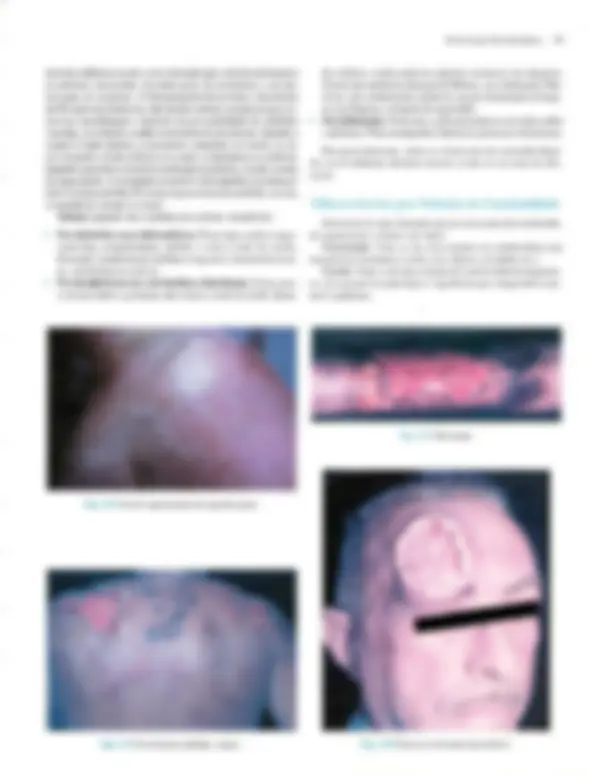
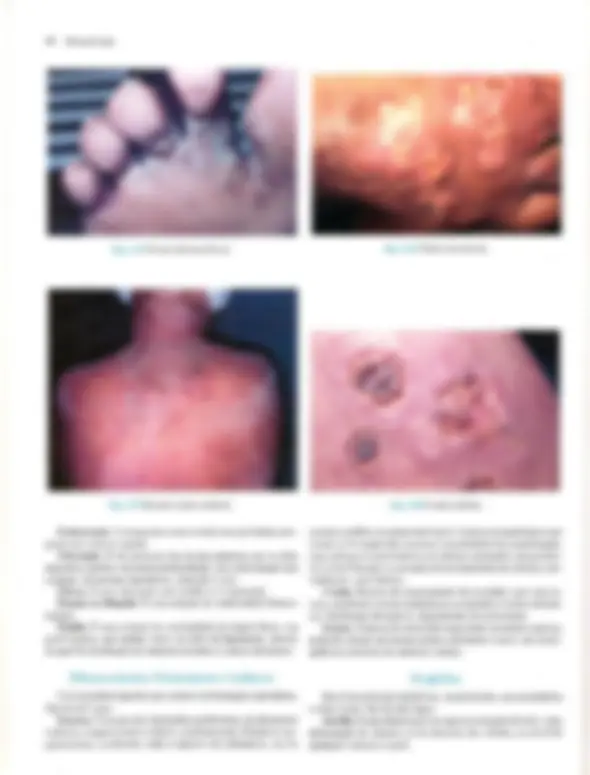
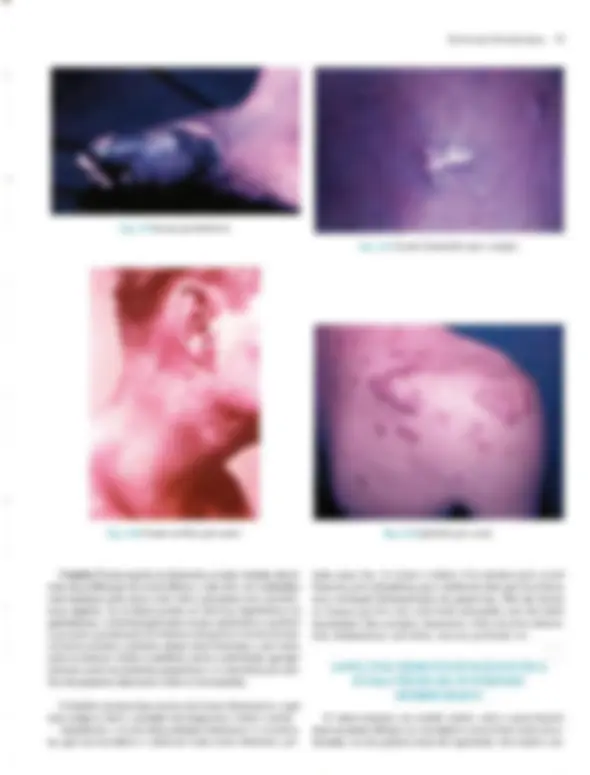
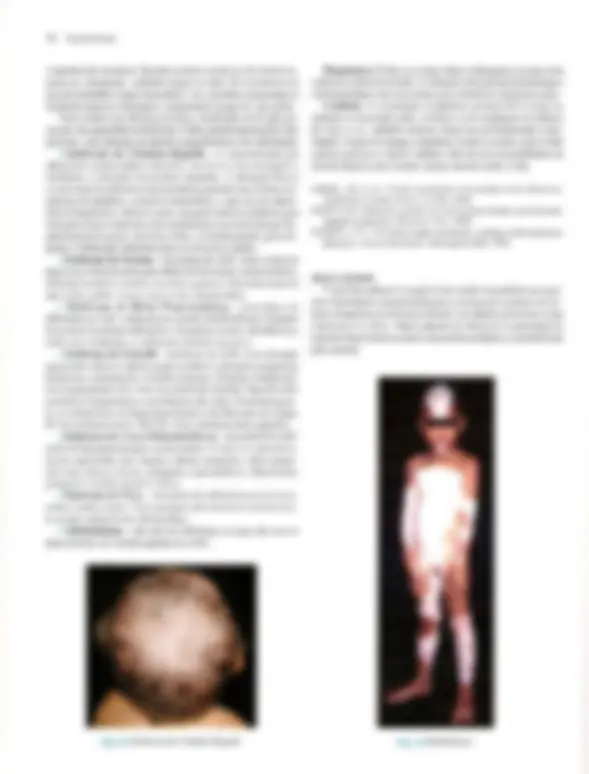
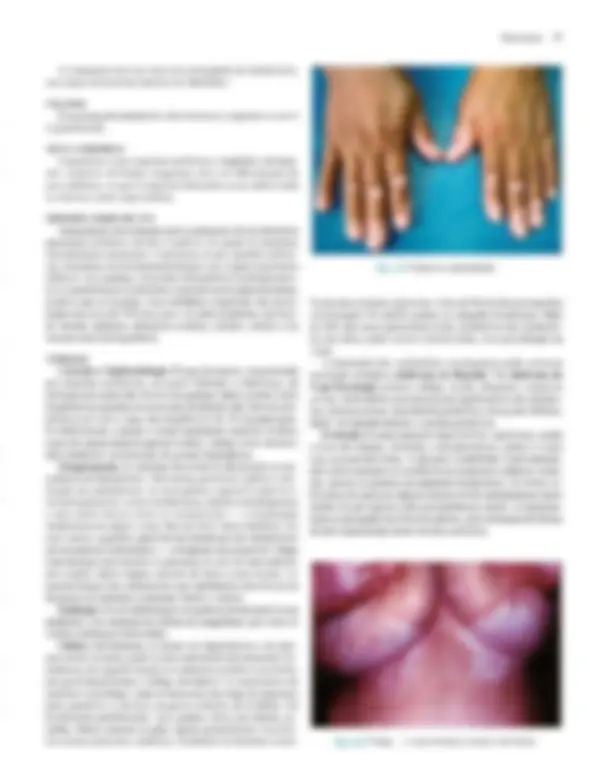
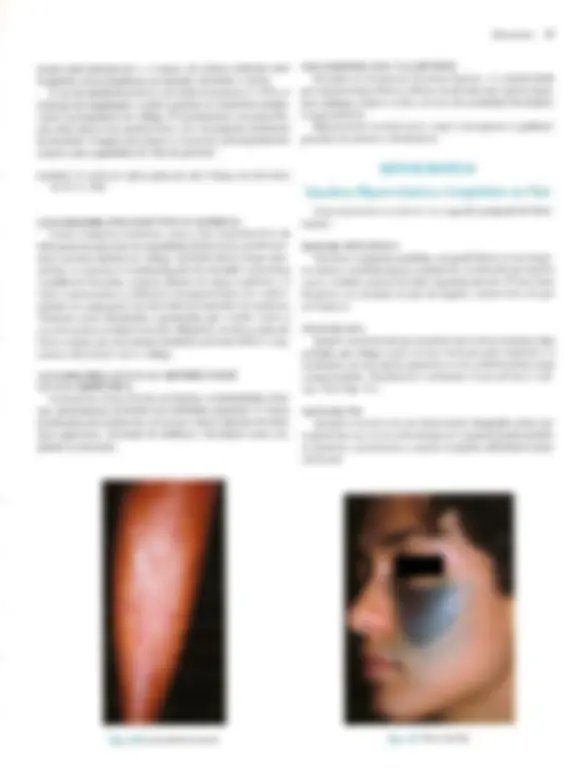
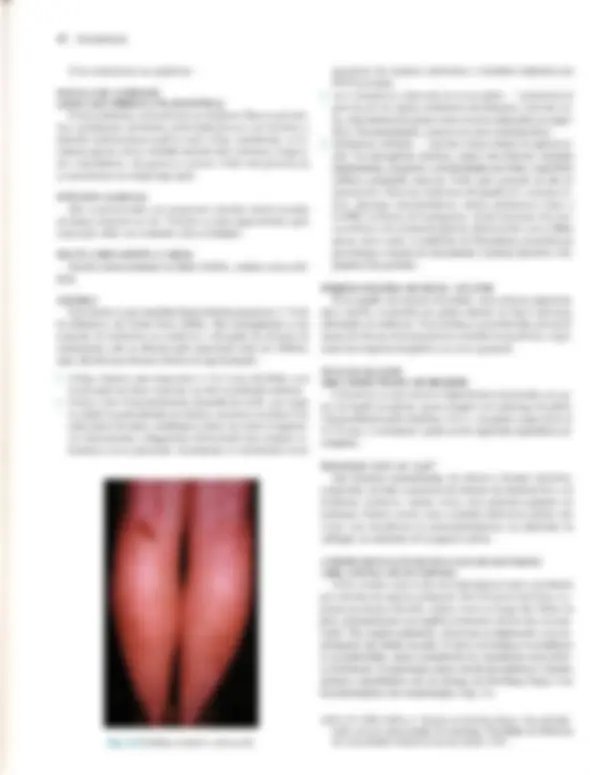
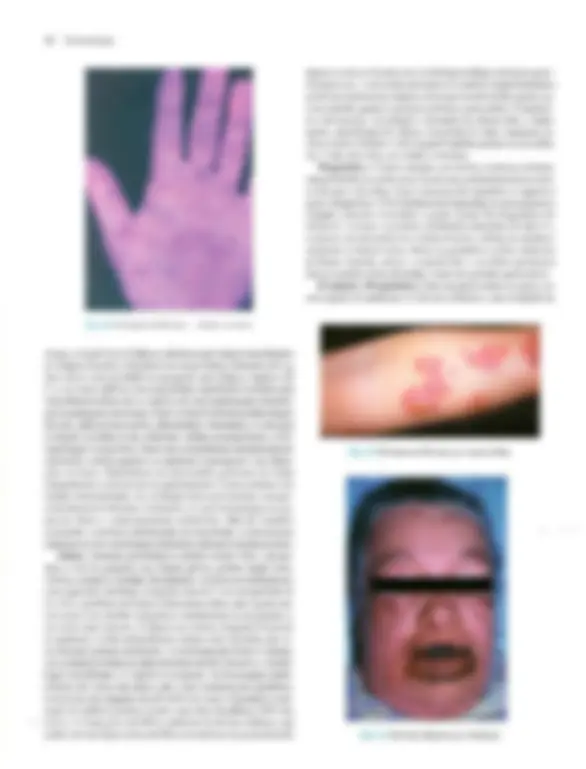
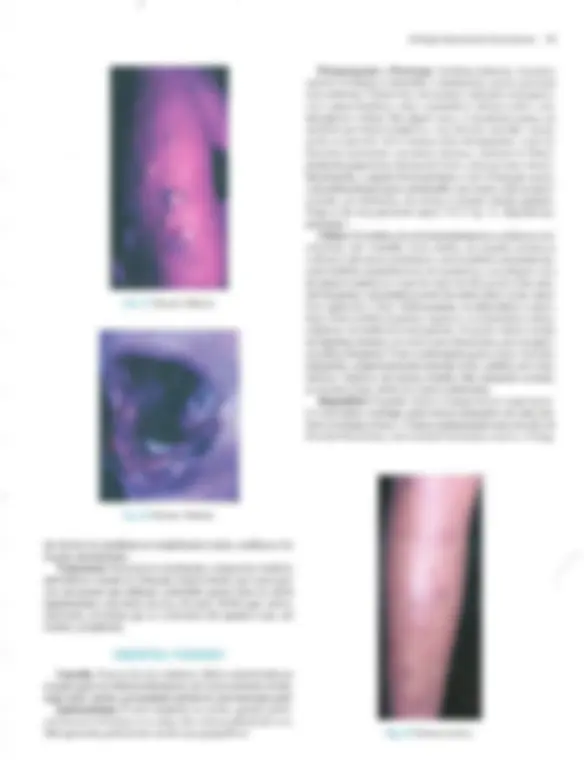
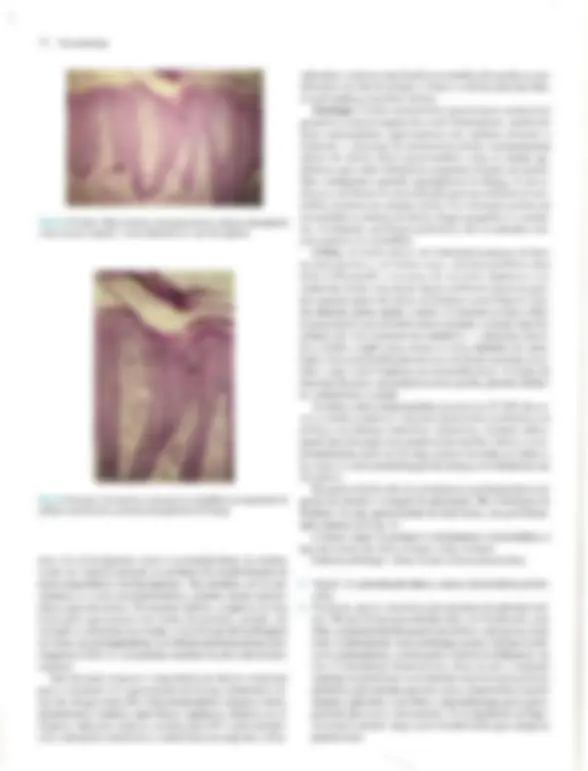
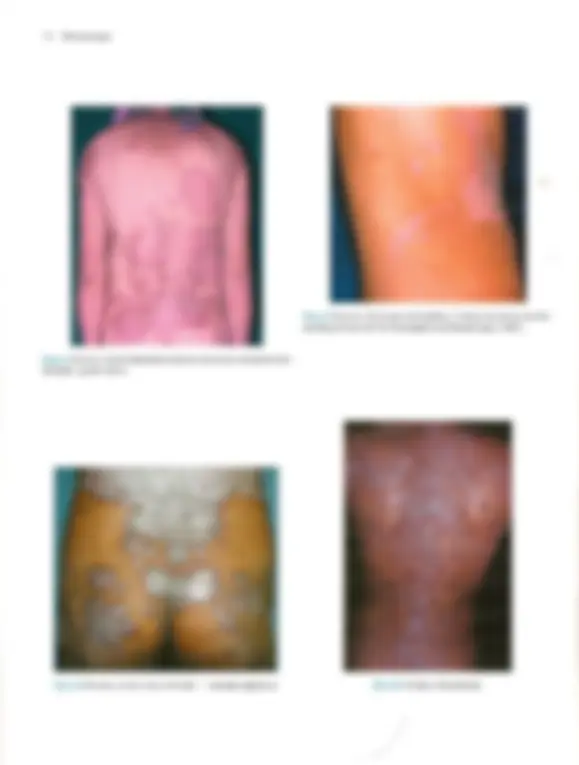
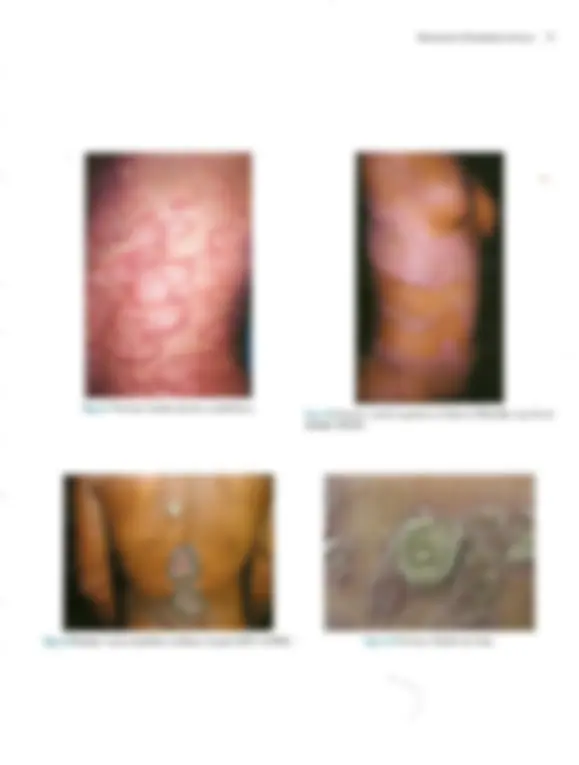
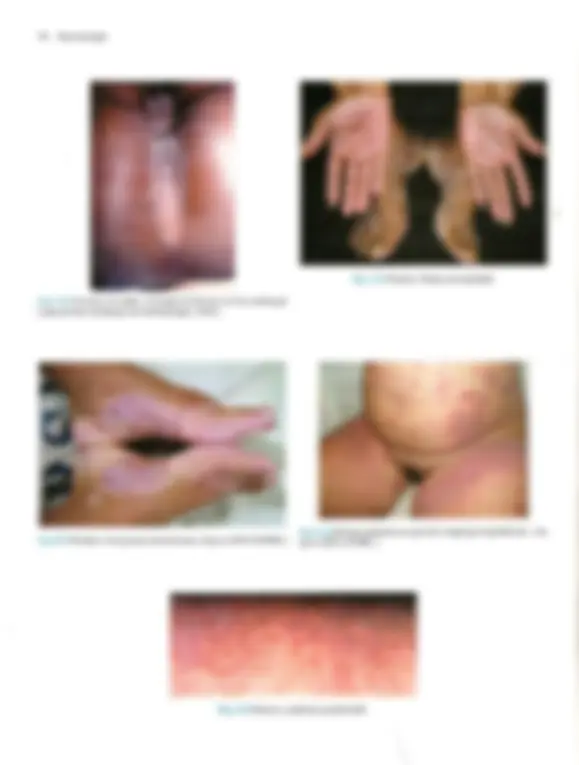
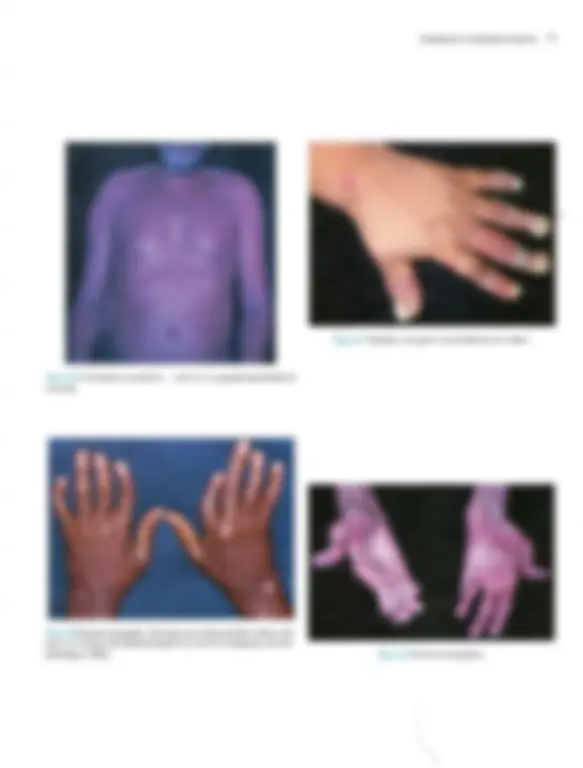
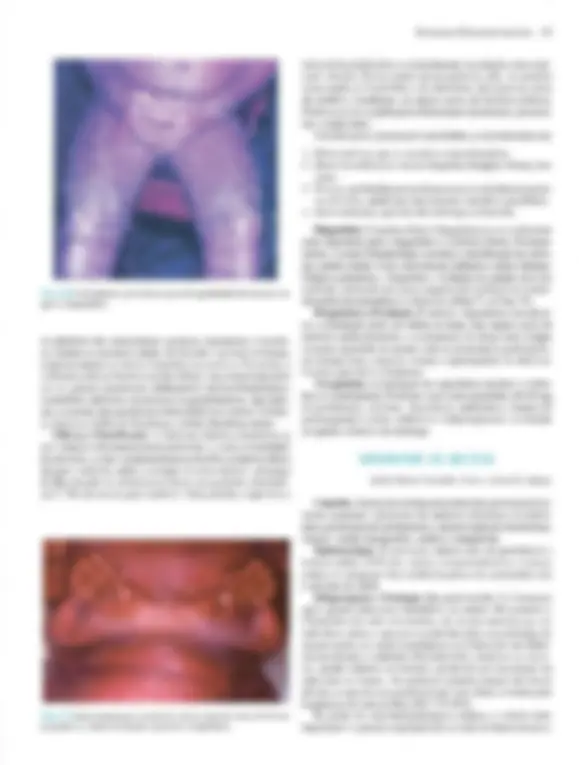
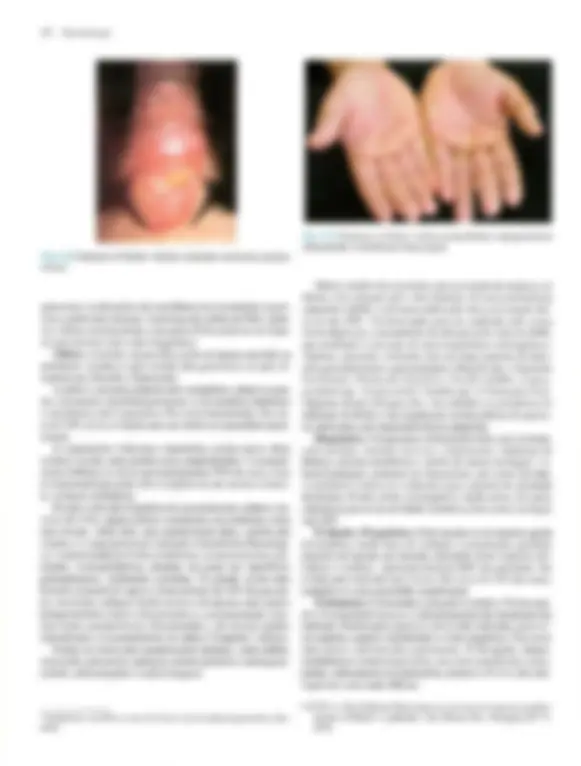
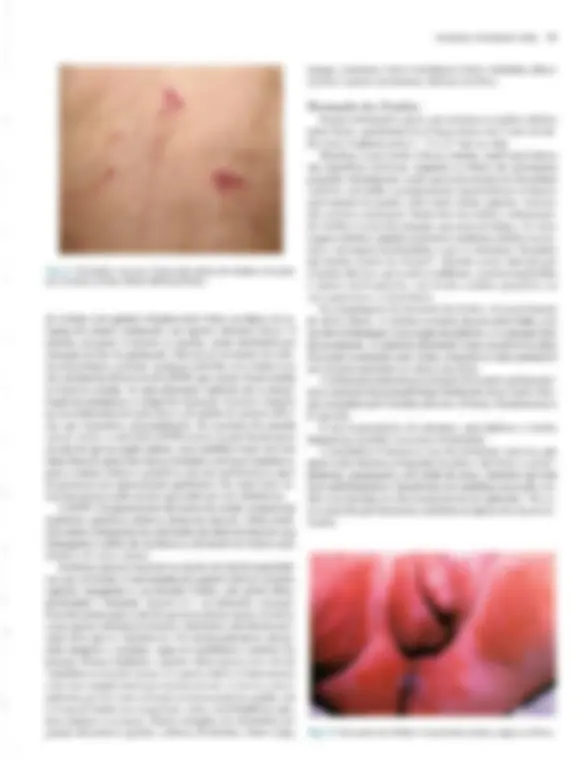
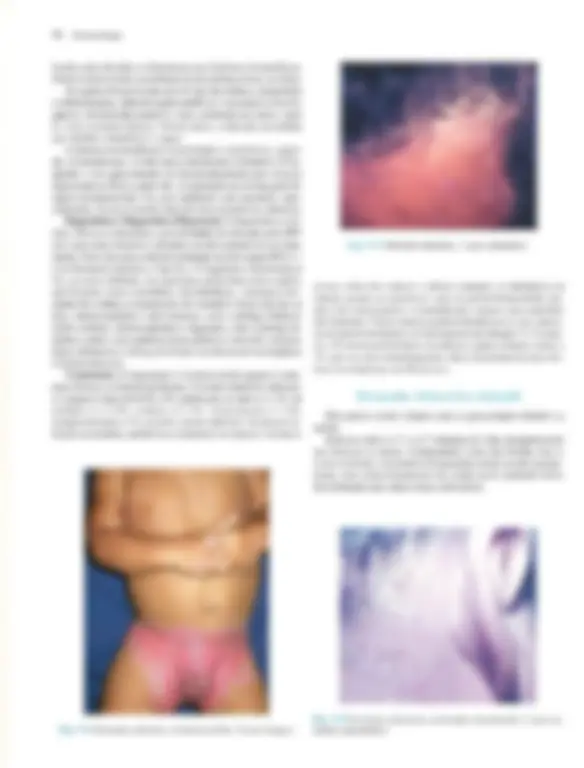
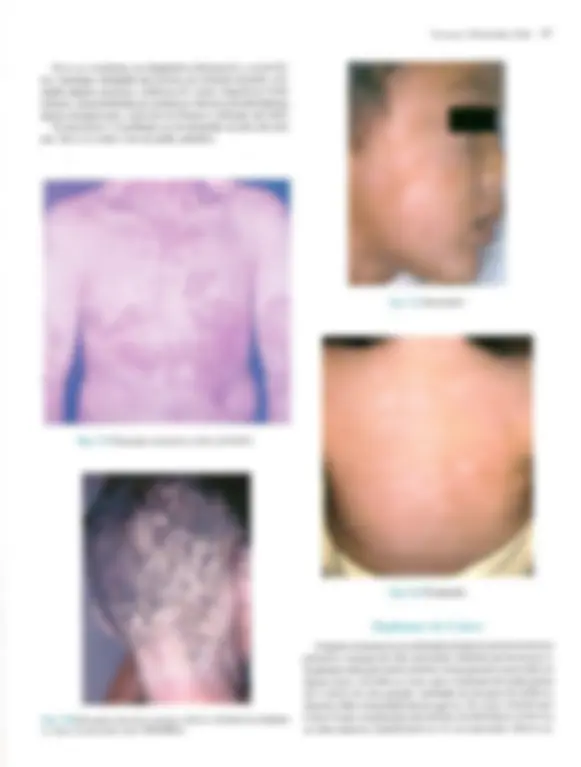
Pré-visualização parcial do texto
Baixe DERMATOLOGIA e outras Notas de estudo em PDF para Enfermagem, somente na Docsity!
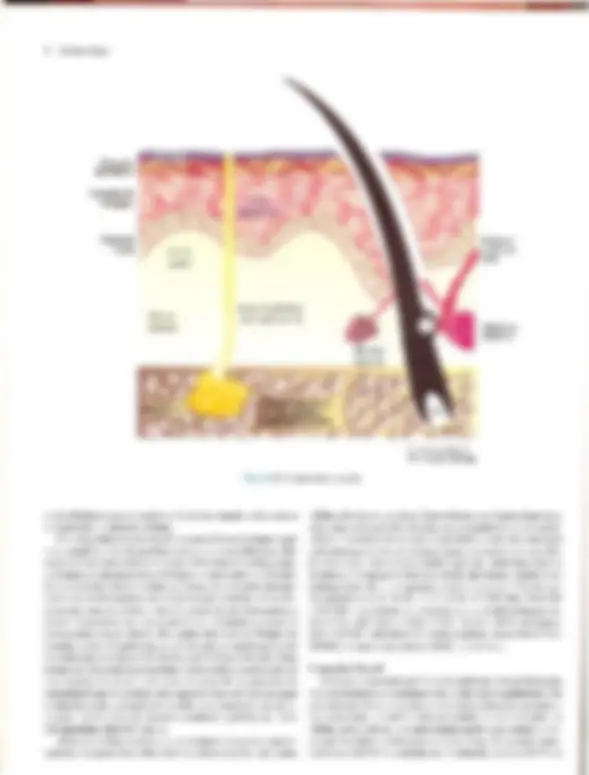
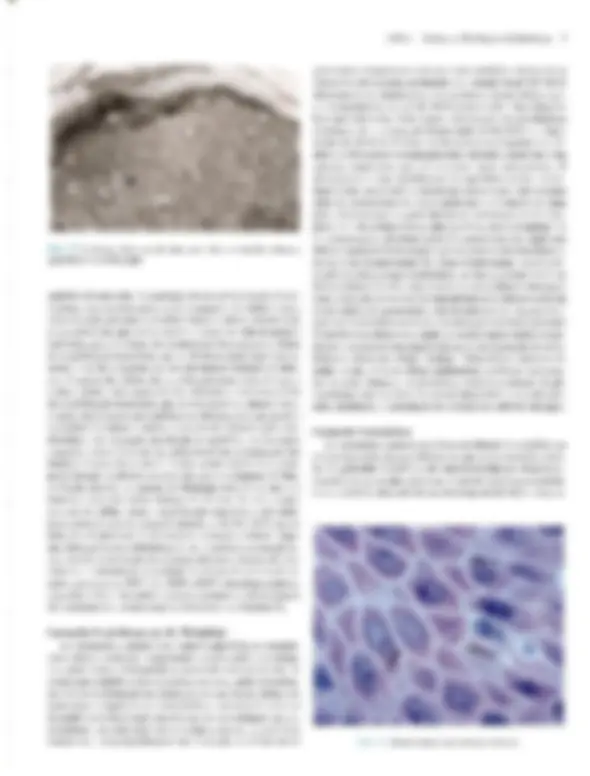
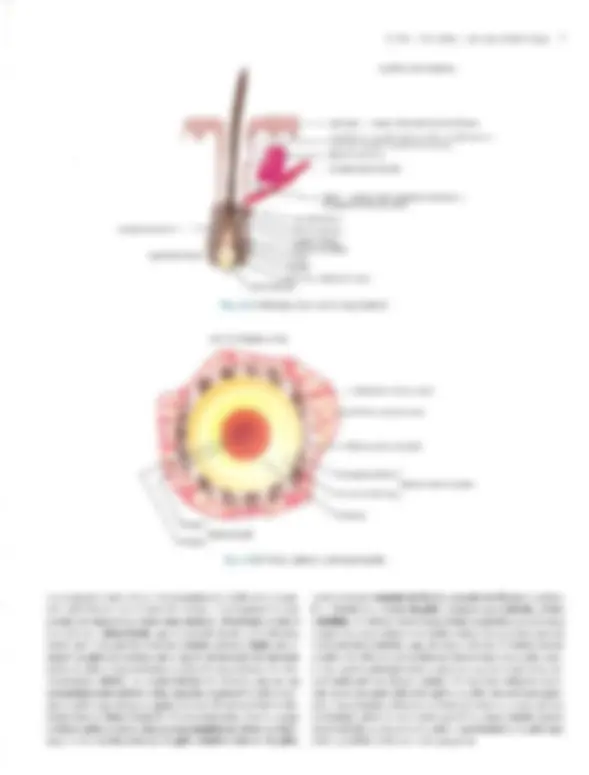
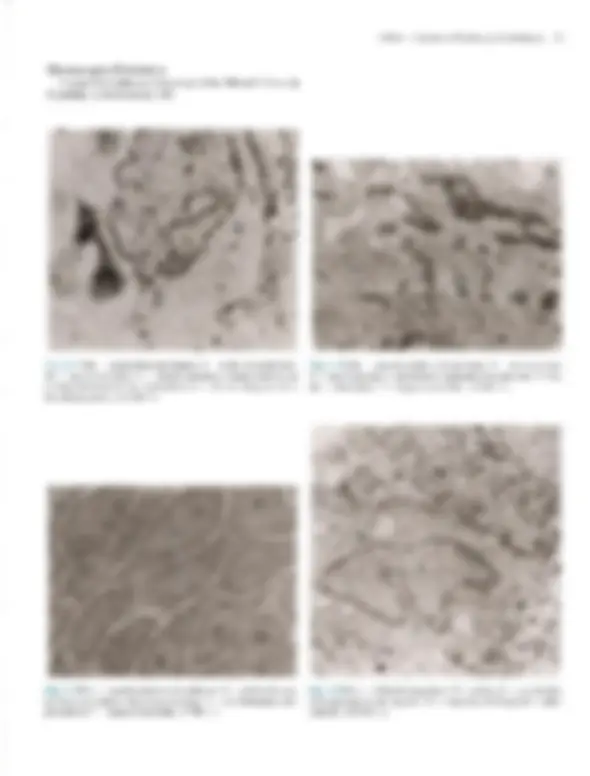
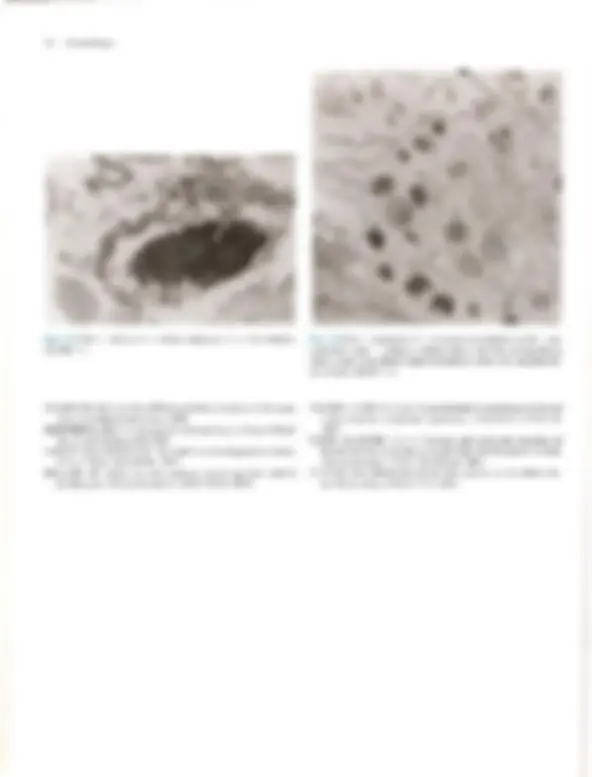
coco A Doomltobogia A Pele — Estrutura, Fisiologia e Capítulo 1 Embriologia Celso Tavares Sodré. David R. Azulay e Rubem D. Azulay A pele do homem, que corresponde a 15% de seu peso cor- poral, é um órgão que reveste e delimita o organismo, protegen- do-o e interagindo com o meio exterior. Pela sua resistência e flexibilidade, determina a sua plasticidade. Caracteristicamente dinâmica, a pele apresenta alterações constantes, sendo dotada de grande capacidade renovadora e de reparação, e de certo grau de impermeabilidade, Tem como função maior e vital a conser- vação da homeostasia (termorregulação, controle hemodinâmi- co e fabrico e excreção de metabólitos). Desempenha, ainda, função sensorial, através dos elementos do sistema nervoso si- tuado na derme, e função de defesa contra agressões físicas, químicas é biológicas, para a qual se destacam, pela sua impor- tância, a ceratinização, o manto lipídico e o seu sistema imuno- lógico. Embora participe de forma absolutamente interativa e interdependente do sistema como um todo, não raras vezes ma- nifestando alterações de órgãos internos, por motivos didáticos iremos abordá-la de modo particular neste capítulo. Não é de- mais chamar a atenção para o fato de que muitas vezes a pele é usada para manifestar condições psíquicas do indivíduo, além de ter conotações de ordem racial, social e sexual. ESTRUTURA E FISIOLOGIA A pele é constituída basicamente por três camadas interdepen- dentes: a epiderme, mais externa, a derme, intermediária, e a hipoderme, mais profunda. A transição entre a epiderme e a der- me é denominada de junção dermoepidérmica ou zona da mem- brana basal. derme É, basicamente, um tecido epitelial estratificado ceratinizado (queratinizado), com variações estruturais e funcionais signifi- cativas na dependência do seu sítio anatômico. É constituída por: sistema ceratinocítico, composto por células epiteliais denomi- nadas ceratinócitos, responsáveis pelo corpo da epiderme é de seus anexos (pêlos, unhas e glândulas); sistema melânico, for- mado pelos melanócitos; células de Langerhans, com função imunológica; células de Merkel, integradas ao sistema nervoso, e células dendríticas indeterminadas, com função mal definida. A derme desempenha uma influência reguladora sobre a morfo- gênese e diferenciação epidérmica, sendo fundamental para a determinação de sua espessura, arquitetura, tipo de diferencia- ção e padrão dos seus anexos. Sistema Ceralinocítico Responsável por, pelo menos, 80% das células epidérmicas, é caracterizado pela disposição lado a lado de suas células e por sua constante renovação. O alto índice de multiplicação celular dos ceratinócitos da sua camada mais profunda, camada basal, fornece as células, que, a seguir, vão gradativamente se modifi- cando (diferenciação) e migrando para a superfície, formando a camada espinhosa ou de Malpighi; essas células, após passa- rem por um rápido estágio em que se apresentam com o citoplas- ma mais basofílico e granuloso, a camada granulosa, transfor- mam-se subitamente em células anucleadas, corneócitos, sendo Fig. 1.1 A Histologia da pele (hematoxilina-cosina) Epiderme: observar, de cima para baixo, camada cómea, à sranulosa e a de Malpighi. epitélios de transição. A população dessas células basais é hete- rogênea; um pequeno percentual é composto de células-tronco caracterizadas por uma velocidade baixa de mitose durante toda a sua existência, que talvez supere 0 tempo de vida do próprio indivíduo, gerando clones de ceratinócitos denominados células de amplificação transitória, que se dividem muito mais rapida- mente, mas são programadas para um número limitado de mito- ses. A mitose das células de amplificação transitória dá origem a duas células com características diferentes: uma nova célula de amplificação transitória, que permanecerá na camada basal, e outra, denominada pós-mitótica ou diferenciada, que perde a capacidade de mitose e inicia o processo de diferenciação cera- tinocítica e de migração em direção à superfície. A renovação completa, desde a divisão da célula basal até a eliminação das lâminas córneas, faz-se em 52-75 dias, assim distribuídos: o tem- po de duração da divisão celular é de aproximadamente 19 dias, o trânsito através da camada de Malpighi dura 26-42 dias e o trânsito através do estrato córneo é de 19 dias. No rato, os per- centuais de células-tronco, amplificação transitória e pós-mitó- ticas corresponderiam, respectivamente, a 10, 50 e 40% das cé- lulas da camada basal. O processo de reparação tecidual e algu- mas doenças hiperproliferativas (p.ex., psoríase) podem provo- car, através da produção de diversas citocinas e fatores de cres- cimento, o aumento na velocidade de mitose das células da ca- mada germinativa. TGF alfa, EGF e KGF estimulam a mitose, enquanto o TGF beta inibe a mitose e promove a diferenciação dos ceratinócitos, assim como os retinóides e a vitamina D,. Camada Espinhosa ou de Malphigi Ao deixarem a camada basal rumo à superfície, os ceratinó- citos sofrem contínuas e importantes modificações morfológi- cas, moleculares e histoguímicas, passando a ser poligonais, de citoplasma acidófilo e ricos em desmossomos, sendo denomina- das células de Malpighi ou células espinhosas. Essas células são numerosas e dispostas em várias fileiras, cujo número varia na dependência da localização anatômica, fatores endógenos (p.ex., hormônios, vascularização etc.) e exógenos (p. ex., ultravioleta, trauma etc.). Ao progredirem na sua migração, as células vão se A Pele — Estrutura, Fisiologia e Embriologia 3 achatando e tomando-se cada vez mais acidófilas. Apesar de os filamentos de ceratina produzidos na camada basal (K5-K14) deixarem de ser sintetizados, eles persistem nessas células, ago- ra acompanhados do par KI-KIO aí produzido, Em situações hiperproliferativas fisiológicas (reparação) ou patológicas (psoríase), há a diminuição da produção de K1-KIO e o surgi- mento de K6-K 16. Embora os desmossomos (espinhos ou nó- dulos de Bizzozero) estejam presentes em toda a epiderme, é na camada espinhosa que se mostram mais numerosos. O desmossomo é uma modificação da superfície celular, cálcio- dependente, responsável pela adesão intercelular; está presente entre os ceratinócitos de toda à epiderme e é formado por uma placa desmossomal na parte interna da membrana celular com- posta por seis polipeptídeos: placoglobina, desmoplaquinas 1 é 