



Tese de Doutorado
Caracterização do material magnético em
antenas de formigas Pachycondyla
marginata: um possível sistema
magnetosensor
Jandira Ferreira de Oliveira
Centro Brasileiro de Pesquisas Físicas
Rio de Janeiro, março de 2010






















































































Estude fácil! Tem muito documento disponível na Docsity

Ganhe pontos ajudando outros esrudantes ou compre um plano Premium

Prepare-se para as provas
Estude fácil! Tem muito documento disponível na Docsity
Prepare-se para as provas com trabalhos de outros alunos como você, aqui na Docsity
Encontra documentos específicos para os exames da tua universidade
Prepare-se com as videoaulas e exercícios resolvidos criados a partir da grade da sua Universidade
Responda perguntas de provas passadas e avalie sua preparação.
Ganhe pontos para baixar
Ganhe pontos ajudando outros esrudantes ou compre um plano Premium
Artigo mostrando magnetismo em formigas
Tipologia: Manuais, Projetos, Pesquisas
1 / 138

Esta página não é visível na pré-visualização
Não perca as partes importantes!
Agradecimentos :
i
Resumo. ................................................................................................................................... iii Abstract .................................................................................................................................. v Lista de Figuras ................................................................................................................... vii Lista de Tabelas ..................................................................................................................... xv Introdução ............................................................................................................................... 1
Capitulo I - Órgãos sensoriais na antena dos insetos e seu possível uso como magnetoreceptores .................................................................................................................. 3
Capitulo II - Revisão
Capitulo III – Material e métodos
Capitulo IV – Resultados e discussão
iv
Análises posteriores, utilizando microscopia eletrônica de transmissão para imagem e obtenção de padrões de difração de elétrons, combinada com análise elemental, revelaram a presença de grãos cristalinos ultrafinos (20-100 nm) de magnetita/maghemita (Fe 3 O 4 , γ- Fe 2 O 3 ), hematita (α-Fe 2 O 3 ), goetita (α-FeOOH) além de (alumo)silicatos e compostos de Fe/Ti/O em distintas partes da antena, que são, as junções entre o terceiro segmento e o pedicelo, pedicelo e escapo e escapo e cabeça, respectivamente. A presença de (alumo)silicatos e compostos de Fe/Ti/O sugerem que a maior parte, se não toda, dos minerais encontrados no tecido sejam partículas incorporadas do solo e não biomineralizadas pelas formigas. No entanto, já que as partículas foram observadas dentro de tecidos elas não representam contaminação. A quantidade de material magnético associado ao órgão de Johnston e outras junções parece ser suficiente para produzir um output mecanosensorial modulado pelo campo magnético, que pode portanto ser a base do sensor magnético desta formiga migratória.
