Baixe Enhancing Cell Growth & Cellulose Production in G. xylinum: Novel Cell Recovery Method e outras Notas de estudo em PDF para Engenharia Elétrica, somente na Docsity!
Partial bioenergetic characterization of Gluconacetobacter
xylinum cells released from cellulose pellicles by a novel
methodology
J.L. Cha´vez-Pacheco^1 , S. Martı´nez-Yee^1 , M.L. Contreras^1 , S. Go´ mez-Manzo^1 ,
J. Membrillo-Herna´ ndez^2 and J.E. Escamilla^1
(^1) Instituto de Fisiologı´a Celular, and 2 Instituto de Investigaciones Biome´dicas, Universidad Nacional Auto´noma de Me´xico, Me´xico
2005/0027: received 11 January 2005, revised 11 May 2005 and accepted 20 May 2005
ABSTRACT
J. L. C H A´ V E Z - P A C H E C O , S. M A R T I´N E Z - Y E E , M. L. C O N T R E R A S , S. G O´ M E Z - M A N Z O , J. M E M B R I L L O - H E R N A´ N D E Z A N D J. E. E S C A M I L L A. 2005.
Aims: Gluconacetobacter xylinum is well known for its ability to produce large amounts of cellulose, however, little
is known about its cell physiology. Our goal was to study the respiratory metabolism and components of the
respiratory system of this bacterium in static cultures. To reach our goal, a medium formulation had to be designed
to improve cell growth and cellulose production together with a novel method for the recovery of cells from cellulose
pellicles.
Methods and Results: Successive modifications of a nutrient medium improved G. xylinum cell growth 4Æ5-fold
under static culture conditions. A blender homogenization procedure for the releasing of cells from the cellulose
matrix gave a high yield of cells recovered. Respiratory activities of purified cells were greatly stimulated by
exogenous substrates and showed to be resistant to KCN. Unexpectedly, exogenous NADH was oxidized at high
rates. Cytochromes a, b, c and d were identified after spectral analyses.
Conclusions: Partial bioenergetic characterization of G. xylinum cells allowed us to propose a scheme for its
respiratory system. In addition, the growth medium for biomass production and the procedure for the efficient
recovery of cells from cellulose pellicles were significantly improved.
Significance and Impact of the Study: This work provides the first-ever bioenergetic characterization of
G. xylinum grown in static cultures. In addition, a novel methodology to obtain purified cells in suitable quantities
for biochemical research is described.
Keywords: bacterial cellulose, bacterial cytochromes, Gluconacetobacter xylinum.
INTRODU CTION
The gram-negative bacterium Gluconacetobacter xylinum is well known for its prolific production of bacterial cellulose, which was first described by Brown (Brown 1886). Bacterial cellulose is endowed with unique physical properties, including a highly crystalline ultrafine fibre network of high
purity, therefore is a biological material with many potential industrial uses (for a review see Yamanaka and Watanabe 1994). A major effort has been dedicated to improve biotechnological production of cellulose in cultures (Ross et al. 1991) by testing selected strains (Toyosaki et al. 1995), different fermentor designs (Chao et al. 2001; Cheng et al. 2002; Krystynowicz et al. 2002; Mormino and Bungay 2003), different nutrient formulations (Naritomi et al. 1998a,b; Ramana et al. 2000) and by modifying partial O 2 and CO 2 pressures (Kouda et al. 1997; Hwang et al. 1999).
Correspondence to: J.E. Escamilla, Instituto de Fisiologı´a Celular, Universidad Nacional Auto´noma de Me´xico. Apdo. Postal 70-242 Me´xico City 04510 (e-mail: [email protected]).