1, desmoioguina, proteína banda 6 e a ceratocalmina, sendo esta última responsável pela manutenção da inserção dos tonofilamen- tos na placa desmossomal. Da placa desmossomal, partem cin- co glicoproteínas transmembrânicas, as desmogleínas 1 e 3 e as desmocolinas 1, Ile TIT, cujos domínios extracelulares interagem com os das glicoproteínas transmembrânicas do desmossomo da célula adjacente, garantindo a adesão intercelular. As gap junc- tions são verdadeiros canais de comunicação intercelular por onde fluem diversos elementos, dando um caráter quase sincicial à epi- derme é assumindo um importante papel na regulação do meta- bolismo celular das células vizinhas. Além dessas estruturas de união, existe, entre as células epidérmicas, o cimento intercelu- tar, também chamado de glicocálice. Este é constituído de gli- coproteínas que auxiliam na coesão intercelular, enquanto per- mite, facilmente, a circulação de substâncias solúveis em água. Camada Granulosa Ao deixarem à camada espinhosa em direção à superfície, as células formarão algumas fileiras em que se apresentarão reple- tas de grânulos basofílicos de cerato-hialina no citoplasma, constituindo à camada granulosa. O par de ceratina característi- coéo K2-K11], derivado do metabolismo do K1-K10. Essa ca- 3 Desmossomos (microscopia óptica). 4 Dermatologia M: Membrana celular T: Tonofilamento (ceratina) P: Placa desmossômica PT: Proteínas transmembrânicas (desmogleinas e lll e desmocolinas |, Il e III) Obs.: Desmogleina é um neologismo resultante da associação de desmossomo com giue (cola). osso M: Membrana da célula basal T: Tonofilamento (ceratina) PH: Placa hamidesmossômica constituída por PBAg, (230 kd) e plectina PBAg, (180 kd) ou colágeno Xvil integrina ab. Fil: Filamento de ancoragem LL: Lâmina lúcida LD: Lâmina densa PA: Placa de ancoragem Fig. 14 A, Desmossomo. B, Junção dermoepidérmica. mada caracteriza-se por grande atividade metabólica, objetivan- do a síntese dos elementos necessários ao processo final da cor- mificação, que resultará no súbito surgimento da camada córnea. Esses elementos são armazenados, em grande quantidade, na sua forma pré-ativada, tanto no interior de organelas como livremen- te. Os grânulos de cerato-hialina são constituídos por pró- filagrina, filamentos de ceratina é loricrina. A pró-filagrina irá ser convertida em filagrina (filament agregation protein), que promoverá a agregação e compactação lado a lado dos filamen- tos de ceratina, uma característica dos comeócitos. As proteínas (involucrina, ceratolinina, loricrina, envoplaquina etc.) do enve- lope celular dos corneócitos (camada protéica densa na parte interna da membrana celular) já estão presentes no interior das células granulosas, embora só venham a formar o envelope de- pois de ativadas pelas transglutaminases da membrana celular. Finalmente, os grânulos lamelares (corpos de Odland ou ceratinossomos) morfologicamente semelhantes a lipossomos, contendo no seu interior glicoproteínas, ácidos graxos, fosfoli- pídios, glicosilceramidas e colesterol, são inicialmente vistos nas células das porções superiores da camada espinhosa, mas é na camada granulosa que se apresentam em grande número; o con- teúdo desses grânulos será liberado no espaço intercelular du- rante a transição súbita da camada granulosa para a córnea, quan- do, sob a ação de suas hidrolases, será remodelado, e seus lipídi- os transformados em ceramida (45%), colesterol (25%), ácidos graxos (15%), esfingosina livre, sulfato de colesterol, ésteres do colesterol e triglicerídeos. Todos estes se depositarão em forma de bainha dupla em tomo de cada célula, originando a grande barreira lipídica à passagem de água e substâncias polares da epiderme, principal responsável por sua relativa impermeabili- Fig. 1.5 Epiderme: observar os melanócitos. 6 Dermatologia Fig. 1.6 Unidade cpidermomelânica. Tem sido demonstrado, in vitro, que os ceratinócitos exercem controle sobre a proliferação, diferenciação e atividade dos melanócitos através de fatores mitogênicos, FGFb e TGF alfa, e inibidores de mitose, TL-1, IL-6 e TGF beta. Os melanócitos tam- bém sofrem a ação de hormônios (MSH e hormônios sexuais), mediadores inflamatórios e vitamina D3 produzida na epiderme. O bronzeamento produzido pelo sol deve-se a uma excitação da tirosinase pelo ultravioleta, levando à formação de melanosso- mos maiores e mais numerosos. CÉLULA DE LANGERHANS E CÉLULA INDETER- MINADA. Há, ainda, na epiderme, duas outras células dendríticas: a célula de Langerhans, que se cora pelos sais de ouro, ea célula dendrítica indeterminada, que só é percebida pela mi- croscopia eletrônica. A célula de Langerhans é um dos principais componentes do sistema imunológico da pele, sendo responsável pelo reconhecimen- to, intemalização, processamento e apresentação de antígenos solú- veis e haptenos presentes na epiderme. Originada na medula óssea, corresponde a 2-8% das células epidérmicas e distribui-se da cama- da basal à granulosa, tendo preferência pela posição suprabasal, Não estabelece adesão através de desmossomos com os ceratinócitos. Na microscopia óptica de rotina, pode ser visualizada como uma célula de núcleo convoluto e citoplasma claro; na microscopia eletrônica são encontradas pequenas estruturas assemelhadas a raquetes, denominadas grânulos de Birbeck, que funcionariam como fagoli- sossomos. A célula de Langerhans é uma célula processadora-apre- sentadora de antígenos; em seu estágio não ativado tem significati- vaação fagocítica, internalizando antígenos e processando-os, o que provoca transformações que a fazem perder o potencial fagocítico, mas que aumentam sua capacidade apresentadora de antígeno para os linfócitos Te, desse modo, produzem uma resposta imunológi- ca. As células de Langerhans encontram-se reduzidas em número em algumas condições, como psoríase, sarcoidose, dermatite de contato e após irradiação com UV: neste último caso, também es- tão prejudicadas funcionalmente. Alguns de seus marcadores his- toquímicos de superfície são: ATP-ase, CDla, CD4, CD45, S100, T200, HLA-DR, DLA-DQ, DLA-DPe receptores de Fee C3. Além da epiderme, as células de Langerhans podem ser encontradas em outros epitélios (mucosas), órgãos linfóides (baço, timo e Tinfono- dos) e na derme normal. As células dendríticas indeterminadas não têm melanossomos nem grânulos de Langerhans, porém há autores que julgam que possam dar origem a melanócitos ou a células de Langerhans. Não-dendríticas CÉLULA DE MERKEL. À célula de Merkel, relativamente rara e aparentemente derivada de uma célula-tronco epidérmica, localiza- se entre as células basais, às quais está aderida por desmossomos. Funciona como um tipo de mecanorreceptor de adaptação lenta em locais de alta sensibilidade tátil, parece ser estimulada pela defor- mação nos ceratinócitos adjacentes, provocada por contatos exter- nos, respondendo com a secreção de transmissores químicos nas sinapses que estabelece com as terminações nervosas livres da jun- ção dermoepidérmica. Em determinadas localizações, organiza-se em estruturas especializadas denominadas discos táteis. Tem núcleo lobulado ou oval e citoplasma claro, onde são encontrados grânulos osmiófilos produzidos no aparelho de Golgi, que contêm neurotrans- missores, Seus marcadores imuno-histoquímicos são os filamentos de ceratina K8, KI8. K19e K20, sendo este último exclusivo dela. Na microscopia óptica, a impregnação pela prata revela os discos de Merkel, que nada mais são que a porção inferior da célula. OS ANEXOS CUTÂNEOS. Folículo