Pré-visualização parcial do texto
Baixe Capitulo 03CEM BILHOES e outras Notas de estudo em PDF para Bioquímica, somente na Docsity!
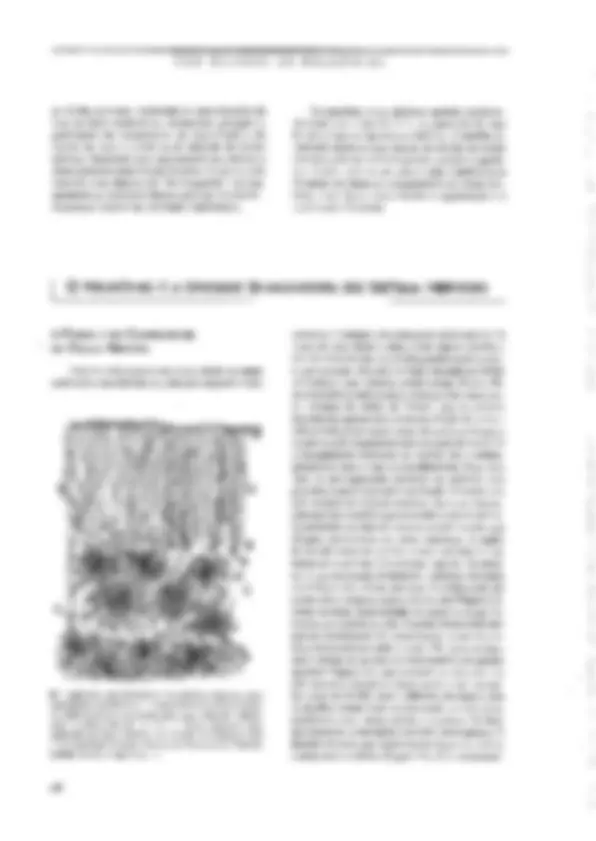
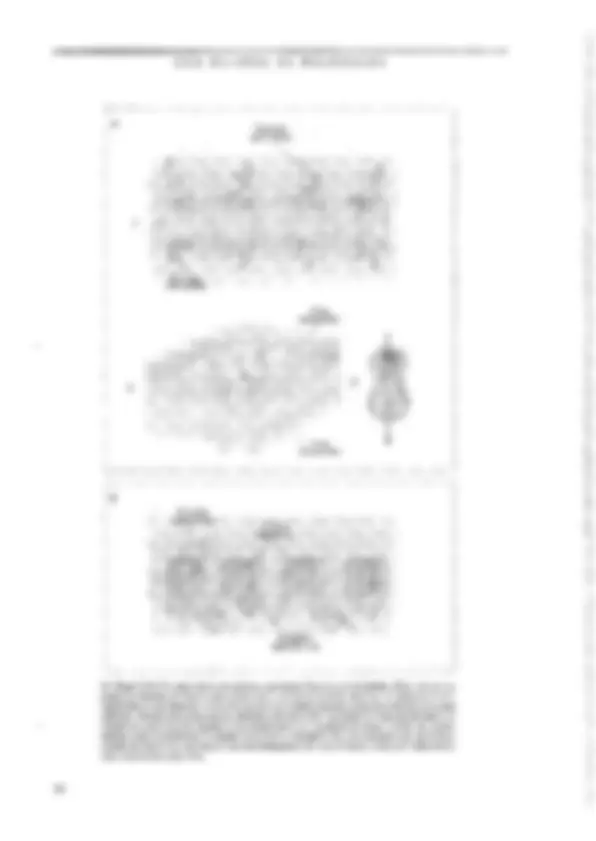
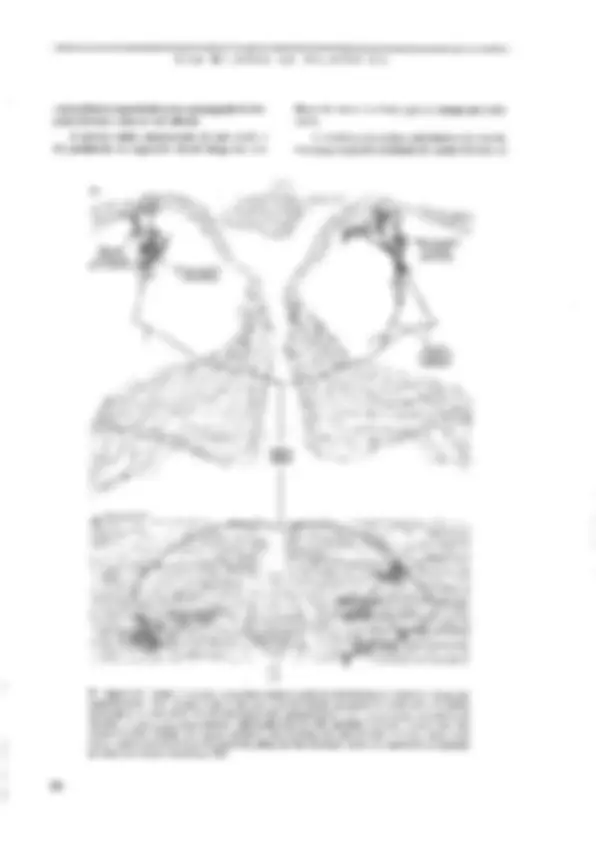
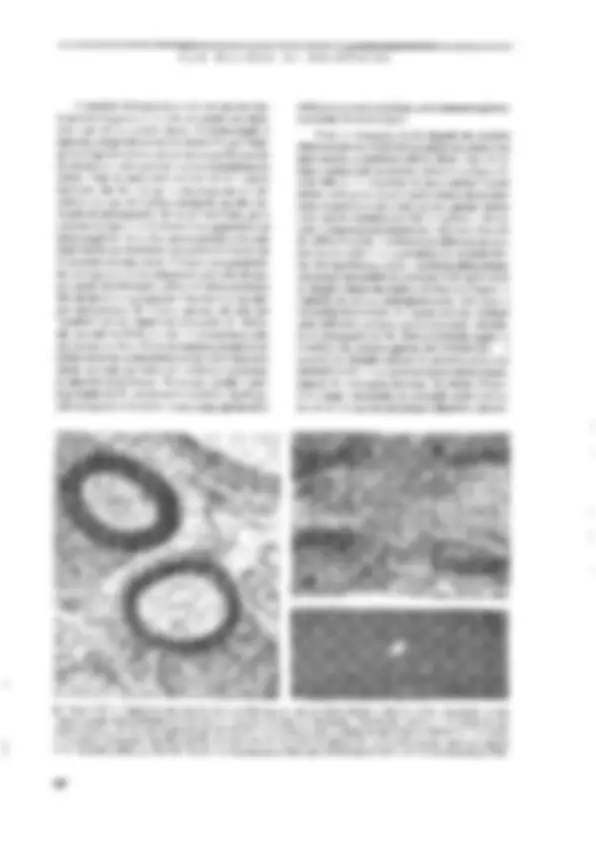
SABER O PRINCIPAL Resumo O neurônio é a unidade sinatizadoia do-sistgi gélula especializada, com. vários piolonganiêntos pira a E nais e um finico para a emissão desinais. Sua estrátga intêma é s lhants à das demais células animáis; córh algimas: agmtaridadês -D) de sua natureza sjnalizadora. É dentro e para fora do neurônio. Nurna situação hipotêtica-de “repotigh funciônal”, eneurônia ap; senta um estado constante chamado potencial de repousáida céluta ngr- vosa. Como gm todas as células o“interigrdo agurômb:d nem relação ao exterior, uma diferexça de potencialimantida constaniê: continuo fluxo de íons através ia membrana. É O sinal elétrico que-o neurênio utiliza como gimidadé de infor: É o impulso nervoso ct potencial de ação. Esté é um episódio rápido de inversão da polaridada da membrana, produzido seletiva e consecutiva de cai de Na* & K*, causamio-um gaudal fluxo iônico através da inembrana que provoca a inversão dejgua pol dade elétrica. Q) potencial de ação é propagável ag. longo. do axônio; portanto, conduzido de'uma exijzimidadg à outra dá neuzônio. O sistema nervoso não é constituído fipenas de peurônios, nas bém de uma outra família de células chamadas coletivamente-de ts absorção de substências do inejy externo vizinho agncurônio e suátigirs= formação em substâncias (fteig:a ele, o-isolamento da meiizbrana axônios, a defesa imunitária de'sistema nervoso g muitas outras, CEM BILHÕES DE NEURÔNIOS as células nervosas, controlam as concentrações de ions no meio extracelular, armazenam glicogênio, participam dos mecanismos de cicatrização e de defesa em caso de lesão ou de infecção do tecido nervoso, fornecem uma capa isolante aos axômios e desempenham outras tantas funções. Como se pode concluir, suas funções são “de retaguarda”, ou seja, garantem as condições básicas para que os ncurôni- os passam exercer sua atividade sinalizadora, | O NEURÔNIO É A UNIDADE SINALIZADORA DO SISTEMA NERVOSO Os neurônios e os gliúcitos operam cocrdena- damente (ver o Quadro 3,2). A separação de suas funções é apenas um recurso didático. A estreita co- operação entre as duas classes de células do tecido nervoso pode ser avaliada quando o tecido é agredi- do e lesado, pois nesses casos a glia contribui para bloquear ou diminuir a degeneração da célula ner- vosa, e em alguns casos facilita a regeneração e a restauração funcional, A FORMA E OS COMPONENTES DA CÉLULA NErvOSA Todo neurônio possui um corpo eclular ou suma, onde estão concentradas as principais organelas intra- B> Figura 3.2, Cajal desenhou também gliócitos. Neste caso, estão representados os astrócitos (C) é os oligodendrácitos (D) do cerebeto. As células B são neurônios (conhecidos como células de Purkinje), com um uxônio marcado pela lera à, Outros elementos estão assinalados por letras diferentes. Reproduzido dc 5, Ramón y Cajal (1955) Hlisiologie du Systême Nervetas de | Homune & des Vertébrás, Instituto Ramon y Cajal, Espenhu. 68 celulares. O diâmetro do soma pode variar mais de 10 vezes de uma célula a outra, desde alguns micrôme- tros até cerca de cem, As células granulares do cerebe- lo, por exemplo, têm cerca de 6pum, enquanto as células de Purkinje, suas vizinhas, podem atingir 80 mm, Es- sas dimensões conferem aos neurônios, individuaknen- te, volumes da ordem de 105m?, que no entanto representam apenas uma mimúscula tração do volume celular total, já que quase todos cles possuem longos e numerosos prolongamentos que emergem do soma. Os prolongamentos neuronais ou neuritos são o axônio, geralmente único, e um ou mais dendritos. dois tipos de prolongamento conferem ao neurônio uma polaridade que é essencial à sua função. O axônio vei- cula os sinais de saída do neurônio, isto é, as informa- ções que esse neurônio gera e conduz a outras células. Os dendritos, por sua vez, recebem as informações que chegam, provenientes de outros neurônios. A região de contato entre um axônio e outro neurônio é a si- napsc, tema principal do próximo capítulo. Os neuri- tos é que expressam fortemente a extrema variedade morfológica das células nervosas. O axônio pode ser muito curto e simples, como o de um cone (Figura 3.3), célula da retina especializada em captar a energia lu- minosa que penetra no olho. O axônio dessa célula tem poucos micrômetros de