v
Abstract
The influence of the geomagnetic field on the behaviour of living beings has been studied in phenomena like migration, homing, etc. Magnetite crystals were observed in magnetotactic bacteria at about 30 years ago. It was the first physical evidence of a passive mechanism, magnetotaxis, through the alignment of magnetotactic bacteria with a magnetic field (Blakemore 1975). Behavioural studies have shown that the geomagnetic field is also an environmental information used for orientation and navigation by a great number of animals. Magnetoreception is a complex mechanism, yet poorly understood, that involves the detection of the geomagnetic field and a subsequent transduction of the signal to the nervous system. The information is transferred to a specific brain area and used for different proposals. For magnetoreception, two hypotheses are currently discussed, one proposing a chemical compass based on a radical pair mechanism, the other postulating processes involving magnetite particles, named ferromagnetic hypothesis. Migration of the Pachycondyla marginata ant is significantly oriented at 13o^ with respect to the geomagnetic north–south axis. On the basis of previous magnetic measurements of individual parts of the body (antennae, head, thorax and abdomen), the antennae were suggested to host a magnetoreceptor. Electron Paramagnetic Resonance (EPR) measurements on individuals, parts or groups of insects at room temperature and a temperature dependence for the characterization of the magnetic material in the samples were done. Magnetic extraction of the samples measured by EPR identified particles containing Al, Si, Fe, O and Ti. However, there was no indication for biomineralized material in any of the body parts analysed. The only possible way to find out biomineralized material