pilossebáceo, glându- las sudoríparas e unha. São estruturas que surgem de modifica- ções da epiderme ainda na vida embrionária. FOLÍCULO PILOSSEBÁCEO. Composto de folículo pilo- so, glândula sebácea e músculo eretor do pêlo, é sempre provido de terminações nervosas; em algumas regiões (axilas, púbis, mama etc.) desembocam no folículo as glândulas sudoríparas apócrinas. FOLÍCULO PILOSO, Forma-se na vida embrionária como uma projeção de ceratinócitos modificados (tricócitos) para den- tro da derme; isso ocorre por influência de células mesenquimais que, mais tarde, constituirão a papila folicular, que se localizará Fig. 1.7 Folículos pilossebáceos. A Pele — Estrutura, Fisiologia e Embriologia 7 CORTE LONGITUDINAL acotríquio — porção intra-apidérmica do folículo infundíbulo — porção entra a saída na epiderme e o ponto de inserção da glândula sebácea glândula sebácea músculo eretor do pêlo istmo — porção entre à glândula sebácea e a inserção do músculo ereior Fig. 1.8 A Folículo piloso (corte longitudinal). CORTE TRANSVERSAL Haste do pólo Medula —+ Membrana vítrea acelular Células mesenquimais Fig 1.8 B Folículo piloso (corte transversal). na sua porção mais inferior. Funcionalmente, o folículo e sua pa- pila estabelecem uma relação tão íntima & interdependente que podem ser entendidos como uma unidade. Histologicamente é dividido em: infundíbulo, que se estende desde a sua abertura (Óstio) até a inserção do ducto da glândula sebácea; istmo, da in- serção da glândula sebácea até a região da inserção do músculo eretor do pêlo e da protuberância (área de concentração de célu- las-tronco); e bulho, ou porção inferior do folículo, que, na sua extremidade mais inferior, tem a matriz, responsável pela produ- ção do pêlo e que abraça a papila folicular. Os melanócitos se dis- põem entre as células da matriz, Transversalmente, observa-se que o folículo piloso é circundado por uma membrana vítrea, celular, seguida da: bainha externa do pêlo; bainha interna do pêlo, constituída por camada de Henle, camada de Huxley e cutícu- la; e, finalmente, a haste do pêlo, composta por cutícula, córtex e medula. A linha de Auber é uma linha imaginária que atravessa aregião de maior diâmetro do bulbo, abaixo da qual está a área de maior atividade mitótica, que dá origem à haste e à bainha interna do pêlo. Os folículos pilossebáceos existem em toda a pele, exce- to nas regiões palmoplantares e algumas regiões da genitália, de- nominadas por isso de pele glabra. Apresentam variações regio- nais; assim, no couro cabeludo e púbis, os pêlos são terminais (gros- sos), com glândulas sebáceas também bem desenvolvidas; na face predomina o pêlo tipo velo ou lanugo (fino), com glândulas muito desenvolvidas; e, nas extremidades, o predomínio é de pêlo tipo velo e glândulas sebáceas muito pequenas. Eponíquio Lâmina ungueal Fig. 1.10 Corte longitudinal da unha. noradrenalina, como neurotransmissor; a acetilcolina estimula a secreção do suor e a contração das células mioepiteliais. promovendo a sudorese, que atinge, em média, 10 lídia numa pessoa bem aclimatizada. O suor, que inicialmente é semelhante a um ultrafiltrado isotônico do plasma, composto por NaCI, K, HCO,, lactato, uréia, amônia e traços de aminoácidos e proteínas, após a reabsorção de NaC] e HCO, pelas células ductais dá origem à se- ereção hipotônica que chega na superfície. UNHA. As unhas são lâminas de ceratina que recobrem as últimas falanges, e originam-se na imatriz ungueal, Além da matriz, devemos considerar os seguintes componentes: lúnula, eponíquio, lâmina ungueal, leito ungueal e hiponíquio, bem re- presentados no esquema anexo da Fig. 1.10. Crescem de mam ra contínua, em média 0,5-1,2 mm por semana. (Ver Cap. 36.) Derme A derme é uma camada de tecido conjuntivo composta por um sistema integrado de estruturas fibrosas, filamentosas e amorfas, na qual são acomodados vasos, nervos e anexos epidér- micos. Fibroblastos, histiócitos, células dendríticas e mastócitos são suas células residentes, enquanto linfócitos, plasmócitos e outros elementos celulares do sangue são encontrados em número variável e de forma transitória. Sua interação com a epiderme é fundamental para a manutenção dos dois tecidos; ambos cola. boram na formação da junção dermoepidérmica e dos anexos epidérmicos, assim como no processo de reparação da pele. A derme pode ser dividida em três partes distintas: (1) superficial ou papilar, com grande celularidade e onde predominam finos feixes fibrilares de colágeno dispostos mais verticalmente; (2) profunda ou reticular, constituída por feixes mais grossos de colágeno, ondulados e dispostos horizontalmente; (3) adventicial, disposta em tomo dos anexos e vasos e constituída de feixes fi- nos de colágeno, como na derme papilar. Colágeno Proteína distribuída amplamente pelos tecidos conjuntivos, corresponde a aproximadamente 75% do peso seco da derme e provê resistência e elasticidade ao tecido; cora-se bem pela cosi- na; suas fibras variam em diâmetro entre 2 e 15 pum e apresentam- se em rede ondulada fina, na derme papilar. ou espessa, na derme reticular. Os diversos tipos de colágeno têm em comum o fato de A Pele — Estrutura, Fisiologia e Embriologia 9 serem compostos por três cadeias polipeptídicas (cadeias alfa), com uma sequência fixa de glicina a cada 3.º posição, intercaladas por dois outros aminoácidos, que, muito fregiientemente, são a prolina ea hidroxiprolina; essa composição faz com que as três cadeias se entrelacem de modo helicoidal, à semelhança de uma corda, au- mentando a resistência da molécula à tração. No ser humano exis- tem pelo menos 19 tipos de colágeno, designados de L a XIX. O colágeno tipo Lresponde por 80% do colágeno dérmico do adulto, enquanto o tipo TII, que predomina na vida embrionária, represen- ta 10% do colágeno da vida adulta. O tipo TV compõe as membra- nas basais, inclusive a lâmina densa da junção dermoepidérmica: astipos V e VI são ubíquos, estando presentes na derme em quan- tidade relativamente pequena. O tipo VIT é o principal componen- te das fibrilas de ancoragem da junção dermoepidérmica, tendo sido identificado como o antígeno da epidermólise bolhosa adquirida. O tipo XVII localiza-se na porção intracelular dos hemidesmos- somos e corresponde ao antígeno penfigóide 2 de 180 kd. O colá- geno é secretado para o espaço extracelular como pró-colágeno; este sofre a ação de enzimas, provocando a formação de fibras, e estas se unirão para formar os feixes. Do equilíbrio dinâmico da síntese e degradação do colágeno, depende a normalidade do teci- do nas situações fisiológicas e de reparação. Fibras Elásticas As fibras elásticas não se coram pelas técnicas rotineiras, mas são reveladas pela orceína ou resorcina-fucsina. Estão presentes em quase todos os órgãos em proporções variáveis. Na pele, essas fibras formam uma rede que se estende da junção dermoepidér- mica ao tecido conectivo da hipoderme, estando elas também pre- sentes na parede dos vasos e em torno do folículo piloso; corres- pondem apenas a 1 à 2% do peso seco da derme e entremeiam-se com as fibras colágenas. O sistema elástico é composto por três tipos de fibra: as finas fibras oxitalânicas e eulanínicas, e as fibras elásticas maduras, que são mais espessas. As fibras oxitalânicas são encontradas na derme papilar, dispostas verticalmente a partir da junção dermoepidérmica, e estão conectadas a uma rede hori- zontal de fibras eulanínicas na interface entre derme papilar e re- ticular, essas fibras se relacionam com a rede de fibras