ª 2005 The Society for Applied Microbiology
Journal of Applied Microbiology 2005, 99, 1130–1140 doi:10.1111/j.1365-2672.2005.02708.x
Despite the fact that O 2 availability in G. xylinum cell cultures has been recognized as a crucial factor for the production of cellulose, the study of the bioenergetic properties of this bacterium has been overlooked mainly due to the lack of a proper procedure to obtain cells from the cellulose matrix (Watanabe and Yamanaka 1995; Verschuren et al. 2000). Remarkably, notwithstanding this limitation, Benziman and Galanter (1964) carried out seminal experi- ments on the partial purification and characterization of the respiratory chain-linked malate dehydrogenase. Later on, the same group identified Ubiquinone 10 as constituent and functional part of the respiratory system and provided a tentative composition of membrane cytochromes (Benziman and Goldhamer 1968). More recently, Matsuoka et al. (1996) reported on the respiratory activities associated to membranes and determined the H+^ :O 2 ratios evoked by several substrates in intact cells. It is important to note that all the above studies were performed in cells obtained from shaking cultures, a growth condition where cellulose is made as an irregular-shape product with low value for biotech- nological applications. In static cultures, G. xylinum cells are confined within the growing cellulose network and remain firmly attached after harvesting. To free cells from cellulose pellicles, hand squeezing or gentle mechanical treatments have been used to prepare limited cell quantities for further studies (Swissa et al. 1980; Hwang et al. 1999; Son et al. 2001; Heo and Son 2002); alternatively, cellulose accumulation has been counteracted by the addition of commercial cellulase to cultures (Matsuoka et al. 1996); in both cases, low cellular yields are obtained therefore hampering biochemical studies where high quantities of biomass are required. In this communication, a partial bioenergetic character- ization of whole cells of G. xylinum was carried out to get insight into the respiratory metabolism and into the nature of molecular components of its respiratory system. At the same time, culture conditions and the cell release procedure to obtain higher cells yield were improved.
MATERIALS AN D METHO DS
Organism and culture conditions
Storage conditions. Gluconacetobacter xylinum IFO 13693 was kept at ) 70 °C in a basal-modified medium (BM- medium) plus glycerol, containing (g l)^1 ): sucrose, 50; yeast extract, 5; KH 2 PO 4 , 3, MgSO 4 .7H 2 O, 0 Æ5, and glycerol, 10%. The pH of the medium was adjusted to 6Æ0. The BM-medium has the same components of the WY- medium reported earlier (Watanabe and Yamanaka 1995) except that our BM-medium does not contain ammonium sulfate.
Culture conditions. The strain was activated in 250 ml Erlenmeyer flasks containing 100 ml of BM-medium, incubated at 30°C for 48 h at 150 rev min)^1. To optimize cell growth yields in static cultures, the BM-medium was modified as indicated in Fig. 1. Static 500 ml cultures were grown in 2 l Fernbach flasks and incubated for 10 days at 30 °C. Ethanol (1Æ4% v/v), sugar (5% w/v) and nitrogen sources (1% w/v) were added as indicated in Fig. 1 legend. To verify the buffer capacity of the media, potassium phosphate concentration was changed in the ranges of 20–200 mmol l)^1 , initial pH was adjusted to 6Æ 0 with NaOH.
Preparation of isolated cells from cellulose pellicles Cell recovery. Pellicles [800 g wet weight (ww)] were harvested after 10 days of growth on square aluminium trays (45 · 35 · 8 cm) containing 1 l of the selected culture medium. Pellicles were suspended in 2 l of potassium phosphate 100 mmol l)^1 , pH 6 Æ 0 and homogenized (six cycles of 30 s, resting on crushed ice for 5 min between cycles) in an industrial blender (5 l jar) at 4°C. The resulting suspension was filtered through a piece of felt mesh (commercial fabric) and centrifuged at 8670 ·g for 10 min. Filter-retained cellulose was subjected to a second round of blending. Cell pellets were mixed, washed twice with same phosphate buffer and quantified as ww. In some experi- ments, the homogenization buffer was supplemented with NaCl (0Æ1 mol l)^1 ) or Tween 20 (0Æ1%) in order to promote cell release. Cellulose residues were boiled in NaOH 0 Æ5 mol l)^1 for 30 min, thoroughly washed with distilled water and quantified as dry weight (dw) after 5 h at 80°C.
Electron microscopy analysis. Cellulose and cells were fixed in buffered glutaraldehyde 2Æ5% followed by osmium tetroxide 1%. Fixed samples were washed once with potassium phosphate 0Æ1 mol l)^1 , pH 7Æ0 and dehydrated by the addition of alcohol (from 30 to 100%). Preparations were dried to critical point and gold stain was carried out before visualization in a JEOL JSM- 5410LV scanning microscope (Molinari et al. 1998).