is through identification of such particles within the tissue. Light microscopy was used to identify Fe 3+/Fe 2+^ sites in antennae tissue through Prussian/Turnbull’s blue staining methods. Further analysis using transmission electron microscopy imaging and diffraction, combined with elemental analysis, revealed the presence of ultra-fine-grained crystals (20–100 nm) of magnetite/maghemite (Fe 3 O 4 , γ-Fe 2 O 3 ), hematite (α-Fe 2 O 3 ), goethite (α-FeOOH) besides (alumo)silicates and Fe/Ti/O compounds in different parts of the antennae, that is, in the joints between third segment/pedicel, pedicel/scape and scape/head, respectively. The presence of (alumo)silicates and Fe/Ti/O compounds suggests that most, if not all, of the minerals in the tissue are incorporated soil particles rather than biomineralized by the ants. However, as the particles were observed within the tissue, they do
vii
Lista de figuras:
Capítulo I
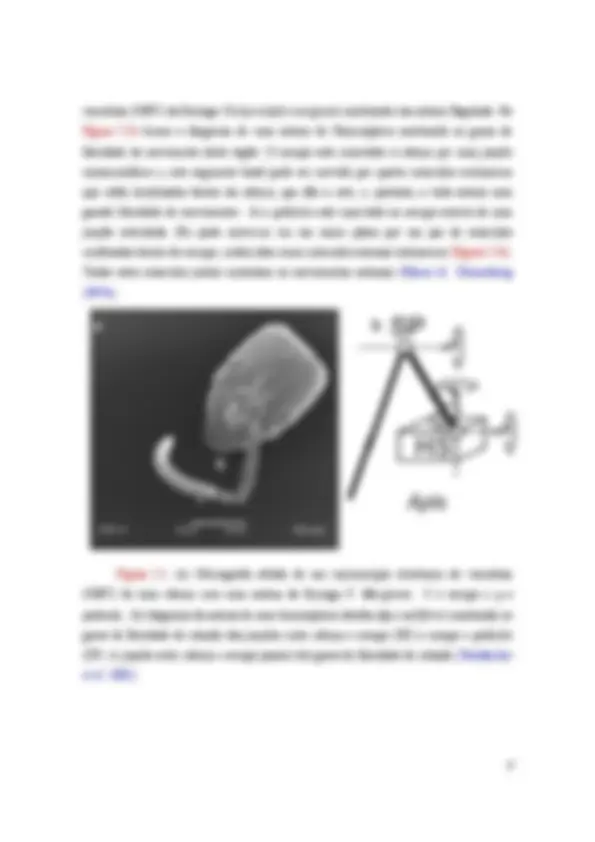
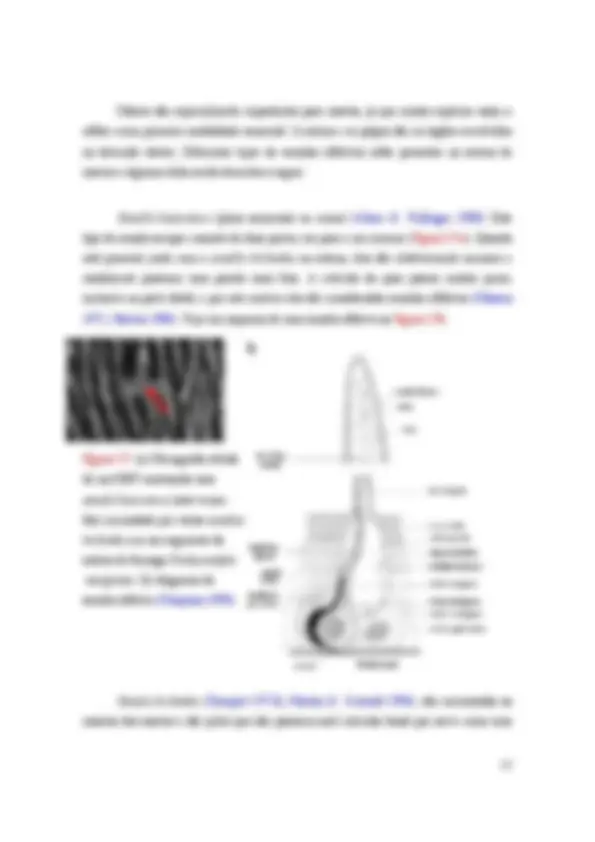
Figura I.1: (a) Micrografia obtida de um microscópio eletrônico de varredura (MEV) de uma cabeça com uma antena da formiga P. Marginata. S é escapo e p o pedicelo. (b) diagrama da antena de uma himenóptera (abelha Apis mellifera ) mostrando os graus de liberdade de rotação das junções entre cabeça e escapo (HS) e escapo e pedicelo (SP). A junção entre cabeça e escapo possui três graus de liberdade de rotação (Staudacher et al. 2005).
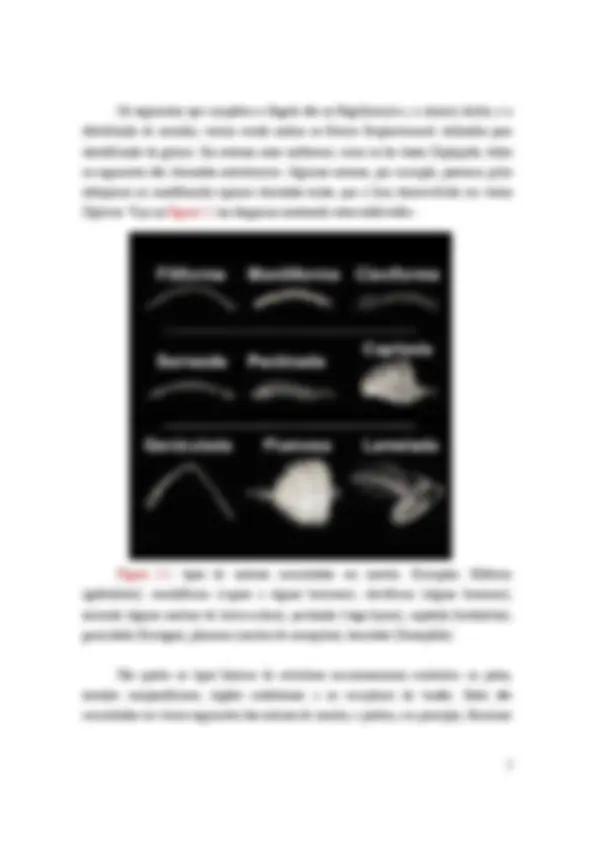
Figura I..2: tipos de antenas encontradas em insetos. Exemplos: filiforme (gafanhotos), moniliforme (cupins e alguns besouros), claviforme (alguns besouros), serreada (alguns machos de louva-a-deus), pectinada (vaga-lumes), capitada (borboletas), geniculada (formigas), plumosa (machos de mosquitos), lamelada (Drosophila).