elásticas maduras, também disposta horizontalmente, que atravessa toda a derme reticular. Os três úpos de fibra formam uma trama com funções de ancoragem e de oposição às forças de distensão e com- pressão. Embora sejam sintetizadas por diversos tipos de célula, inclusive ceratinócitos, na pele a principal fonte de fibras elásti- caséo fibroblasto, sendo sua renovação muito lenta. Pela micros- copia eletrônica, venfica-se que as fibras elásticas maduras são constituídas de microfibrilas com 10-12 nm de diâmetro e de uma substância protéica amorfa central, denominada elastina, enquan- to as fibras oxitalânicas são um agrupamento de microfibrilas en- volvido por elastina. A elastina é extremamente insolúvel devido a ligações covalentes complexas dependentes de cobre, denomi- nadas desmosinas; especula-se que, durante a distensão da fibra, os grupos hidrofóbicos da elastina sejam expostos ao meio aquo- so, e que a energia para a contração da fibra seja derivada do re- torno desses grupos a sua situação não-polar. Tem sido observada atividade de elastase nas bolhas da dermatite herpetiforme e do penfigóide bolhoso, possivelmente responsável pela dissolução das fibras elásticas e outras estruturas da junção dermoepidérmica. 10 Dermatologia Proteoglicans e Glicosaminoglicans — Substância Fundamental A substância fundamental ou mucopolissacarídeo é constitu- ída por proteoglicans. A sua capacidade de ligação a múltiplos componentes do meio extracelular promove a aderência neces- sária entre diversas estruturas (células, fibras, fatores de cresci- mento, integrinas, água etc.), desempenhando papel fundamen- tal na organização e viabilidade dos tecidos. Embora correspon- dam apenas à 0,2% do peso seco da derme, são capazes de ligar até mil vezes o seu volume em água, influenciando o conteúdo de água da derme, a sua compressibilidade e volume. Os proteoglicans se caracterizam por serem compostos de um eixo protéico ao qual uma ou mais diferentes cadeias de carboidra- tos, os glicosaminoglicans, se ligam. Os principais glicosamino- glicans são o áeido hialurônico, sulfato de heparana, sulfato de condroitina, sulfato de keratano e sulfato de dermatan (condroi- tina sulfato B). O ácido hialurônico é o único capaz de se apre- sentar sem estar associado ao eixo protéico; é o mais simples deles e nunca é sulfatado; predomina na pele fetal e nos processos de reparação; quando é necessário facilitar a migração celular, cora- se pelo alcian blue é apresenta metacromasia quando submetido ao azul de toluidina em pH 3,0; os outros glicosaminoglicans são sulfatados em maior ou menor proporção e têm as mesmas pro- priedades tintoriais que o ácido hialurônico em pH 3,0 ou pHO,S. As metaloproteinases da matriz extracelular (colagenases, gelatinases, estrom inas, matrilisina, metaloelastase e metaloproteinase da matriz da membrana) são as enzimas res- ponsáveis pela degradação das macromoléculas da matriz extra- celular. Elas são sintetizadas e secretadas sob a forma pró-ativa- da por diversos tipos de células, de especial interesse o fibroblas- to; são zinco e cálcio dependentes é sua produção está sob con- trole de fatores estimuladores (TL-1, EGF, PDGF, TNF alfa, ul- travioleta) e inibidores (corticóide, TGF beta e ácido retinóico). Células da Derme As células da derme são representadas por células mesenqui- mais primitivas, fibroblastos, macrófagos/histiócitos e mas- tócitos. As células mesenquimais primitivas são as únicas existen- tes no início da vida fetal, diferenciando-se depois em outras células. Em certas condições patológicas, essas células, de mor- fologia dendrítica, são ativadas, dando origem às células das li- nhagens: histiocítica, linfocítica e granulocítica. Os fibroblastos são células fusiformes e estreladas, com nú- cleo volumoso é citoplasma claro, com retículo endoplasmático nítido e granular; têm grande ação enzimática, sendo os princi- pais responsáveis pela síntese e degradação das proteínas do te- cido conectivo e de vários fatores solúveis que funcionam come mensageiros para a epiderme, vasos e outras células. Formam uma grande família de células que, embora provenientes da mesma célula-tronco mesenquimal, se diferenciam em tipos es- pecializados em diferentes funções. Fibroblastos respondem a vários mediadores imunológicos, incluindo TL-1 alfa e beta, que estimulam a síntese de KGF, IL-1 alfa e beta e TL-8. Os histiócitos/macrófagos e dendrócitos dérmicos são os representantes dérmicos do sistema retículo-endotelial, deriva- dos de células precursoras da medula óssea. Os hist crófagos têm capacidade de fagocitar, apresentar antígeno, mi- crobicida, tumoricida, secretar moléculas imunomoduladoras, citocinas e fatores de crescimento, além de propriedades hema- topoiéticas. Morfologicamente semelhantes ao fibroblasto, são diferenciados por apresentarem, na sua superfície, os antígenos CDile, CD6 é KiM8. Em algumas condições patológicas, ori- ginam as células epitelióides e gigantes dos granulomas. Os dendrócitos dérmicos são macrófagos apresentadores de antí- geno, localizados em maior número nas porções superiores da derme, principalmente ao redor dos vasos. Os mastócitos podem ser identificados por seu núcleo arredon- dado e pelos abundantes grânulos escuros do seu citoplasma, que, corados pelo Giemsa e outros métodos, apresentam metacroma- sia, isto é, têm cor diferente da do corante; essa capacidade de mo- dificar a cor do corante deve-se à presença de mucopolissacaríde- os ácidos. Os grânulos são de dois tipos: secretórios e lisossomai os primeiros contêm heparina, histamina, triptase, quimase, car- boxipeptidase, fator quimiotático para neutrófilos e fator quimio- tático para eosinófilo, enquanto os segundos contêm hidrolases ácidas que digerem intracelularmente glicosaminoglicans, proteoglicans e complexos glicolipídicos. Os mastócitos também são capazes de secretar alguns fatores de crescimento, citocinas, leucotrienos e fator ativador de plaqueta. Com esse painel tão eclético de produtos, o mastócito desempenha um importante pa- pel na reparação do tecido, reação de hipersensibilidade do tipo L, defesa contra parasitas. quimiotaxia, ativação e proliferação de eosinófilos, promoção da fagocitose, permeabilidade vascular, ação antitumoral (TNF alfa) e angiogênese, É originado das células-tron- co CD34+ da medula óssea, é diferencia-se em mastócito quan- do, no tecido, sofre ação de fatores produzidos por outras células, como a IL-3. O mastócito é encontrado em maior quantidade na derme papilar, em torno dos anexos e nos vasos e nervos do plexo subpapilar. Em condições patológicas, encontramos na derme cé- lulas da série micloblástica (mielócitos, neutrófilos, cosinófilos) e da linforreticular (linfócitos T e B). Vascularização A rica vascularização sangiiínea da pele supera o necessá- rio ao seu suprimento metabólico, fato justificado pelo papel que desempenha na regulação da temperatura, pressão arterial, cica- trização é fenômenos imunológicos. Os vasos sangúíneos da derme (arteríolas, capilares arteriais e venosos e vênulas) estão distribuídos em duas redes horizontais ligadas por vasos comu- nicantes, os vasos perfurantes dos músculos subjacentes dão origem ao plexo inferior, no limite com a hipoderme, deste deri- vam vasos que ascendem até o plexo superior e outros que su- prem os anexos; o plexo superior ou subpapilar, entre a derme papilar e a reticular, dá origem aos capilares das papilas dérmicas. Devemos destacar, na derme, certos corpos vasculomusculares que unem, facultativamente, arteríolas a vênulas diretamente; os glomos, que se localizam na derme reticular, sobretudo nos dí- gitos, são ricos em células musculares e têm por finalidade man- ter à termorregulação e a homeostasia. A rede linfática, exclusivamente coletora, inicia-se nos capi- lares linfáticos com fundo cego, presentes na derme papilar, que 12 Dermatologia A derme e a hipoderme diferenciam-se a partir do mesoder- ma, sendo inicialmente representadas por um tecido afibrilar e amorfo, a substância fundamental, nesse momento constituído quase que só por ácido hialurônico e por células mesenquimais primitivas de morfologia dendrítica. No 3.