comprimento, e termina em uma intumescência muito simples. No outro extremo está o axônio de um Lipo de motoneurônio da medula espinhal (Figura 3.3), que comanda os músculos dos pés: seu comprimento no homem pode atingir um me- to (cerca de 20.000 vezes o diâmetro do soma), e ele se ramífica muitas vezes comunicando-se com outros neurônios e com várias células musculares. No lado dos dendritos, a varicdade é também muita grande. O dendrito do cone, que mencionamos há pouco, é único e curto como o axônio (Figura 3.3). Já os motoneurô- As UNIDADES DO SISTEMA NERVOSO nios medulares possuem vários dendritos emergindo do soma. Cada um deles se ramifica várias vezes nas proximidades do corpo celular (Figura 3.3), formando O que se conhece como árvore ou campo dendrítico, quena verdade compõe a região “receptiva” do neuró- nio, ou seja, aquela que recebe as informações dos ontros neurônios. A Figura 1.8€C apresenta um exem- plo extremo de complexidade dendrítica, o da célula de Pukinje do cerebelo, cuja árvore é tão ramificada que permite o contato de 200.000 fibras aferentes! Essas numerosas varicdades morfológicas das células nervosas não são casuais, mas, sim, representam adap- tações da forma neuronal às diferentes fanções excrci- das pelos neurônios. de wês camadas (lâminas), sendo a central elétron-lú- cida (transparente aos elétrons). e as duas de fora elé- tron-densas (vpacas ao feixe de elétrons). Essa constituição laminar reflete a estrutura molecular da membrana, que é formada por uma dupla paliçada de lípídiosS, dentro da qual flutuam proteinasS de dife- rentes tipos e funções (Figura 3.4B). Não é só a borda externa da célula nervosa que é envolvida por mera- brana. Como tudas as células, o neurônio também pos- sui um complexo sistema de cisternas envolvidas por membranas de constituição e estrutura minto semelhan- te à da membrana plasmática. Essas cisternas têm como função gerenciar o tráfego interno das moléculas sin- tetizadas sob comando genético. Além disso, o núcleo te Figura 1.3. Através dos desenhos de Cajal podermos comparar os cones da retina de um peixe (4), assinalados pela Ictra a à esquerda (em vermelho), com os moluneurônios da medula espinhal de um pinto (B), indicados pelas letras à, à direita, também em vermelho, Outros elementos estão representados por letras diferentes (em preto). No desenho, o denárito do cone aponta para cima, enquanto o axônio uponia para baixo. Os dendritos dos motoneurônios emergem em todas as direções, « o axênio está assinalado pela letra b. A modificada de S. Ramón y Cajal (1972) The Struchue af the Retina. Charles C. Thomas, EUA. B modificado de S. Ramin y Cajal (1935) Ilistologie du Systême Nervewx de iºHomme & des Vertébrés. Instituto Ramón y Cajal, Espanha. O neurônio, com seus prolongamentos, é uma uni- dade completamente envolvida por uma membrana que separa o compartimento intracelular do extracelular (ver no Quadro 3.1 como se chegou a essa certeza). Como veremos, essa separação não isola totalmente o interior do neurônio do meio extracelular. Muito pelo contrário, sua permeabilidade seletiva permite inten- sas trocas ds íons entre esses dois compartimentos, e é exatamente essa propriedade que torna possível a ge- tação e a condução de sinais bioelétricos. A membra- na plasmática aparece ao microscópio eletrônico como uma fita continua que envolve a célula sem interrup- ções (Figura 3.44). Examinando-a em grande aumen- to, os morfologistas verificaram que ela é constituída e muitas organelas celulares são também delimitados por membrana com as mesmas características da mem- brana plasmática, O citoplasma e o núcleo compõem todo o interior da célula nervosa (Figura 3.44). Não se deve imagi- nar que o citoplasma seja constituído exclusivamente de um líguido no qual flutuariara as organelas intrace- lulares. Sua constituição é bem mais complexa. Na verdade, o citoplasma é composto por um meio líqui- do denso (quasc uma gelatina, chamada citosol) « por proteinas organizadas na forma de fibrilas, que com- púem o citoesqueleto. É o citoesqueleto que mantém a forma peculiar de cada neurônio. É ele também que [24 AS UNIDADES DO SISTEMA NERVOSO transcrição, isto é, a síntese de moléculas de RNA mensageiro (mRNA), que formam réplicas perfeitas de segmentos do DNA destinadas à síntese de protei- nas. E é justamente através dos poros do envelope nu- clear que 0s mRNAs passam para o citoplasma, onde se relinem a pequenos grânulos chamados ribossomos (Figura 3.68). Os ribossomos vão se associar em se- giiência ao mRNA, formando os polissomos, e