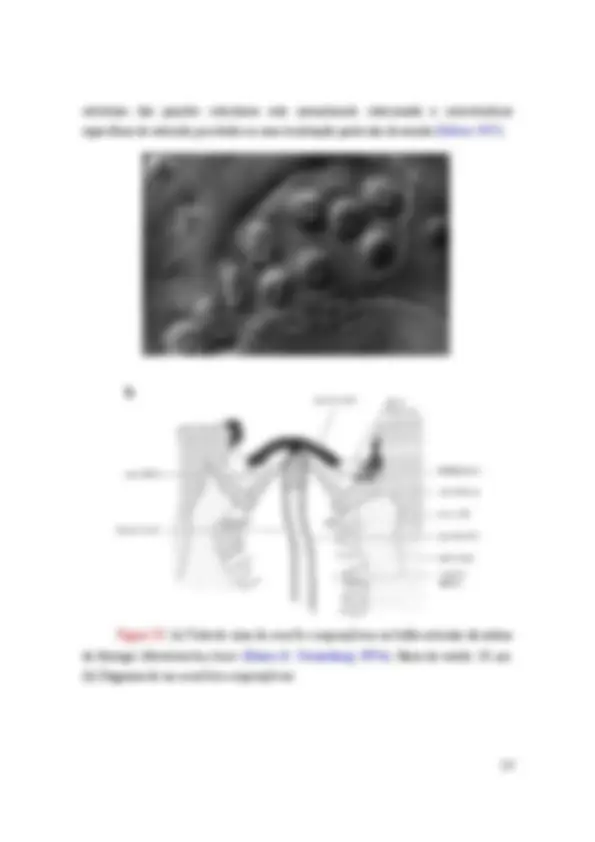
Figura I.3: órgão de Johnston em Formica rufa. A parede dorsal do pedicelo é removida para mostrar o arranjo dos escolopídeos dentro do pedicelo (Vowles 1954).
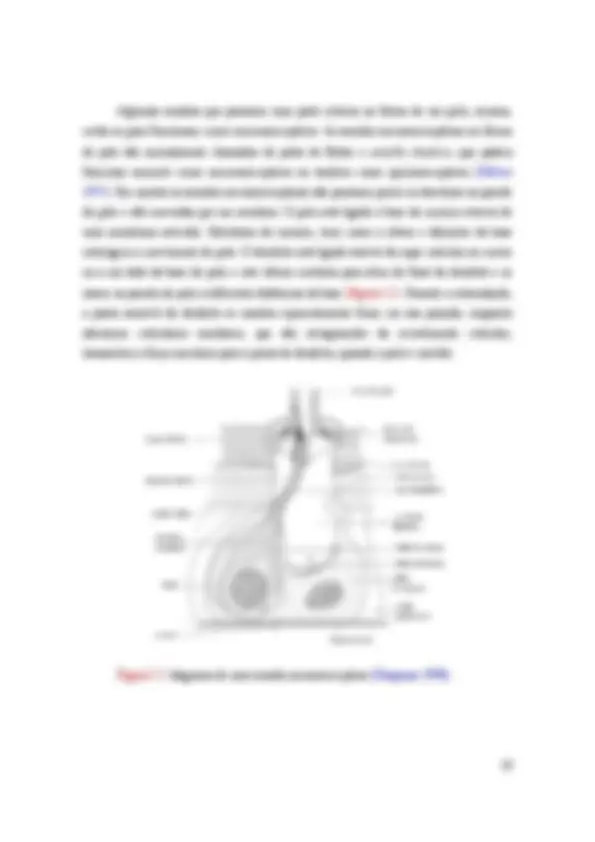
Figura I.4: diagrama, representando a maior parte do segmento antennal de uma abelha operária ( Apis mellifera ) em uma seção longitudinal e em perspectiva e uma pequena seção do terceiro segmento (longitudinal, transversal e em perspectiva) mostrando o órgão de Johnston que consiste de dois grandes grupos de células sensoriais (SC) cujas fibras nervosas (NP) correm para dois nervos antenais (N) e cujas fibras sensoras (SF) estão ligadas a botões (K) na membrana articular. Dois botões são cortados no sentido de seus comprimentos, mostrando a cavidade em forma de cone que se abre para o exterior, enquanto os outros estão inseridos na membrana articular. Tr, traqueia, Hyp, hipoderme, Ch, quitina dura, Ch1, faixas de quitina macias e flexíveis que mantém o segundo e terceiro segmentos firmemente ligados (McIndoo 1922).
Figura I.5: diagrama de uma sensila mecanoreceptora (Chapman 1998).