º mês de vida, surgem as primeiras fibras argentafins; são as fibras reticulares ou colá- geno TI, que podem proliferar no adulto em determinadas condi- ções patológicas devido à hiperatividade dos fibroblastos (sar- coidose, dermatofibromas, sarcomas, linfomas etc.); com o cor- rer do tempo, a argirofilia tende a desaparecer e surgem os fei- xes de fibras colágenas. Ao mesmo tempo, as células mesenqui- mais assumem o aspecto fusiforme dos fibroblastos, que presi- dem todas as alterações histoguímicas. Só mais tarde, em torno da 22.º semana, aparecerão as fibras que se impregnam pela orceína — as elásticas. No 5.º mês, começa a formar-se a hipoderme através do acúmulo cada vez maior de lipídios no interior de células, que passam, então, a ter uma morfologia ar- redondada, com o núcleo rebatido para a periferia — são os adi- pócitos. Os vasos surgem no mesênquima em torno do 3.º mês de vida, e, inicialmente, são do tipo capilar, mais tarde diferenciam-se em artérias e veias. As estruturas nervosas começam a aparecer em tomo da 5.º semana e têm diversas origens: as células de Schwann originam- se da crista neural, enquanto os axônios provêm da medula e dos gânglios posteriores. Aos 4 meses, já estão esboçados os nervos, que, inicialmente, são amielínicos, só mais tarde surgindo a mielina. FUNÇÃO DA PELE Em toda a escala animal, a pele exerce funções diversas que vão se modificando filogeneticamente; a pele exerce funções res- piratórias em determinados animais, do mesmo modo que a pro- teção física é altamente desenvolvida em outros (escamas, nos peixes), bem como atração sexual pelo cheiro das secreções glan- dulares. No homem, a pele é um órgão de grande importância, pois visa a manter um equilíbrio com o meio exterior, no sentido da manutenção vital do meio interior. Destacaremos as seguintes fun- ções * Proteção. É exercida das mais diversas maneiras contra as agressões do meio exterior. A pele tem uma resistência rela- tiva aos agentes mecânicos, por sua capacidade moldável e elástica (fibras colágenas, elásticas e hipoderme). No sentido físico, essa proteção se faz pela capacidade de, através de seu sistema melânico, impedir e absorver as radiações calóricas ultravioleta e, até mesmo, ionizantes (parcialmente). Cabe sa- lientar que a produção de melanina, além do controle genéti- co e ambiental, sofre interferência da porção intermediária da hipófise através do hormônio intermedina, ou MSH. Por ou- tro lado, a melatonina, produzida pela hipófise através da ação da noradrenalina, faz o clareamento da pele por induzir a agre- gação dos grânulos de melanina em torno do núcleo das célu- las. Pela sua relativa impermeabilidade à água e aos eletróli- tos, a pele mantém o equilíbrio hidroeletrolítico, Outros tipos de proteção são: a físico-química, no sentido da manutenção do pH ácido (5,4-5,6) da camada córnea; a química, através do manto lipídico com atividade antimicrobiana; e a imuno- lógica, presente, na epiderme, através das células de Lan- gerhans e, na derme, à custa de macrófagos, linfócitos e mas- tócitos. e Percepção. Os elementos nervosos que existem, sobretudo na derme, permitem o reconhecimento de sensações especiais, como calor, frio, dor e tato, o que conduz a um mecanismo de defesa no sentido de sobrevivência. * Hemorregulação e termorregulação. A pele, através de seus extensos plexos vasculares e corações periféricos (os glomos), colabora na manutenção e regulagem do débito circulatório. Em determinadas ocasiões, o aumento do débito sangúíneo periférico é compensado pela constrição dos glomos, com desvio da circulação para a rede capilar, e pela utilização ple- na da capacidade total de enchimento de outros vasos, já no choque, a dilatação dos glomos e constrição dos vasos cutá- neos provocam a palidez característica, que denuncia a ele- vada função hemorreguladora da pele. A homeoterímia ou termorregulação é mantida por mecanismo comandado pelo centro termorregulador através das vias do sistema nervoso autônomo, levando à vasoconstrição ou vasodilatação. Além disso, os vasos são sensíveis a duas substâncias químicas cir- culantes: a noradrenalina e a acetilcolina. No mecanismo de termorregulação, exercem uma ação especial as glândulas sudoríparas écrinas, que, sob estímulo colinérgico, aumentam a sudorese, causando a perda de calor. * Secreção. Como elementos produzidos pela pele, destacamos aceratina, a melanina, o sebo e o suor, todos exercendo fun- ções definidas e harmônicas. O sebo colabora na formação do manto lipídico com atividade antimicrobiana, emulsificado- ra de substâncias e de barreira protetora. e Excreção. As glândulas écrinas secretam água, eletrólitos, HCO, uréia, metais pesados etc., à semelhança do rim. * Metabolização. A pele também sintetiza hormônios, dentre eles a testosterona e diidrotestosterona, que têm um papel muito importante na alopecia androgenética, acne e hirsutis- mo. Tem também uma ação decisiva no fabrico é na metabo- lização da vitamina D, Quadro de Abreviaturas CORP Peptídeo Relacionado ao Gene da Calcitonina EGF Fator de Crescimento Epidérmico FGFb Fator de Crescimento Fibroblástico básico GM-CSF Fator Estimulador de Colônia de Granulócito-Macrófago IL Interlcucina KGF Fator de Crescimento Ceratinocítico LTB4 Leucotrieno B4 M-CSF Fator Estimulador de Colônia de Macrófago MSH Hormônio Estimulante de Melanócitos PDGF Fator de Crescimento Derivado de Plaquetas PGE: Prostaglandina E? TGF Fator Transformador do Crescimento TNF Fator de Necrose Tumoral SCF Fator de Células-Tronco VEGF Fator de Crescimento Endotelial Vascular O A Pele — Estrutura, Fisiologia e Embriologia 13 Microscopia Eletrônica Imagens fornecidas gentilmente pela Dra. Mirian N. Sotto, da Faculdade de Medicina da USP. Fig. 1.11 Pele — junção dermoepidérmica. N — núcleo de melanócito, Fig. 1.13 Pele — camada espinhosa da epiderme. D — desmossomos; M — melanossomos; T — tonofilamentos citoplasmáticos de T— tonofilamentos citoplasmáticos agrupados formando tonofibrilas: ceratinócito basal; lâmina basal (setas), F — fibrilas colágenas; H — — M — mitocôndria;* — espaço intercelular. (28,500 x.) hemidesmossomo. (14.700 x.) Fig. 1.12 Pele — camada espinhosa da epiderme. N — núcleos de cera Fig. 1.14 Pele — célula de Langerhans. N — núcleo: C — corpúsculos tinócitos da espinhosa; desmossomos (setas); T — tonofilamentos cito- de Langerhans ou em “raquete”; G — complexo de Golgi; M — mito- plasmáticos;* — espaço intercelular. (5.700 x.) côndrias. (28.500 x.) Noções Básicas de Imunologia Cutânea Capítulo 2 INTRODUÇÃO A pele é um órgão importante de expressão imunológica, e os dermatologistas têm sido pioneiros em muitos aspectos da imunologia. Assim, Jadassohn (1891) introduziu a noção de hi- persensibilidade a um metal (mercúrio), inclusive com a reali- zação dos testes de contato, desenvolvidos mais tarde por Bruno