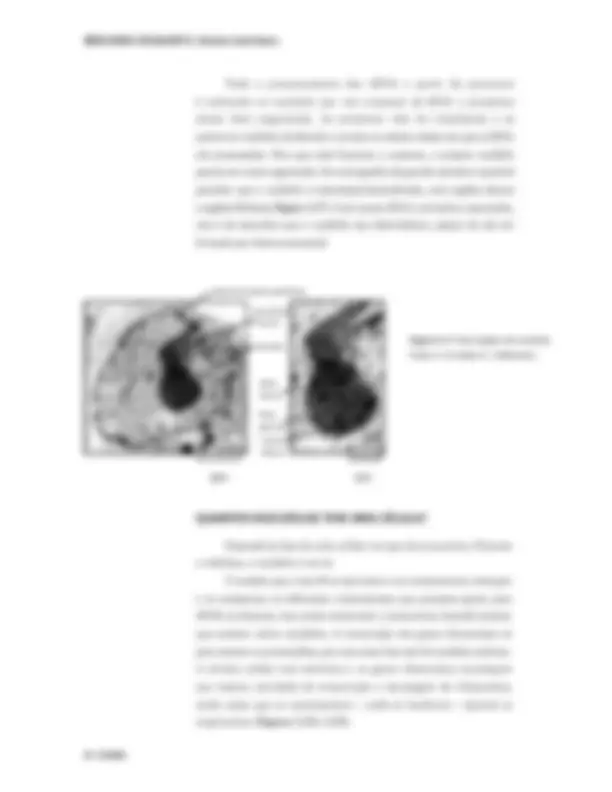
Depende da fase do ciclo celular em que ela se encontra. Durante a intérfase, o nucléolo é um só. À medida que a fase M se aproxima e os cromossomos começam a se compactar, os diferentes cromossomos que possuem genes para rRNA se afastam, mas ainda continuam a transcrever, fazendo parecer que existem vários nucléolos. A transcrição dos genes ribossomais só pára mesmo na prometáfase, por isso nessa fase não há nucléolo nenhum. A divisão celular mal terminou e os genes ribossomais recomeçam sua intensa atividade de transcrição e montagem de ribossomos, ainda antes que os cromossomos – onde se localizam – possam se reaproximar ( Figuras 3.18 e 3.19 ).
BIOLOGIA CELULAR II | Núcleo interfásico
Figura 3.20 : Imunofluorescência usando anticorpos que reconhe- cem diferentes subcompartimentos nucleares. As setas apontam os corpúsculos de Cajal. As regiões brancas correspondem a speckles. Os grandes círculos escuros correspondem ao nucléolo (essa célula está bem no início de G1). O cinza do fundo corresponde à cromatina. Foto: Judith Sleeman.
CONCLUSÃO
O ambiente nuclear é, portanto, altamente organizado, com compartimentos formados pela associação de macromoléculas que se mantêm unidas sem estarem cercadas por uma membrana. Apenas o envoltório nuclear é formado por duas bicamadas lipídicas, como veremos na próxima aula.
R E S U M O
- Nos eucariotos, o genoma fica dentro de um compartimento específico, o núcleo.
- A maioria das células eucarióticas possui apenas um núcleo, mas existem células binucleadas, como alguns protozoários; multinucleadas, como as fibras musculares e células anucleadas, como os eritrócitos de mamíferos.
- A presença do núcleo faz com que os eventos de transcrição e tradução ocorram em compartimento separados, possibilitando o processamento do RNA.
- Nucleossomos são arranjos octaméricos de histonas em torno dos quais se enrola a dupla fita de DNA com cerca de 200 pares de bases. O DNA entre os nucleossomos é chamado DNA de ligação.
- A cromatina corresponde ao DNA compactado por proteínas e pode estar alternadamente na forma de eucromatina, em processo de transcrição, ou de heterocromatina, estado mais compactado em que não há transcrição.
- O nucléolo corresponde à região em que estão sendo sintetizados os ribossomos, por isso apresenta-se mais denso que o resto da eucromatina.
AULA
3
MÓDULO 1
EXERCÍCIOS
- Quais as vantagens evolutivas do aparecimento do núcleo?
- Construa um esquema com a organização geral do núcleo interfásico.
- Qual a importância da compactação do DNA?
- O que é um nucleossomo?
- O que é DNA de ligação?
- Qual o papel da histona H1?
- Diferencie eucromatina de heterocromatina.
- O que vem a ser a heterocromatina aderida ao envoltório nuclear de células interfásicas?
- O nucléolo é heterocromatina? Por quê?
- Podemos dizer que os genes que codificam os ribossomos ficam todos em um mesmo cromossomo?