Figura I.6: Pelos de Boehm no bulbo articular da formiga Odontomachus bauri. Barra de escala: 10 μm. (Ehmer & Gronenberg 1997).
viii
Figura I.7: (a) Vista da parte distal do escapo e início do pedicelo obtido por MEV. Setas mostram sensilla chaetica. (b) diagrama da sensila gustativa (Chapman 1998).
Figura I.8: (a) Vista de cima de sensilla campaniforme no bulbo articular da antena de formiga Odontomachus bauri (Ehmer & Gronenberg 1997). Barra de escala: 10 μm. (b) Diagrama de um sensillum campaniforme.
Figura I.9: (a) Micrografia obtida de um MEV mostrando uma sensila basiconica (seta verme- lha) circundado por várias sensilas trichodea em um segmento da antena de formiga Pachycondyla marginata. (b) diagrama da sensila olfativa (Chapman 1998).
Figura I.10: sensilla trichodea distribuídas ao longo do 3º segmento da antena de formiga Pachycondyla marginata. Na parte mais distal deste segmento algumas sensilla chaetica podem ser observadas.
Figura I.11: MEV do flagelômero antennal 11 de um macho africanizado ( Apis mellifera ) mostrando detalhes das sensilla campaniforme (seta pequena), sensilla ampulacea (seta media), sensilla coeloconica (seta longa). SP é um sensillum placodea. (Stort & Moraes- Alves 1999).
Figura I.12: Representação esquemática de uma sensila sem poro com encaixe não-flexível capaz de perceber variação térmica e de umidade (Altner & Loftus 1985).
Capítulo II
Figura II.2.1: (a) Diagrama ternário TiO 2 – FeO – Fe 2 O 3 mostrando as linhas de soluções sólidas das titanomagnetitas e titanohematitas e o campo das titanomaghemitas. Durante a oxidação de alta e baixa temperatura das titanomagnetitas, a composição volumétrica segue as linhas tracejadas horizontais. (b) Linhas de mesma temperatura de Curie para titanomagnetitas sintéticas (Dunlop & Özdemir 1997).
Figura II.2.2: Célula unitária do espinélio mostrando a localização dos íons em dois dos oito octantes. Os íons de ferro dos sítios A e B da magnetita dão origem a uma subestrutura
x
flagelo e as setas vermelhas dos longitudinais. Cortes oblíquos também são obtidos dependendo do alinhamento do flagelo em relação ao escapo.
Figura III.8.1: O microscópio eletrônico como uma simples câmera de difração de elétrons.
Capítulo IV
Figura IV.1.1: Espectros de RFM (a) de solo: comparação de diferentes amostras de 1g posteriormente dividida em amostras de 0,500g, 0,250g, 0,125g e 0,100g e extrato magnético 0,01 g (linha tracejada) (pmw 2mW). (b) 5 amostras de solo, m = 0,0016g (pmw 4mW). Intensidade em (b) dividida por 14,1.
Figura IV.1.2: Diagrama dos parâmetros obtidos dos espectros de RFM, segundo Weiss et al. 2004 das amostras de terra medidas a temperatura ambiente.
Figura IV.1.3: variação térmica dos espectros (a) T3 e (b) T4.
Figura IV.1.4: Variação térmica dos parâmetros dos espectros de RFM. (a) campo de ressonância, Hr (b) largura de linha, ΔHpp (c) fator de assimetria, Aw, seta aponta para temperatura de transição (T = 210 K) (d) 2ª integral ou área do espectro de absorção, S.
Figura IV.1.5: Variação da largura de linha com o inverso da temperatura para a componente CA1 da amostra T3 e da amostra T4. Linha cheia ajuste da eq. I.3.9, com KV= (2,7±0,2)10 - (^14) erg e pré-fator = 725±33 Oe e KV= (7,6±0,2)10-14 (^) erg e pré-fator = 1370±7 Oe, para CA
de T3 e T4 respectivamente.
Figura IV.1.6: Espectros a temperatura ambiente de 3 formigas P. marginata “mortas”.