Respiratory activities and spectral analyses Oxidase activities. Oxidase activities at 30°C in whole cells were determined with a Clark oxygen electrode using a 53YSI oxygenmeter as previously described (Contreras- Zentella et al. 2003). Briefly, cells were suspended in a final volume of 2 ml of potassium phosphate buffer (100 mmol l)^1 pH 6Æ0 or 7Æ4). Respiratory substrates were added at 10 mmol l)^1 (glucose, ethanol or acetaldehyde) with the exception of NADH (5 mmol l)^1 ). The reaction
G. X Y L I N U M R E S P I R A T O R Y S Y S T E M 1131
our experimental conditions, grenetin did not improve cell growth. By contrast, sodium glutamate increased cell growth by about 1Æ5-fold compared with that of the BM-medium supplemented with ethanol plus glucose (compare Fig. 1c,d). Casein hydrolysate and ammonium sulfate were less effective (about 1Æ2 and 1Æ1-fold, respectively). Our results are consistent with a recent report where sodium glutamate, among other compounds, was identified as a good nitrogen source for G. xylinum static cultures (Ramana et al. 2000). Taken together, these results indicate that the modifications made to the BM-medium (addition of ethanol, glucose instead of sucrose, sodium glutamate and 100 mmol l)^1 phosphate) favour cell growth (about 4Æ5-fold higher) and cellulose production (only 1Æ8-fold higher) in static cultures. Our final medium formula, here called BEGG-medium contains (g l)^1 ): glucose, 50; yeast extract, 5; sodium glutamate, 10; MgSO 4 .7H 2 O, 0Æ5; KH 2 PO 4 , 13Æ 6 and ethanol 1Æ4% (v/v).
Gluconacetobacter xylinum cell recovery from cellulose pellicles
A reliable method for the recovery of G. xylinum cells trapped in cellulose pellicles has been addressed here. The hand-squeezing of pellicles is the standard procedure used to obtain low amounts of cells (Hwang et al. 1999; Son et al. 2001). Moreover, cellulose accumulation has been counteracted by the costly addition of cellulase (Matsuoka et al. 1996). Thus, it was decided to look for a more simple and reliable method. Preliminary attempts showed that mechanical homogenization of pellicles in a home blender liberated significant amounts of cells; therefore, it was decided to scale up the procedure in a commercial blender (3 l working volume in a 5 l jar) using six 30 s homog- enization cycles with intervals of 5 min, in an ice bath between cycles. Thereafter, the homogenizate was filtered through a felt mesh. The filtrate thus obtained was centrifuged for 10 min, at 8670 ·g. Pelleted cells were washed twice with 100 mmol l)^1 potassium phosphate buffer (pH 6 Æ0). The use of blender homogenization showed a significant increase in cell recovery (about twofold; Table 1).
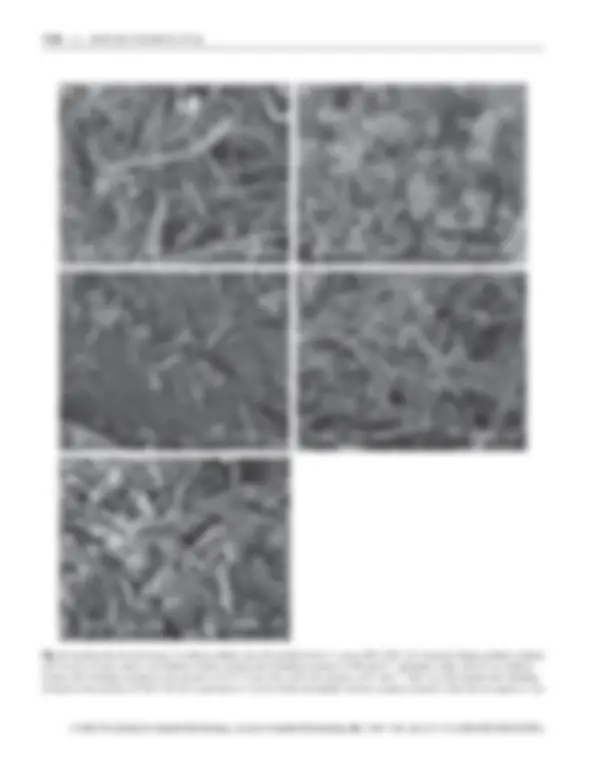
Scanning electron microscopy of cellulose pellicles before and after the blending treatment (Fig. 2a,b, respectively) showed that significant amounts of released cells form aggregates (Fig. 2b), suggesting that cell aggregates were retained together with cellulose particles by the felt mesh. In order to prevent cell aggregation, the blender homogeniza- tion step was either performed in the presence of a gently detergent (0Æ1% Tween 20, Fig. 2c) or salt (0Æ1 mol l)^1 NaCl, Fig. 2d). In both cases, cell cluster dispersion was evident. The addition of Tween 20 or NaCl increased biomass recovery about 2 Æ5–3Æ6-fold, respectively when compared with the hand squeeze protocol (Table 1). It is important to note that cells recovered after blender homo- genization were apparently intact but still moderately contaminated with cellulose debris that could not be washed out by centrifugation (Fig. 2e).