vários destes se ligarão à superfície externa do retículo endo- plasmático, dando-lhe um aspecto rugoso caracteristi- co (Figura 3.64). Nos polissomos ocorre a tradução (Figura 3.6), ou seja, a sintese de proteínas do neu- rônio. O retículo endoplasmático Tugoso é muito pro- nunciado nos neurônios devido à intensa atividade de sintese protéica dessas células, e pode ser identificado em preparações histológicas coradas como grânulos de aspecto “tigróide” (assim os descreveram os histolo- gistas do século passado!) cujo conjunto ficou conhe- cido como substância de Nissl (Figura 3.7). Algumas proteínas recém-sintetizadas vão difum- dir de volta ao núcleo, outras ficam no citosol para serem incorporadas às organelas citoplasmáticas, e um terceiro grupo é armazenado no interior do retículo para ser posteriormente transportado aos prolungamentos do neurônio e até secretado para o meio extracelular. Do retículo endoplasmático brotam pequenas vesicu- 5» Kigura 3.5. Os elementos do citoesqueleto podem ser identificados ao microscópio eletrônico dentro de um axônio cortado transversal (A) ou longitadinalinento (RB). A organização molecular dos elementos da citoesqueleto foi decifrada por técnicas bioquímicas (C-E). Os microtibulos são formados por 13 protofilamentos de tubulina formando um cilindro (CJ. Os neurotitamentos são constituídos par muitas unidades fibrilures mais ias trançadas entre si (D), & os microfilamentos compõem-se de duas segilências helicoidiuis de moléculas globnlares de actina (E). Fotos Ae B cedidas por Ana M. D. Martincz, do Depurtumento ds Histologia e Embriologia do Instituto de Ciências Biomédicas da UFRJ. 71 CEM BILHÕES DE NEURÔNIOS 72 Cem BILHÕES DE NEURÔNIOS Cisternos roreino. Mitocbngria => Figura 3.6, A membrana nuclear é continua com o sistema de cisternas do neurônio (sotas vermclhas em A). As cisternas participam ativamente da sintese de proteinas. Os RNA mensageiros (mRIA) são sintetizados no núcico a partir do DNA (B), c exportados pelo citoplasma, onde se associam aos ribossomos para formar os polissomos (C). É nestas estruturas que ocorre a síntose das proteinas citoplasmáticas, mitocondriais & vesientares. Foto em A cedida por Ana M.B. Martinez, do Departamento de Histologiu e Embriologia do Instituto de Ciências Biomédicas da UFRJ. Praticamente todos os componentes do soma neu- ronal estão presentes nos dendritos, especialmente na- queles mais calibrosos que emergem do soma. Nos ramos mais finos diminui ou desaparece a substância de Nissi, assim como o aparelho de Golgi e os micro- túbulos do citoesqueleto. O axônio emerge do soma através de uma re- gião funcionalmente especializada chamada cone de implantação, segmento inicial ou zona de disparo. Como veremos adiante, essa região é muito excitável, 2 é nela que aparece o impulso nervoso que será poste- riormente conduzido ao longo do axônio na direção de sua extremidade. O axônio é uma parte do neurônio de grande importância pelo papel que exerce como condutor dos impulsos nervosos. Historicamente, axô- nios de animais inverichrados (como as lutas) foram Substância: dg Nisst E» Figura 57, À chamada substância de Nissl recebeu esse nome em homenagem ao psiquiatra alemão Franz. Nissl (1859-1949), o primeiro à revelá-la utilizando anílinas. O método de Niss! foi aplicado pelos primeiras histelogistas e uindi rotineiramente utilizado. A cétuta cm A é um motoneurônio da medula do coelho, desenhado por Cajal. Um corpúsculo de Nissl é assinulado pela letra b, a fetra e denota estruturas mais finas do citoplasma, d assinala o mícico, ee a mostram dendeitos. A foto em B mostra neurônios piramidais do córiox de um hamster, corados pelo método de Nissl e fotografados vo microscópio pico. A reproduzido de S. R. Cajal (1955) Histologie du Systême Nervezs de Homme & des Vertébrês. Instituto Ramón y Cujal, Espanha. 74 AS UNIDADES DO SISTEMA NERVOSO do Figura 3.8. As espinhas dendríticas são diminutas prorrusões que emcigem do um tronco dendrílico principal (A). As fotos 4, B e C são sucessivas ampliações (retângulos brancos) de uma célula preenchida cum um corante Auoresceate, vista ao micrascópio óptico. Os círculos brancos em B assinalam cspifihas dendriticas. A foto C mostra ima delas em grande ampliação. À foto D mostra um corte ultratino visto «o microscópio eletrômica, ilustrândo uma sinapée (seta) de um terminal axônico (à direita) com uma espinha (à esquerda). A foto E representa o molie metálico de um dendrito visto ao microscópio eletrônico de varredura, apresentando uma espinha dendrífica integta e inúmeros “talós quebrados” de espinhas (setas vermelhas). D modificado de A. Peters et ul. (1976) The Fine Structure of the Nervous System. WB. Saunders Co., EUA. E modificado de K.M. Harris ct ul. (1992) Journal af Neuroscience 12: 265 -2705. Fatos 4, B e C cedidas por Monica Rocha, do Departamento de Farmacologia do Instituto de Ciências Biomédicas da UFRJ. os modelos experimentais que possibilitaram a com- preensão dos mecanismos moleculares geradores dos sinais elétricos neuronais. A membrana do axônio re- eebe o nome particular de axolema, embora sua estru- tura e sua função não sejam muito diferentes daquelas dos dendritos. O mesmo se pode dizer do citoplasma, que no axônio recebe o nome particular de axoplasma. No axoplasma — como no citoplasma dos dendritos mais finos — não existe o retículo endoplasmático ru- goso que constitui a substância de Nissl. Por isso, as técnicas histológicas comuns (chamadas técnicas de Nissl) revelam apenas os corpos celulares e os ramos dendríticos mais calibrosos (Figura 3.7), mas não os axômias. Num curte histológico corado pela técnica de Nissl, os feixes de fibras apresentam-se com uma ima- gem-“negativa”, isto é, sem cor, mostrando-se apenas por contraste corh.as repiões. que contêm os corpos cu- lulares. Oaxoplasma, entretanto, contém mitocôndrias esparsas, vesículas ém trânsito e microtúbulos, neurofi- lamentos e microfilamentos. Os microtúbulos são es- senciais à concretização de uma importante função de comiumicação entre o soma e as extrentidades do axô- nio: o fluxo axoplasmático. O fluxo axoplasmático é um movimento contínuo de moléculas ou de organelas membranosas que utili- zam os microtúbulos como trilhos (Pigura 3.9). O flu- xo pode partir do soma em direção às extremidades do axônio, caso em que se denomina fluxo antcrógrado, 75 AS UNIDADES DO SISTEMA NERVOSO Muitos axônios, tanto no sistema nervoso central como no sistema nervoso periférico, são envolvidos por uma cobertura isolante composta por lipídios e proteínas, chamada bainha de mielina (Pigura 3.5). No SNC, a bainha de mielina é produzida pelos oligoden- drócitos, e no SNP pelas células de SchwannS, A dife- rença entre o SNC e o SNP quanto à bainha de mielina não se resume ao gliócito que a produz, mas se esten- de à sua composição molecular. Recentemente, desco- briu-se que há proteinas na mielina central que bloqueiam o crescimento regenerativo de axônios le- sados. Essas proteínas não existem na miclina perifé- rica, € é isso que explica por que os axônios periféricos são capazes de regeneração, enquanto os axônios cen- trais não são, Nasua extremidade distal, o axônio pode se ramifi- car profusamente, formando uma arborização terminal chamada telodendro. Cada ramo de telodendro, por sua vez, forma múltiplos botões sinápticos que se encon- tram apostos aos dendritos cu ao soma de outros neurô- nios, formando as sinapses, Essas estruturas de comunicação entre neurônios são o tema do Capítulo 4. A MEMBRANA E OS SINAIS ELÉTRICOS DO SisTEMA NERVOSO Talvez seja um exagero dizer que a membrana é o constiluinto mais importante do neurônio, já que muitos outros constituintes são essenciais à vida e ao funcionamento da célula nervosa. Mas é certo, pelo menos, que a membrana do neurônio apresenta uma propriedade muito particular que o distingue da mai- oria das células do organismo. Essa propriedade — exoitabilidade -- permite que o neurônio produza, conduza e transmita a outros neurônios os sinais elé- tricos em código que constituem a linguagem do sis- tema nervoso. A descoberta dessa propriedade dos neurônios — a capacidade de gerar sinais elétricos — foi feita ainda no século XIX pelo fisiologista alemão Emil DuBois- Reymond (1818-1896), antes mesma de se conhecer a existência dos neurônios. Os mecanismos biofísicos subjacentes, entretanto, só foram revelados 100 anos depois. Desde o início se compreendeu que a bioele- trogênese não deveria depender do fluxo de elétrons, como nos circuitos elétricos comuns, mas de íons, as moléculas eletricamente carregadas tão conuns den- tro e fora das célutas. Cedo se compreendeu também que as correntes iônicas passavam através da membra- va neuronal. No entanto, como explicar que fosse as- sim? Os íons biológicos são fortemente hidrofilicos, e dificilmente conseguiriam atravessar um meio lipídi- co como é a membrana. Rra preciso imaginar a pre- sença de poros eu canais na membrans. que permitis- sem a passagem dos fons e, assim, possibilitassem a geração de sinais elétricos. Mas nem todos os íons atravessam a membrana neuronal. Na verdade, pou- cos deles