Figura IV.1.7: Distribuição dos parâmetros de RFM: (a) Hr, (b) ΔHpp , (c) S para 4 mW e (d) 10 mW.
Figura IV.1.8: Espectros a temperatura ambiente de formigas P. marginata “mortas” (a) e “vivas” (b).
xi
Figura IV.1.9: variação térmica dos espectros de (a) 88 escapos, (b) 92 flagelos. pmw: 20mW, ganho: 110^5 e 4 varreduras. (c) extratos magnéticos de antena. pmw: 20mW, ganho: 6,310^4 e 1 varredura.
Figura IV.1.10: Variações térmicas dos parâmetros dos espectros de RFM de escapos, flagelos e extratos de antena. (a) campo de ressonância – Hr, (b) largura de linha – ΔHpp (c) fator de assimetria – Aw (d) 2ª integral ou área do espectro de absorção.
Figura IV.1.11: Variação da largura de linha com o inverso da temperatura para (a) escapos, (b) flagelos e (c) extratos de antenas. Linha cheia é o ajuste com a eq. I.3.9, ΔH 0 = 1313± 28 Oe, 1238 ± 18 Oe e 1519 ± 25 Oe e KV= (6,9±0,8)10 -14^ erg, (7,6±0,4)10 -14^ erg, e
(6,6±0,5)*10 -14^ para escapos, flagelos, e extratos de antenas, respectivamente.
Figura IV.1.12: variação térmica dos espectros de (a) cabeças, (b) abdomens e (c) tórax. Pmw: 20mW, ganho: 6,3*10^4 e 1 varredura.
Figura IV.1.13: Variações térmicas dos parâmetros de RFM de extratos de cabeça, abdômen, tórax e antena de formiga. (a) campo de ressonância, (b) largura de linha ΔHpp Hr, (c) fator de assimetria Aw e (d) 2ª integral da derivada da absorção S obtida pelo winepr.
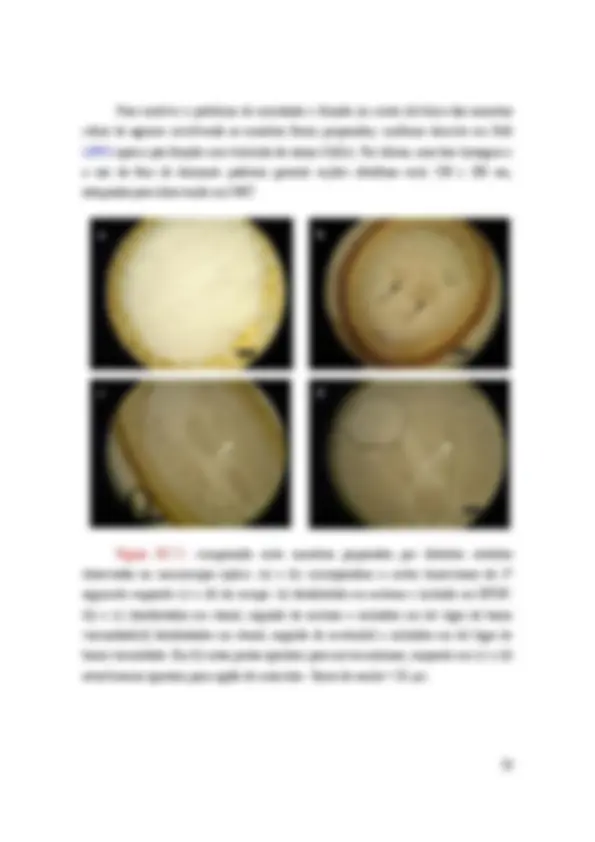
Figura IV.2.1: (a) Solução de solo e seu respectivo padrão de difração (b). Micrografias de partículas de tórax obtidas pelo método de extração magnética (Set/06) (c) e seu padrão de difração (d).