Respiratory activities of Gluconacetobacter xylinum purified cells Physiological studies of cells released by hand-squeeze or blender homogenization were compared by determining respiration activities of periplasmic PQQ-dehydrogenases and membrane-bound NADH dehydrogenase evoked by different substrates, such as glucose, ethanol or acetalde- hyde. As shown in Table 1, cells obtained by hand- squeezing, as the most gentle procedure, displayed the highest activities, close followed by cells recovered after blender homogenization in buffer alone and buffer plus NaCl. Respiratory activities of cells released in buffer plus Tween 20 were consistently slightly lower. Surprisingly, exogenous NADH evoked high respiratory activities in cells released by blending. As NADH oxidation by Complex I (NADH dehydrogenase) takes place at the inner side of the cytoplasmic membrane and exogenous NADH usually does not permeate the membrane (Overkamp et al. 2000), membrane damage caused by the blender treatment was suspected; however, cells released by the gently hand- squeeze treatment showed similar NADH oxidation activ- ities (Table 1). Respiration evoked by exogenous NADH in whole cells was previously reported for Methylophilus methylotrophus, however it was later shown to be an artefact
Table 1 Biomass yields and substrate stimulated respiration of G. xylinum cells released from cellulose pellicles (^) Treatment Biomass (g wet cells l) (^1) )
O 2 uptake (nmol O 2 min)^1 mg)^1 wet cells) Acetaldehyde Ethanol Glucose NADH
Hand squeeze 1 Æ 1 80 23 8 30 Blender homogenization: Buffer 2 Æ 1 70 18 8 32 Buffer plus NaCl 3 Æ 6 75 19 7 33 Buffer plus Tween 20 2 Æ 5 53 13 4 25
G. X Y L I N U M R E S P I R A T O R Y S Y S T E M 1133
(a) (b)
(c)
(e)
(d)
Fig. 2 Scanning electron microscopy of cellulose pellicles and cells purified from G. xylinum IFO 13693. (a) Untreated cellulose pellicles obtained after 10 days of static culture. (b) Cellulose residues obtained after blending treatment in 100 mmol l)^1 phosphate buffer, pH 6Æ0. (c) Cellulose residues after blending treatment in the presence of 0Æ1% Tween 20 or (d) in the presence of 0Æ1 mol l)^1 NaCl. (e) Cells purified after blending treatment in the presence of NaCl. The bar is equivalent to 1 lm for all the micrographs with the exception of panel b where the bar equals to 5 lm