Bloch; Jadassohn antecipou-se em 10 anos ao conceito de aler- gia de Pirquet (1906). Foi ainda Jadassohn quem mostrou o mecanismo imunológico da estrutura tuberculóide, criando a lei das proporções invertidas. O estudo das vasculites foi iniciado por Gougerot (doença trissintomática); quanto à auto-agressão, salientamos os trabalhos pioneiros de Withfield (1929) sobre a auto-eczematização. O sistema imunológico é composto por órgãos, tecidos e cé- lulas especializadas ou não que visam defender a integridade do organismo, reconhecendo e eliminando qualquer agente estranho a ele; discrimina o que lhe é intrinsecamente próprio do que não lhe é próprio, isto é, reconhece que determinada substância não faz parte do organismo e, portanto, deve rejeitá-la. É constituído por órgãos linfóides primários (medula óssea e timo) e órgãos linfóides secundários (baço, linfonodos e tecido linfóide associado a mucosas — MALT — como a do TGI e do trato aéreo respiratório). Conta ainda com a participação de ele- mentos inespecíficos como barreiras físicas (pele e mucosas), fatores solúveis (citocinas, quimiocinas e enzimas) e células (macrófagos, mastócitos, basófilos etc.) que atuam conferindo a imunidade inata, não-específica. A imunidade adquirida requer a participação de células es- pecializadas, os linfócitos (a maioria), capazes de reconhecer com grande especificidade uma infinidade de antígenos estranhos, graças a moléculas na superfície celular resultantes do rearranjo de genes (receptores nos linfócitos T e imunoglobulinas nos lin- fócitos B). O objetivo desse mecanismo de defesa é evitar que o orga- nismo seja agredido por substâncias vivas (microorganismos, vírus, protozoários etc.) ou inanimadas (proteínas estranhas e Mario Geller, David R. Azulay e Rubem D. Azulay outros). No rechaço a essas substâncias, pode processar-se, por mecanismos de hipersensibilidade, de modo paradoxal, agressão ao próprio ser; é um exagero da defesa que passa a fazer parte da etiopatogenia de várias doenças (auto-imunidade e reações alér- gicas). A um desses efeitos nocivos do sistema imunológico cabe o termo alergia, enquanto, ao efeito benéfico de defesa, impe- dindo a agressão, reserva-se o termo imunidade. Por outro lado, quando o sistema imunológico é hipoativo, o organismo é sus- cetível a infecções fregiientes (deficiência imunológica). Há outros aspectos a considerar no sistema imunológico: homeos- tasia e vigilância imunológicas. A homeostasia imunológica visa à remoção, por mecanismo imunológico, de células alteradas por qualquer mecanismo; quando essa atividade passa a ser hipera- tiva, surge a auto-agressão, processo cada vez mais importante em patologia. A vigilância imunológica visa a reconhecer e eli- minar células mutantes (espontâneas ou induzidas por vírus ou agentes físicos e químicos); quando esse mecanismo é hipoativo, surgem as neoplasias. Há também a considerar a apoptose, mor- te celular não-inflamatória, de programação genética individu- al, que faz parte do processo natural do envelhecimento biológi- co. No estudo do sistema imunológico, devemos estudar as cau- sas (imunógenos e/ou antígenos), as células reatoras (linfócitos, macrófagos, células de Langerhans, e outras), a especificidade do processo e a memória imunológica celular. Várias substânci- as produzidas por essas células imunorreatoras participam de processos imunoinflamatórios complexos: citocinas, quimioci- nas, moléculas de adesão, imunoglobulinas, complemento, en- tre tantas outras. Para melhor desempenho de suas funções, o sistema imune funciona integrado aos sistemas do complemen- to, das cininas e da coagulação. Imunógenos são substâncias dotadas de imunogenicidade, ou seja, capazes de provocar uma resposta imunoespecífica, quer pela indução e elaboração de anticorpos circulantes específicos, quer pela atividade do sistema imune mediado por células com a conversão de linfócitos T em linfócitos especificamente sensi- bilizados. Os imunógenos correspondem aos antígenos comple- 16 Dermatologia tos, isto é, per se, estimulam a formação e produção de anticor- pos. Os haptenos (antígenos incompletos) possuem somente antigenicidade e necessitam unir-se a uma proteína para induzi- rem a formação de anticorpos, ou seja, necessitam do acoplamen- to com suporte protéico para adquirirem imunogenicidade. Está implícito, pois, que a imunogenia requer estrutura molecular complexa, ou seja, a molécula protéica; as substâncias de molé- culas menos complexas (lipídios, glicídeos, substâncias simples ou compostas) não têm capacidade antigênica, a não ser que se acoplem a uma estrutura protéica, e, nesse caso, a resposta espe- cífica ocorre em relação a essas moléculas pouco complexas, e não à proteína que serviu de suporte. As glicoproteínas são os antígenos mais prevalentes e importantes. Epítopo ou determinante antigênico é a parte do antígeno à qual o anticorpo se liga; portanto, os anticorpos são específicos para os epítopos. Os denominados anticorpos antiidiotípicos são modalidades de auto-anticorpos voltadas contra as segiiências de aminoácidos presentes nas porções variáveis das imunoglobuli- nas. Entende-se por superantígenos um grupo de proteínas (exo- toxinas) produzidas por várias bactérias (Streptococcus, Sta- phylococcus aureus, Mycoplasma etc.) e vírus. Ao contrário dos antígenos normais, não necessitam obrigatoriamente da presen- ça de células apresentadoras de antígenos para serem capazes de ativar células T e são capazes de ativá-las de maneira inespecífi- ca, ou seja, mais de 20% de todas as células T, em vez de 1/ 100.000 células, como nas respostas antigênicas específicas. Superantígenos são potentes ativadores policlonais de células T (ver Quadro 2.1). Para que o organismo se sensibilize imunologicamente, é necessário um contato prévio com o antígeno e um período mí- nimo de latência de dias (para antígenos com grande capacidade de sensibilização), a partir do qual haverá, a cada nova exposi- ção, uma resposta imunológica; isto é devido à memória celular, que permite o seu reconhecimento, e é antígeno-específico. Quadro 2.1 Ativação das Células T na Presença de Antígeno (Peptídeo) e de Superantígeno Receptor ass car cab Antígeno Superantígeno Classe II MHC Célula Célula apresentadora apresentadora | Ag Ag O antígeno se liga na depressão antigênica da molécula de MHC II e ativa célu- las T através dos seus receptores. Já o superantígeno se liga com a molécula de MHC II por fora da depressão antigênica com determinadas cadeias f do recep- tor de célula T, levando à ativação policlonal dos linfócitos T. A explicação recente desse reconhecimento é devida à teoria clonal de Burnet, pois, desde o nascimento, um ser tem linfóci- tos com receptores para cada espécie de antígeno. Este, ao pene- trar no organismo, liga-se à superfície da célula de Langerhans ou do macrófago (células apresentadoras de antígenos), onde é interiorizado, processado, exteriorizado e apresentado como peptídeo antigênico, o que possibilita a ativação do linfócito T mais adequado ao mesmo. Essa ativação se dá no local, em me- nor escala, e através da migração, via linfática, das células de Langerhans, macrófagos e linfócitos T já sensibilizados para o linfonodo regional, e se processa de duas maneiras: e proliferação, que compreende as divisões celulares sucessi- vas no sentido da produção de células com as mesmas carac- terísticas imunológicas; e diferenciação, isto é, o citoplasma, que era mínimo no linfó- cito inativo, torna-se exuberante, graças à