conseguem passar. Logo, os canais iônicos deveriam atuar como filtros de permeabilidade sele- tiva, permitindo que alguns ions passassem de fora para dentro e de dentro para fora da célula, enquanto outros fossem bloqueados em ur dos dois comparti- mentos, O conceito de canais iônicos evoluiu até os tempos atuais, e está no centro dos mecanismos mole- culares que possibilitam a capacidade sinalizadora dos neurônios. CANAIS lÔNICOS Canais iônicos são proteínas integrais de membra- na, isto é, proteinas incrustadas na bicamada lipídica, que têm a capacidade de deixar passar íons de modo seletivo, continuamente ou em resposta a estímulos elétricos, químicos ou mecânicos. Os canais que dei- xam passar os ions continuamente são chamados ca- nais abertos, e os que só abrem cm resposta a estímulos especificos são chamados canais controlados por comportas. Tanto uns como outros podem ser alta- mente especificos: há canais para cátionsS, como o sódio (Na?), o potássio (K*) e o cálcio (Ca"), e canais pata ânionsS, como o cloreto (CH). Os canais contro- lados por comportas podem ser abertos por alterações da voltagem que existe naruralmente na membrana enire o interior c o exterior da célula nervosa, Neste caso, são considerados dependentes de voltagem. Outros podem ser abertos por substâncias específicas (ligantes) como neurotransmissores, neuropeptideos e hormônios, e neste caso são considerados dependen- des de ligantes. Finalmente, outros canais são abertos por certos tipos de energia mecânica e radiante que incidem diretamente sobre a membrana. Ao pensar em canais incrustados na membrana neuronal, capazes de deixar passar íons de modo sele- tivo e assim gerar sinais bioclétricos, algumas pergun- tas fundamentais se colocam. Qual a estrutura molecular desses canais iônicos? Como se abrem é como se fecham? Como passam os íons? Como são selecionados? Comecemos pela estrutura molecular. Tem sido possível conhecer detalhos da estrutura molecular dos canais dependentes de voltagem, e mais recentemente de um dos canais dependentes de ligantes, precisamente aquele que é aberto pelo newrotransmissor acetilcoli- na, Todos eles são glicoproteínas, geralmente forma- das por subunidades repetidas ou diferentes (Figura 3.10). O canal de Na” dependente de voltagem, por 77 78 CEM BILHÕES DE NEURÔNIOS Dominio extracelular intracelulor Face » Foce intracelular Domínio entracelular Domínios intracelulgres às Figura 3.10. Os canais iônicos são proteinas, geralmente formadas por subunidades. Estas, por sua vez, podem ser tormadas por motivos moleculares, isto 6, segilências protéicas repetitivas. A representa as cinco subunidades do canal dependente de acerilcolina (cm bege), também chamado receptor de acetileolina ou receptor colinérgico. B representa apenas uma das subunidades do canal do Na+ dependente de voltagem (subunidade 2), formada por quatro motivos repetitivos (em laranja) unidos por sequências da mesma protein que formam domínios intra e extracelulares. No desenho em AL estão es subumidedes do canal colinérigico (em lons de bege), reunidus em círculo (A2) para form u estrutura tridimensional do canal. A3 mostra o icone dos canais iômicos, como representamos neste livro. CEM BILHÕES DE NEURÔNIOS of ii efe de Ê Ojo igagio ne õ pm sado Fe hidr Fosfolipídio Membrana." ços Er Segsos iênico: Plismática - “sai iônico o SBEbS 2 Gagos? “Sogvo aê “4, Be Figura 3.11. Os dois compartimentos separados pela membrana plasmética (desenho de baixo) contêm fons bidratados (envoltos por uma nuvem de moléculas de 4gué). Nessa condição (A), os ions interagem com a parte externa da membrana (região bidrofiiva), mas não conseguem ultrapassar a parte interna (região hidrofábica). Os canais iônicos fornecem uma via de passagem seletiva para os. íons. Alguns canais como o de Na+ (B) possuem ura “fikvo molecular” no seu interior, na verdade um sítio de ligação para o Nat que permite à sun passagem para 0 outro lado da membrana. Modificado de S.A. Siegslbaum ]. Koester (2000), em Principies af Neural Seienee, 4a. Ed. (E.R. Kande! 