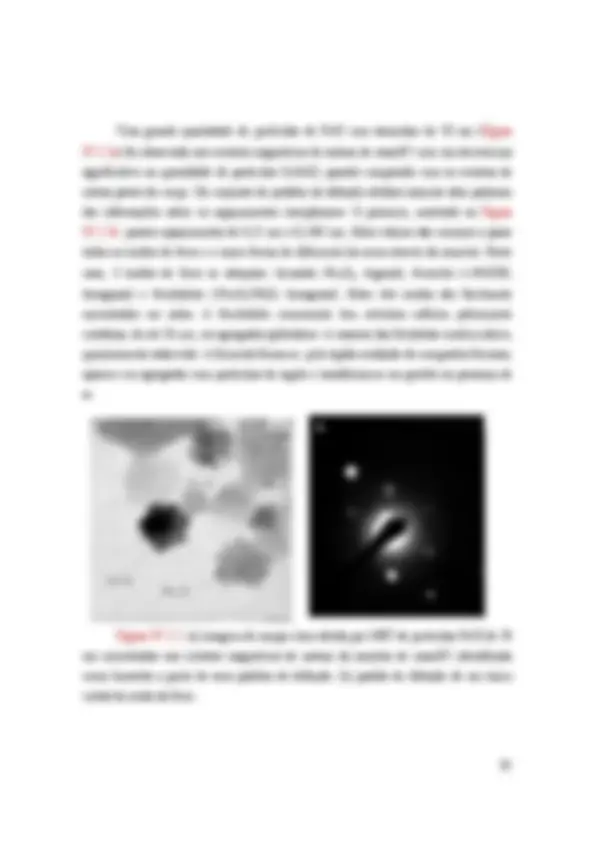
Figura IV.2.2: (a) imagem de campo claro obtida por MET de partículas Fe/O de 50 nm encontradas nos extratos magnéticos de antena da amostra de maio/07 identificada como hematita a partir de seus padrões de difração. (b) padrão de difração de um único cristal de óxido de ferro.
Figura IV.2.3: (a) Dados de EDS obtidos a partir de um único cristal contendo Fe e O. (b) padrão de difração de um destes cristais.
b
xiii
Figura IV.4.5: (a) diagrama mostrando flagelo antenal. Seta aponta para o último segmento. (b) MEV de parte deste segmento. Seta aponta para sensila basicônica.
Figura IV.4.6: Micrografias obtida por MET de cortes longitudinais de sensilas do escapo contendo partículas de solo (setas). Através da análisa qualitativa por EDS foi possível detectar a presença de Al, Si, Fe, O e Ti.
Figura IV.4.7: MET de sensila mecanoreceptora do escapo. Seta preta: material eletrondenso. Seta branca dendrito sob a forma de um cílio.
Figura IV.5.1: Imagens obtidas por MET de cortes transversais ultrafinos da junção entre o terceiro segmento e o pedicelo. (a) botão quitinoso (diâmetro, 4-5 μm). A seta branca aponta
para um longo processo sensorial (diâmetro, 1 μm). Inserção: MEV da antena de P.
contendo partículas. (c) Região aumentada do botão quitinoso na qual cristais de hematita e goetita (seta) além de silicatos / alumosilicatos foram identificados. (d) Padrão de difração do cristal de goetita (seta em c). d = 0,338; 0,256 e 0,257 nm. (e) EDS da região da figura (b)
Figura IV.5.2: (a) Imagens obtidas por MET de cortes transversais ultrafinos da junção entre o pedicelo e o escapo mostrando parte de uma estrutura com partículas circundando-a. Os eixos do elipsóide são mostrados. Inserção: imagem de MEV mostrando a região da junção onde as partículas foram encontradas, parte do escapo (S) e pedicelo (P). (b) EDS das partículas da inserção (região similar de outra grade) mostrando uma grande quantidade de ferro. Inserção: seta aponta para uma partícula identificada como magnetita/maghemita pelo seu padrão de difração de elétrons. d = 0,37, 0,264 nm.