elaboração de um abundante retículo endoplasmático, ao lado de certas altera- ções nucleares (imunoblasto). Essa diferenciação correspon- de à transformação blástica e ocorre na zona paracortical do linfonodo. Alguns desses linfócitos tornam-se linfócitos efetores e migram, via corrente sangiiínea, para os locais que contêm o antígeno. Pequenos linfócitos permanecem nos lin- fonodos, tornando-se células de memória. Os linfócitos T sensibilizados, ou seja, aqueles que sofreram transformações blásticas, elaboram linfocinas que induzem a resposta infla- matória. É digno de nota que, no processo inflamatório, ape- nas 2% dos linfócitos estão sensibilizados; as demais células foram recrutadas através das linfocinas e não interagem de maneira específica com o antígeno. Há também a participa- ção nas dermatoses imunoinflamatórias, como a dermatite atópica, de células epidérmicas dendríticas inflamatórias (IDEC), de linhagem monocitária, que promovem e perpetu- am esse quadro inflamatório. O mecanismo descrito anteriormente corresponde ao da hi- persensibilidade retardada de que temos, na prática dermatoló- gica, como melhor exemplo, por sua fregiiência, o eczema de contato (dermatite alérgica de contato). Segundo os conceitos atuais, a resposta imune é mediada pelos linfócitos T (imunidade celular) e B (imunidade humoral), que in- teragem em diversos níveis, visando à proteção do organismo. Eles reconhecem os antígenos através de receptores específi- cos em suas membranas que atuam em sincronia com os antíge- nos de histocompatibilidade. IMUNIDADE CELULAR É representada pelas seguintes células: Linfócitos T São timodependentes; esses linfócitos originam-se na medu- la óssea e migram para o timo. Neste, ocorre a elaboração de substâncias hormônio-símiles (timosina, timopoetina etc.) que, agindo sobre os mesmos, os tornam imunocompetentes. Existem várias subpopulações de linfócitos atuando de ma- neira distinta e reconhecidas por suas diferentes moléculas de 18 Dermatologia sea, penetra no sangue (monócito) e deste aos tecidos; sua qua- lidade bem conhecida é a fagocitose, através da qual apreende e engolfa as substâncias animadas ou inanimadas e as destrói; daí o seu relevante papel na inflamação, quando ativado por deter- minadas linfocinas; deve-se enfatizar que, num infiltrado, resul- tante da imunidade mediada por células, há 95% de macrófa- gos para 5% de linfócitos T. Essa fagocitose pode ser potencializada pela opsonização, isto é, anticorpos específicos (opsoninas/bacteriotropinas) que recobrem o antígeno, facilitan- do, assim, a fagocitose. É o macrófago que apresenta o antíge- no ao linfócito T de modo a provocar e/ou aumentar a resposta imune celular. Sabe-se que o linfócito T por si só interage inex- pressivamente com antígeno livre, mas é estimulado e exaltado na sua função quando esse antígeno é elaborado e apresentado pelo macrófago (célula APC, apresentadora de antígeno). O macrófago também colabora com o linfócito T helper na esti- mulação da produção de anticorpos pelo linfócito B (IgG, IgA, IgM e IgE). Para a realização dessas funções imunológicas, o macrófago expõe peptídeos antigênicos aos receptores de lin- fócitos T (TCR), na presença de moléculas MHC II (major histocompatibility complex IN). Experiências in vitro evidenci- am que efeitos idênticos são obtidos apenas com extratos de macrófagos ou líquidos sobrenadantes de cultura de macrófa- gos. Sob ativação extrema e peculiar, os macrófagos podem fundir-se e constituir as células gigantes multinucleadas, tipi- camente presentes nos granulomas. A maioria dos monócitos é CD14+. Os macrófagos também sintetizam leucotrienos, pros- taglandinas, fator de ativação plaquetária e lipoxinas, contribu- indo, assim, para o estabelecimento de diversos processos imunoinflamatórios. As interações ligantes são primordiais nos processos de apresentação antigênica. As células fagocíticas também estão ativamente envolvidas na patogênese das várias manifestações alérgicas cutâneas. Células de Langerhans Têm origem na medula óssea e apresentam características imunológicas semelhantes às dos macrófagos, só que de locali- zação predominantemente epidérmica (células APC residentes). Apresentam, como os macrófagos, receptores de superfície para a fração Fc da imunoglobulina G, FCeRII, do terceiro compo- nente do complemento (C3) e Ia antígenos (MHC II) que são geneticamente determinados por genes localizados próximo ao locus D, que, no rato, ocorre numa região do cromossomo cha- mado I. As células de Langerhans têm papel importante em vá- rios processos patológicos (micose fungóide, histiocitoses, ecze- ma de contato etc.). São vitais para a imunovigilância cutânea; participam também dos processos imunopatológicos presentes na dermatite atópica e na dermatite alérgica de contato. IMUNIDADE HUMORAL A célula fundamental é o linfócito B, que, nas aves, se origi- na na bursa de Fabricius (daí B) e, no homem, na medula óssea e, provavelmente, nas amígdalas (anel de Waldeyer), placas de Peyer e apêndice. Esses linfócitos B da medula migram para os órgãos linfóides periféricos (gânglios linfáticos), onde comple- tam a sua maturação. Normalmente, esses linfócitos B são en- contrados no sangue, nas áreas cortical e medular dos gânglios linfáticos, e nos folículos esplênicos. Os linfócitos B, células imunocompetentes, mesmo antes da estimulação antigênica específica, são facilmente reconhecidos por apresentarem as imunoglobulinas D e M em sua superfície. Após exposição ao antígeno, um certo número de células B so- fre proliferação clonal e diferenciação (transformação blástica), adquirindo, então, as características do plasmócito, que tem retículo plasmático muito desenvolvido ao lado de numerosos ribossomos, produzindo, então, as imunoglobulinas específicas, voltadas contra o antígeno estimulador, que pode ser um alérgeno. Para que haja produção máxima de anticorpos, é necessária interação entre o macrófago, célula de Langerhans e linfócitos T e B, e isso ocorre através da subpopulação T helper, enquanto a subpopulação T supressor tende a limitar tal processo. A perda do equilíbrio entre essas subpopulações é a origem de inúmeras doenças. Alguns antígenos, no entanto, são capazes de induzir a transformação blástica dos linfócitos T sem a necessidade dessa interação. São os denominados superantígenos, e um exemplo típico é o das exotoxinas do Staphylococcus aureus, capazes de promover e estimular a síntese de IgE específica, contribuindo, assim, para a imunopatogênese da dermatite atópica. Os anticorpos produzidos pelos linfócitos B são as imunoglo- bulinas, que se agrupam em cinco classes: IgG (IgG1, 2, 3, 4), IgA (IgAI, 2), IgM, IgD e IgE. As imunoglobulinas são constituídas de quatro grupos peptí- dicos, sendo dois de cadeias pesadas (peso molecular 50.000) e dois de cadeias leves (peso molecular 25.000); as cadeias pesa- das diferem de acordo com a classe das imunoglobulinas (y, a, E À, £); ligadas entre si por pontes de bissulfeto, possuem dois fragmentos Fab (ab = antigen binding) e um Fe, que permite fixação à superfície de certas células (receptores celulares). As cadeias leves podem ser do tipo x (kappa) e do tipo À (lambda). A IgG está presente predominantemente no plasma e transudato intersticial, e representa cerca de 75% das imunoglo- bulinas do soro e a maioria dos anticorpos, sobretudo antibacte- rianos; a IgG é a de menor peso molecular (150.000) e, por isso mesmo, capaz de atravessar a placenta, sendo, portanto, respon- sável