2 colaboradores, Otgs.). MeGraw-IHL, Nova York, EUA. químico sobre a “upasição” do gradiente elétrico. O correntes iônicas e controlasse precisamente o fluxo atra» mesmo ocorre para os fons CF, que atravessam a mem- vés da seleção dos íons que passam pelos canais. brana de fora para dentro do neurônio. Tudo se passa Os canais abertos têm funcionamento mais sim- como se a membrana normalmente “represasse” as ples que os canais controlados por comportas. Estes so AS UNIDADES DO SISTEMA NERVOSO Meio extracelular (Not > K) Meio intracelular (> Na”) ie» Figura 3.12, À. O meio extracelular tem maior concentração de Na+ e C1., enquanto o meio intracelular tem maior concentração de Ke ânions inorgânicos (A-). B. Quando os cunuis de ua jon se abrem, por exemplo os de Ks, estes fons se deslocam movidos pelo seu gradiente quimico, e as suas concentrações exterma e interna tendem a se igualar. Isso não ocorre, entrctanto, porque eles se aglomeram na borda externa da membrana formando uma fina camade positiva (C), o que cria um gradiento que tende a freur u suída de K+, estabilizando a situação. últimos apresentam um problema para os biofísicos. Como explicar a abertura e o fechamento das compor- tas? A expressão “controle por comportas” não des- creve exatamente o que se passa. Tem um sentido figurado, de uma “represa”, cujas comportas se abris- sem parcialmente para dar vazão ao fluxo iônico. Na verdade, os canais controlados por comportas apresen- tam uma propriedade típica das proteínas, chamada atosteria. Por meio dessa propricdade, as proteinas podem assumir conformações moleculares diferentes, modificando sua disposição espacial. Quando assumem uma certa conformação, os canais não permitem a pas- sagem dos íons através do poro central. São ditos en- tão fechados. Em certas condições, entretanto, as subunidades protéicas se modificam espacialmente, passando a permitir o fluxo iônico. Essa mudança de conformação molecular pode ser provocada por me- canismos diferentes, segundo o tipo de canal. Nos ca- nais dependentes de voltagem, uma alteração da diferença de potencial elétrico da membrana pode ser um estímulo disparador da mudança conformacional. Nos canais dependentes de ligantes, por outro lado, ocorre uma reação química não-covalente do ligante (um neurotransimissor, por exemplo) com o domínio extracelular do canal. E, nos canais “mecânicos”, é ne- cessário um estiramento da membrana para provocar a aberlura da comporta. O estudo experimental dos ca- nais indicou que o comportamento dinâmico deles apre- senta três estados funcionais distintos: (1) um estado de repouso, durante o qual o canal está fechado, mas pode ser ativado (aberto) a qualquer momento; (2) um estado ativo, durante o qual o canal está aberto, e por ele passa o fluxo iônico, e (3) um estado refratário, durante o qual está fechado e não pode ser ativado. No estado ativo, um só canal pode conduzir até 100 mi- thões de íons por segundo. Esse número sugere que. mesmo em pequeno número, os canais iônicos podem ser bastante eficientes na geração de sinais clétricos através da membrana neuronal. NEURÔNIOS EM SILÊNCIO: O POTENCIAL DE REPOUSO Suponhamos a situação pouco comum de um neu- rônio em “silêncio”, ou seja, inativo do ponto de vista da produção de sinais elétricos de informação. Um experimentador que inserisse uma minúscula agulha condutora (microeletródio) dentro da célula observa- ria uma diferença de potencial constante entre as duas faces da membrana. Essa diferença de potencial é co- nhecida como potencial de repouso, e reflete a sepa- ração de cargas elétricas entre a face externa & a face interna da membrana celular (Figura 3.13). Como vi- mos anteriormente, essa separação de cargas é manti- da pela natureza isolante da bicamada lipídica que constitui a membrana e pelo gradiente quimico entre o meio intra € o meio extracelular. Aparecem então duas 81 AS UNIDADES DO SISTEMA NERVOSO igual a -60mV. O potencial de repouso naturalmente resultaria da combinação dos movimentos desses íons, que, como vimos, dependem da concentração de cada um deles e de sua permeabilidade através da membra- na neuronal. Pode-se calcular matematicamente csse potencial utilizando uma segunda equação fisico-qni- mica parecida com a de Nernst. O resultado bate pre- cisamente com o valor medido de -60 a -70mV. Além disso, as diferenças de permeabilidade entre os íons indicam que há mais canais abertos de K* no neurônio que canais de Na* e de Ch, e é por isso que o potencial de repouso é mais próximo do potencial de equilíbrio do K* do que do Na* e do CH. Em outras células, a situação é diferente. É o caso dos astrócitos, por exem- plo. Nessas células o potencial de repouso medido é exatamente igual a -75mV, o que nos leva a supor que elas só têm canais abertos de K*, As medidas de per- meahilidade iônica confirmam essa suposição. Essa situação de equilíbrio que resulta no poten- cial de repouso na verdade ocorre em um espaço mui- to pequeno siluado nas proximidades da face externa e da face interna da membrana neuronal. O gradiente