Figura IV.5.3: Torque magnético atuando na estrutura proprioceptora encontrada na junção entre escapo e pedicelo, em unidades de energia térmica, como função da susceptibilidade magnética , k, e da espessura da camada de partículas, t, para campo magnético local de 0, Oe. A estrutura do tipo célula é considerada como um elipsóide com eixos (2c = 35 μm, 2a = 2b = 7 μm).
xiv
Figura IV.5.4: Imagens obtidas por MET de partículas na junção entre escapo e cabeça. (a) dois cristais grandes formados por Fe, Ti e O e o padrão de difração da partícula (seta branca) (b). (c) cristal grande de Fe e O. (d) EDS da partícula apontada em (c).
xvi
Tabela IV.2.1: Comparação das distâncias interplanares experimentais (d), em nanometros (nm), dos extratos do corpo da formiga e soluções do solo e três óxidos de ferro. As porcentagens entre parênteses são as intensidades relativas medidas a partir de difração de raios-X de amostras policristalinas e estão diretamente relacionadas a probabilidade na qual cada ponto de difração apareça em qualquer padrão de difração. Estes dados foram obtidos do Joint Commitee for Powder Diffraction Standards (JCPDS). Os números correspondentes aos óxidos de ferro são 19-629 para magnetita, 13-534 para hematita e 29-713 para goetita.
Tabela IV.5.1: Comparação de porcentagens atômicas obtidas por EDS em partículas do solo e das junções: entre o escapo e pedicelo (J E/P), cabeça e escapo (J C/E) e num botão quitinoso entre o pedicelo e 3 o^ segmento. Entre parênteses estão os aumentos utilizados para cada medida. Na coluna outros elementos incluem-se C, Cu, Na, K e Os.
Introdução
A influência do campo geomagnético em seres vivos vem sendo estudada através de fenômenos naturais, como migração, volta ao lar, etc. Há cerca de três décadas cristais magnéticos foram observados em bactérias magnetotácticas, com a evidência física do fenômeno de magnetotaxia através do alinhamento passivo destas bactérias por um campo magnético (Blakemore 1975). Estudos comportamentais posteriores têm mostrado que o campo magnético terrestre é mais uma fonte de informação do meio ambiente para orientação e navegação de um grande número de animais. Magnetorecepção em animais é um mecanismo complexo, ainda pouco compreendido, que consiste em detectar o campo geomagnético, transferir a informação recebida para o sistema nervoso, onde é processada e utilizada para distintos propósitos. Entre as hipóteses e modelos que discutem a natureza dos receptores magnéticos, atualmente, a ferromagnética, que sugere nanopartículas magnéticas biomineralizadas como transdutores da informação, e um compasso químico baseado num par de radicais são as mais discutidas. Partículas magnéticas foram encontradas em tecidos específicos ou em extratos de partes do corpo de animais (O´Leary et al. 1981, Vilches-Troya et al. 1984, Mann et al. 1988, Walker et al. 1997, Diebel et al. 2000, Hanzlik et al. 2000), mas somente na truta rainbow Oncorhynchus mykiss (Walker et al. 1997) e nos pombos correio (Hanzlik et al. 2000, Fleissner et al. 2003) a magnetita encontrada parece ter uma conexão neural direta. Estudos comportamentais com insetos sociais tais como cupins, abelhas e formigas mostraram respostas orientacionais e navigacionais ao campo geomagnético (Wiltschko & Wiltschko 1995, Vácha 1997, Walker et al. 1997). A observação de partículas de magnetita superparamagnéticas (SPM) no abdômen de abelhas Apis mellifera levou à proposta de um sistema magnetoreceptor baseado nestas partículas (Gould et al. 1978), que supostamente também foram observadas como grânulos nos trofócitos destas abelhas ( Hsu et al. 2007). As formigas, ou pelo menos a maior parte das espécies, são reconhecidas como animais “químicos”. Os feromônios, principais mediadores da comunicação destes animais, são usados por forrageadoras para transferir informações a outras operárias sobre fontes de alimentos ou para alertar outras formigas em relação a um ataque iminente. Marcas do meio