pela imunidade transferida passivamente ao recém-nato; é capaz de fixar o complemento, e sua concentração no soro é de 5-12 mg/dl. Inúmeros anticorpos pertencem às IgG: antivirais, antitoxinas, hemoaglutininas e antimicrobianos. Há quatro sub- classes de IgG; as IgG1 e IgG3 fixam-se aos macrófagos e exer- cem ação citotóxica. As IgA são os anticorpos que predominam na saliva, secre- ções intestinal, broncopulmonar, genitourinária, lágrima e colostro; existem ainda em pequena quantidade no soro (2 mg/ dl); têm função de defesa nas superfícies das mucosas, possuin- do peso molecular maior que a IgG (400.000); não fixam com- plemento pela via clássica, mas podem fixá-lo pela via alterna- da; não atravessam a barreira placentária e nem se fixam em macrófagos e mastócitos. A IgA existe nas apresentações monomérica e polimérica. A forma polimérica mais prevalente é a dimérica, que contém a cadeia J unindo as duas subunidades. A forma dimérica contém um componente secretório encontra- do nas secreções exócrinas, o que indica a sua origem mucosa, diferentemente da monomérica. As IgM são anticorpos que surgem sobretudo no início da resposta imune, representam as crioglobulinas, o fator reumatói- de, inúmeros anticorpos antibactérias e do sistema ABO; são os anticorpos de maior peso molecular (900.000) e encontram-se no sangue na concentração de 1,2 mg/ml; fixam o complemen- to, porém não se fixam em macrófagos e mastócitos. As IgD são infreqiientes e foram descobertas no mieloma múltiplo específico; é discutível se representam alguns anticor- pos antiinsulina, antipenicilina, e auto-anticorpos; têm peso molecular 185.000, e sua concentração sérica é de 0,03 mg/ml. É a imunoglobulina mais encontrada na superfície do linfócito B. As IgE são os anticorpos reagínicos que se encontram na ato- pia e tomam parte na anafilaxia; provocam reações alérgicas imediatas (onde ocorre a clássica liberação de histamina), fixam- se aos mastócitos e basófilos (receptores de alta afinidade, FCeRI), células dendríticas, linfócitos B e eosinófilos (recepto- res de baixa afinidade, FCeRII); têm peso molecular de 200.000 e sua concentração sérica é insignificante (0,00004 mg/ml); es- tão acentuadamente elevadas nas parasitoses por helmintos, po- rém não por protozoários, e também aumentadas nos processos alérgicos. A hiperimunoglobulinemia E é típica da dermatite atópica, onde fregiientemente está associada à hipereosinofilia sangiiínea, refletindo um excesso de atividade Th2, através da maior liberação de IL-4, IL-5 e IL-13. As células B podem mudar a sua programação de síntese das imunoglobulinas (isorype switching). IL-10 induz switching para IgG1 e IgG3; IL-4e IL-13 promovem switching para IgE; TGF- B causa switching para IgA; IFN-y promove switching para IgG2. Null Cells (Non-B, Non-T Cells) São leucócitos com propriedades tanto de linfócitos quanto de células monocíticas. Representam a terceira linhagem de cé- lulas linfóides. Originam-se da medula óssea sob a influência de IL-2 e IL-15. A maioria contém receptores para Fc IgG. Nessa subpopulação, temos as NK e K. Células NK (natural killer) — são linfócitos com grânulos cito- plasmáticos grandes e que têm capacidade de destruir células tumorais ou infectadas por vírus, sem estarem previamente sensibilizados com anticorpo. Células K (killer) — são células tais como TC, NK ou monóci- tos cuja função é destruir células previamente sensibilizadas por anticorpo, ou seja, ligam-se a este e destroem o alvo. As células killer participam ativamente da citotoxicidade mediada por células dependente dos anticorpos (ADCC), presente nas ações de defesa imunológica antivirais e antitumorais. Citocinas e Quimiocinas As citocinas e as quimiocinas são proteínas associadas com o crescimento, a diferenciação e as funções ativadoras, responsá- veis pelas respostas imunes, o tráfego e a organização celular presentes nas estruturas que integram o sistema imunológico. Noções Básicas de Imunologia Cutânea 19 As citocinas são pequenas moléculas, de natureza protéica, como já mencionado, secretadas por células tais como linfóci- tos, macrófagos, queratócitos, plaquetas etc. As linfocinas são as citocinas produzidas pelos linfócitos. Elas regulam a ativida- de imunológica de outras células que tenham receptores especí- ficos na membrana citoplasmática, embora, ocasionalmente, pos- sam ser compartilhadas por mais de um tipo de citocina. Duran- te a inflamação, ocorre uma interação complexa entre elas, cu- jos efeitos variam em função da natureza, intensidade e duração do processo. Fregientemente, as citocinas têm ações sobrepos- tas e podem agir sinérgica ou antagonicamente. Uma determi- nada citocina pode atuar em diferentes células. Exemplos de ci- tocinas: interleucinas — existem 23 tipos (IL-1 a IL-23); inter- ferons (a, B e Y); fatores estimulantes de colonização para gra- nulócitos, macrófagos, plaquetas etc.; fatores de crescimento (epidérmico, fibroblástico, plaquetário etc.); e fatores de necro- se tumoral (TNF-a e B). As citocinas são produzidas em respos- ta a uma agressão imune e a sua atuação-resposta poderá ser ci- totóxica, humoral, celular ou alérgica. Poderá haver uma resposta em cascata sinergística. As funções das citocinas podem variar de acordo com as suas fontes celulares, alvos direcionados e fa- ses específicas das respostas imunes presentes. Consegientemen- te, as mesmas citocinas podem apresentar tanto potenciais pró- inflamatórios quanto antiinflamatórios. As citocinas podem ter origem mononuclear fagocítica ou linfocítica T; ação imunoló- gica citotóxica (antiviral e antitumoral), humoral, celular e alér- gica. Podem, entretanto, ser imunossupressoras. As quimiocinas são grupadas em famílias. São moléculas de pequeno peso molecular (8-15 kD). Induzem a quimiotaxia de neutrófilos, monócitos, linfócitos, eosinófilos, fibroblastos e queratócitos. Há 47 quimiocinas e 18 receptores de quimiocinas já identificados. Elas participam ativamente dos quadros imunoinflamatórios e dos processos de cicatrização subsegiien- tes. São igualmente importantes na manutenção da homeostasia imune e da imunovigilância. Elas contribuem significativamen- te para o estabelecimento da inflamação alérgica. Vai muito além do objetivo deste compêndio discutir as cito- cinas individualmente. Para melhor situar o leitor e demonstrar a magnitude dessas interações, será abordado especificamente o TNF-a, atualmente muito discutido em Dermatologia. O TNF-« é sintetizado a partir de queratócitos, fagócitos epi- dérmicos, mastócitos, células NK e linfócitos T ativados nos sí- tios de inflamação. Trata-se de polipeptídeo de 17 kD, compos- to por 150 aminoácidos, que é capaz de induzir a necrose hemor- rágica em células tumorais, daí a sua designação. Entre as suas ações, temos: sobre fibroblastos (induzindo a proliferação e pro- dução de prostaglandinas e colagenases); sobre macrófagos (am- plia a produção de citocinas e a citotoxicidade antibacteriana); sobre células endoteliais (ativando a coagulação e o plasmino- gênio): sobre granulócitos, neutrófilos e eosinófilos (aumentan- do a quimiotaxia, aderência, fagocitose). Nos vasos, sua impor- tância é crítica: estimula a expressão das moléculas de adesão ICAM-1, VCAM-1 e E-selectina na superfície de células endo- teliais, fator fundamental para atrair leucócitos circulantes para os sítios em que está ocorrendo a inflamação. Os lipopolissacarídeos (LPS) bacterianos são os principais indutores de TNF através de TLR2 e TLR4 (toll-like receptors),