eletroguímico, entretanto, reflete as diferenças de con- centração iônica em todo o volume intracelular e em todo o volume extracelular. Isso significa que os flu- xos iônicos transmembrana não se interrompem quando a situação de equilíbrio elétrico é atingida. Esse equilíbrio é dinâmico, e os movimentos iôni- cus continuam indefinidamente, Mas se isso é ver- dade, logo mudariam as concentrações relativas dentro e fora do neurônio, e consequentemente mu- daria também o gradiente eletroquímico. Como se explica que isso não ocorra? Entra em cena um elemento importante: a bomba de Na*K* (Figura 3.13). A “bomba” é uma proteína integral da membrana furmada por duas subunidades diferentes: uma subunidade catalítica” que atravessa a membrana (subunidade «:), e uma subunidade gli- coprotéica reguladora (B). A subunidade 0: tem sítios intracelulares de ligação para Na* e para uma molé- cula de alta energia, o ATP!, e sítios extracelulares específicos para o K*. O ATP transfere fosfato para a subunidade ct, cm presença de Na” do lado de dentro e de K* do lado de fora. A energia dessa reação de fosforilação possibilita a exteriorização de três tons Nat, em troca de dois ions K* levados ao interior do neurônio. Devemos perecber que esses movimentos iônicos se dão contra o gradiente eletroquímico, e por isso precisam da energia química fornecida pelo ATP. Além disso, devemos considerar a importância 1 Teifosfato de adenosina.... da homba de Na-/K* como um mecanismo de reposi- ção automática das concentrações iônicas, mutáveis pelo fluxo passivo que ocorre constantemente através dos canais iônicos abertos. NEURÔNIOS EM ATIVIDADE: O PorENCIAL DE AÇÃO O que caracteriza o neurônio não é o potencial de repouso que, afinal, existe em todas as células vivas do reino animal e do reino vegetal. É o potencial de ação (PA), um sinal elétrico muito rápido e de nature- za digital como o que os computadores produzem. Como nos computadores, o PA confere ao neurônio a capacidade de transmitir informação, já que o número de sinais emitidos em cada momento pode ser variado proporcionalmente a estímulos vindos de fora, ou mes- mo a estímulos gerados dentro do neurônio, Cientes de que lidavam com a unidade de código empregada pelo sistema nervoso, os biofísicos mos- traram-se muito interessados, desde o início do século XX, em compreender a bioeletragênese do PA, ou seja, os mecanismos envolvidos na sua geração. Um primeiro problema a resolver era o de como registrar o fenômena elétrico sem conhecer ainda seus determinantes biofisicos. Lsso pôde ser conseguido fa- cilmente, utilizando o mesmo experimento realizado pata registrar o potencial de repouso. Empregou-se um microeletródio inserido através da membrana para o interior do neurônio, seja em seu soma ou no axônio. Na verdade, os primeiros pesquisadores envolvidos com essa questão utilizaram axônios de lula que, como em muitos invertebrados, apresentam grande calibre, sendo por isso chamados axônios gigantes. Se o mi- eroeletródio está posicionado em um axônio, e este estiver desconectado do soma e mantido artificial- mente em um meio líquido apropriado, é preciso es- timular a membrana com minúsculos choques elétricos para obter potenciais de ação. Entretanto, se o experimento é realizado no soma de um neurô- nio íntegro, mantido artificialmente em cultura ou então dentro do SNC do animal, muitas vezes não é preciso estimulá-lo eletricamente, pois aparecem PAs espontaneamente na membrana. Dois pesquisadores britânicos, Alan Hodgkin (1914-1998) e Androw Huxley (1917- ), destaca- ram-se muito nesses experimentos pioneiros e contri- buíram fortemente para o desvendamento da bigeletrogênese do impulso elétrico do neurônio, sen- do por isso contemplados com o prêmio Nobel de me- dicina e fisiologia em 1963. Nos experimentos de Hodgkin e Huxley (Figura 3.14), um axônio gigante era removido da lula e mantido dentro de uma cubeta 83 CEM BILHÕES DE NEURÔNIOS com líquido contendo íons e nutrientes. Um par de ele- tródios de estimulação era posicionado. na superfície da membrana em um ponto do axônio, e ontro par era posicionado a uma certa distância, para registro dos fenômenos elétricos. No par de eletródios de registro, um deles era um microeletródio de ponta muito fina, e pedia ser inserido dentro do 4xônio através da mem- brana, Desse modo era possível registrar a diferença de potencial transmembrana (entre o axoplasma e o meio externo). Hodgkin c Huxley observaram que, em condições de repouso, seus aparelhos de registro mostravam um potencial de repouso constante de -70mV (Figura 