1134 J. L. C H A´ V E Z - P A C H E C O ET AL.
difference spectra were carried out, but in this case, in a buffer containing 25% PEG 3350 instead of DMSO (see above). The peak at 631 nm was bleached in the presence of 5 mmol l)^1 KCN (Fig. 4b) giving a strong support to the identification of this signal as evidence for the presence of a type-d cytochrome (Gil et al. 1992; Ju¨nemann 1997). Carbon monoxide reactive cytochromes were identified using glucose as electron donor (Fig. 5a). The resulting 77 K spectrum was rather complex, indicating the presence of several CO-reactive pigments: A putative cytochrome c- CO adduct was indicated by a peak at 417 nm with troughs at 522 and 550 nm (Fig. 5a). A similar spectrum was reported for the CO-reactive cytochrome c 553 purified from Gluconobacter suboxydans (Matsushita et al. 1981) and also for the cytochrome c-CO adduct detected in membranes of Thiobacillus tepidarius (Kelly et al. 1993). Features in the 425–431 nm region could have contributions of CO adducts of type-c, type-b and type-a cytochromes (Wood 1984; Kelly et al. 1993). The presence of a type-a cytochrome –CO adduct was suggested by a small inflexion at 431 nm, the deep trough at 443 nm and the a-peak at 593 nm; these signals closely resemble those reported for the cytochrome a-CO adduct of cytochrome ba 3 of Gluconacetobacter diazo- trophicus (Flores-Encarnacio´n et al. 1999). In addition, the presence of a type-d cytochrome-CO adduct was indicated
by the a-peak at 640 nm and trough at 626 nm (Ju¨nemann 1997). The photodissociation spectrum (Fig. 5b) clearly showed the photolysis of a cytochrome a-CO adduct with troughs at 431 and 593 nm and peak at 443 nm. Addition- ally, a conspicuous peak at 415 nm was observed, however spectral features in the a-region (see 550–560 nm region) of the spectrum were not clear enough to allow an accurate identification of this pigment. As expected, no spectral features around 626 nm were detected, again supporting the idea of the presence of a type-d cytochrome. The lack of photodissociation signal for type-d cytochromes is due to the extremely fast postphotolysis reassociation rate of CO with type-d cytochromes, this precludes the detection of photo- dissociated species at 77 K (Muntyan et al. 1995).
DISCUSSION
Physiological studies of G. xylinum have been mainly focused on biotechnological aspects of bacterial cellulose production in culture (Legge 1990; Sutherland 1998). Accordingly, culture media formulations (Naritomi et al. 1998a,b; Ramana et al. 2000) and culture conditions (Chao et al. 2001; Cheng et al. 2002; Krystynowicz et al. 2002) have been designed to try to maximize cellulose synthesis, leaving aside cell growth physiology; moreover, the intimate
591 526
554
(^631651)
531
562 523
440
4
2
1
3
A: 0·025 A: 0·
Wavelength (nm)
589
428
3
2 4 1
651
631
(^523 ) 550 553
440
425
415
5
4
3
2
1
5
4
3
2
1
A: 0·
A: 0·
(b)
(a)
Fig. 4 Reduced difference spectra recorded at 77 K of purified cells of G. xylinum IFO 13693 grown in static cultures. (a) Reduced difference spectra were generated by addition of ethanol, dithionite, NADH, acetaldehyde or glucose (traces 1–5, respectively) to the samples and ammonium persulfate to refer- ence (oxidized) cell suspensions. Cell suspen- sions (40 mg ww ml)^1 ) were prepared in 100 mmol l)^1 phosphate buffer, pH 6Æ0 con- taining 25% (v/v) DMSO. (b) KCN differ- ence spectra of cells reduced by ethanol or acetaldehyde (traces 1 and 2, respectively). Ethanol and acetaldehyde reduced difference spectra (traces 3 and 4, respectively) are shown for comparison. The difference spectra were carried out in 0Æ1 mol l)^1 phosphate buffer, pH 6Æ0 containing 25% (w/v) PEG 3350 (instead of DMSO); KCN (5 mmol l)^1 ) was added to samples 3 min before substrates