3.144). Quando aplicavam um estímulo elétrico à membrana, ocorria uma variação súbita e passageira desse valor. O potencial da membrana aproximava-se de zero rapidamente e o ultrapassava, tornando-se po- sitivo em aproximadamente 40-50mV (Figura 3.14B). Essa fase foi chamada despolarização, Em seguida, a despolarização parava e o potencial da membrana retomava rapidamente a um valor próximo ao de re- pouso (Figura 3.14€, D). Essa fase foi chamada repola- tização. Tudo se passava em menos de 1 milissegundo! A rapidez do fenômeno elétrico. tornava quase impossível analisar sua gênese iônica. Hodgkin e Hu- xley, então, lançaram mão de uma técnica inventada alguns anos antes, capaz de “tixar” o potencial da mem- brana do axônio gigante em algum ponto entre -70 e +40mV, utilizando uma aparelhagem eletrônica espe- cial, A técnica tem sido usada até hoje e fico conhe- cida como fixação de voltagem. Com o potencial da membrana fixo no mesmo valor, tornou-se possível medir as correntes iônicas que atravessam a membra- na, No entanto, os registros de. medida combinavam os movimentos de todos os ions envolvidos no fenô- meno, especialmente o Na' e o K:. Seria necessário bloquear um deles para poder observar apenas o ou- tro. Isso foi conseguido utilizando a tetrodotoxina (TTX), bloqueadora dos canais de Na*, e o terraetila- mônio (TEA), bloqueador dos canais de K*. Sob ação do TEA, e com o potencial da membrana fixado em OmvY, registrava-se uma corrente rápida de fora para dentro, atribuída aos íons Na* (setas azuis na Figura 3,148). Sob a ação da TTX, entretanto, registrava-se uma corrente um pouco mais lenta, com polaridade inversa, isto é, com sentido de movimento de dentro para fora (setas violetas na Figura 3.14C). Quando o registro era feito sem nenhum dos bloqueadores, a cur- va obtida indicava uma-composição das duas corren- tes iônicas, A certeza de que eram o Na* e o K* os íons envolvidos no PA veio da utilização de isótopos radi- oativos desses ions, que permitiu medir as concentra- vões de um lado e outro da membrana (através da 84 medida da radioatividade dos isótopos) antes e depois do experimento. Concluiu-se que a fase de despolarização do PA era causada poí uma súbita abertura dos canais de só- dio dependentes de voltagem, que permitia um cauda- loso movimento dos fons-Na* para dentro do axônio durante menos de 1 milissegundo, Os canais de Na”, então; tórnavam-se aos poucos inativos, o que fazia cessaí em alguns milissegundos a corrente de sódio. A fase de repolarização do PA, entretanto, é mais rá- pida que a diminuição do Íluxo de Na*. A explicação é que entra em cena o potássio, cujos canais depen- dentes de voltagem abrem-se mais lentamente que os do sódio. A sáída de K” restaura a polaridade da mem- brana'nos níveis de reponso, mas durante um certo tempo cla permanece inexcitável, incapaz de gerar outros PÁs, Essa fasc inexeitável chama-se período refratário, e deve-se ao fato de que, após se abrirem, os canais iônicos passam ao estado inativo ou refra- tário, e não ao estado de repouso. Mais tarde os ca- nais voltam ao estado de repouso, e a membrana do axônio se torna outra vez excitável. Além disso, a bom- ba de Na*/K* se encarrega de restaurar o gradiente ele troquímico original. Atualmente, consegue-se registrar a atividade elé- trica de neurônios de mamiferos. o que confirma os experimentos e conclusões de Hodgkin e Huxley utili- zando o axônio gigante da lula. Nos neurônios os po- tencíais de ação são produzidos na região de implantação do axônio no corpo celular, uma região conhecida por cone de implantação, segmento inicial ou zona de disparo. Essa é uma região especializada que contém uma alta densidade de canais iônicos de- pendentes de voltagem, 0 que Ihc confere maior exci- tabilidade. Como consegiiência, o segmento inicial tem o mais baixo limiar de excitabilidade do neurônio, isto é, produz PAs quando seu potencial atinge -S5mV, ou seja, é despolarizado de apenas 10mV. Isso contrasta com o corpo celular, cujo limiar é bem mais alto (cer- ca de -45mV). Essa particularidade do segmento inici- ai faz com que nele esteja a origem natura! dos PAs do neurônio e que estes sejam conduzidos não em dire- ção ao soma, mas no sentido dos terminais telodêndri- cos do axônio. À ProrAGAÇÃO DOS SINAIS Etérricos DOS AxÔNIOS Depois do segmento inicial, que tem uma forma cônica, o axônio assume uma forma cilíndrica e se es- tende como um tubo sinuoso através do tecido nervo- so até atingir as suas células-alvo (Figura 3.15). Ao longo do caminho, esse tubo podes se ramificar várias