1136 J. L. C H A´ V E Z - P A C H E C O ET AL.
association of cells with excreted cellulose renders cell growth determinations a difficult task, specially in the case of static cultures. Lack of information on this issue and our interest to obtain suitable amounts of cells biomass for biochemical studies, prompted us to design a protocol for an easy and efficient recovery of cells from cellulose pellicles. Previous reports barely described methodology for cell recovery and did not offer quantitative considerations (Swissa et al. 1980; Hwang et al. 1999; Son et al. 2001). Our results showed that blender homogenization renders up to 3Æ6 fold more cells (Table 1). Cells thus obtained seem to be physiologically competent as judged by electron micro- scopy (Fig. 2e) and respiratory properties (Table 1). The medium formulation was optimized to increase cell growth and cellulose production in static cultures. Several authors have shown that G. xylinum requires a complex medium for optimal cellulose production (Heo and Son 2002); therefore, a medium containing sucrose plus yeast extract as base nutrients (BM-medium) was modificated to improve cell growth. After several tries, it was found that cell growth and cellulose production were increased about 4 Æ5 and 2 fold respectively when the BM-medium was supplemented with 1Æ4% ethanol, 1% sodium glutamate and the original sucrose was replaced by 5% glucose (BEGG- medium). Glucose seemed to be a better carbon source than sucrose, very probably because its utilization as electron donor and carbon source does not require previous hydro- lysis as it is the case for sucrose. Glucose contribution as respiratory substrate could be low in our strain and/or under the culture conditions tested, as suggested by the low glucose oxidase activity exhibited in whole cells (Table 1).
In this regard, it has been proposed that in cellulose- producing strains of G. xylinum, glucose may be mainly used for cellulose synthesis, not generating energy for growth (Matsuoka et al. 1996). On the other hand, ethanol added to increase cell growth must serve as the primary energy source, as suggested by the high respiratory activities evoked by ethanol and by its oxidation product, acetaldehyde (Table 1). In this context, it has been proposed that in the early stages of growth, several substrates like lactate, ethanol, acetaldehyde, pyruvate and acetate divert the metabolic sugar flow from cellulose production towards the tricarboxylic acid cycle, resulting in the stimulation of cell growth (Son et al. 2001). Addition of mixtures or individual amino acids to culture media designed for G. xylinum have a positive impact on cell growth and cellulose production, such is the case of methionine for G. xylinum ssp. sucrofermentans (Matsuoka et al. 1996) or glutamate for Gluconacetobacter sp. (Ramana et al. 2000). Accordingly, our BEGG-medium, having yeast extract as primary nitrogen source and sodium glutamate as supplement rendered the highest growth yields of G. xylinum (Fig. 1d). Whole cells liberated from cellulose pellicles showed the unexpected ability to oxidize exogenous NADH. The possibility that this result was an artefact caused by cellular membrane disruption during the blender homogenization treatment was ruled out, since cells obtained by gently hand squeeze showed similar NADH oxidation rates (Table 1). Still, mechanical (squeeze or blender) detachment of cells from cellulose fibres undergoing synthesis may cause enough membrane damage to explain permeation of exogenous
522
443
Wavelength (nm)
A: 0·
560
640
443
(^431 )
A: 0·
550 (b)
(a) A: 0· (^431 )
560
522 550
553
425
417
415
593 640
626
A: 0·
Fig. 5 Carbon monoxide reactive pigments in cells of G. xylinum IFO 13693. The spectra were recorded at 77 K (a) CO- difference spectrum reduced by glucose. (b) Photodissociation difference spectrum of CO-cytochrome adducts reduced by glucose. Cells (40 mg ww ml)^1 ) were suspended in 100 mmol l)^1 phosphate buffer, pH 6Æ0, containing 25% (v/v) DMSO and 10 mmol l)^1 glucose, and incubated for 1 h at room temperature. Thereafter, a gently stream of CO was bubbled, before freezing, and the spectrum was recorded. The photodissociation spectrum was obtained as described in Materials and methods
G. X Y L I N U M R E S P I R A T O R Y S Y S T E M 1137
Paredes for technical assistance and to Virginia Godı´nez for secretarial assistance.
REFEREN CES
Benziman, M. and Galanter, Y. (1964) Flavine adenine dinucleotide- linked malic dehydrogenase from Acetobacter xylinum. J Bacteriol 88 , 1010–1018. Benziman, M. and Goldhamer, H. (1968) The role of ubiquinone in the respiratory chain of Acetobacter xylinum. Biochem J 108 , 311–316. Brown, A.J. (1886) An acetic ferment which forms cellulose. J Chem Soc 49 , 432–439. Chao, Y., Sugano, Y. and Shoda, M. (2001) Bacterial cellulose production under oxygen-enriched air at different fructose concen- trations in a 50-liter, internal loop airlift reactor. Appl Microbiol Biotechnol 55 , 673–679. Cheng, H.P., Wang, P.M., Chen, J.W. and Wu, W.T. (2002) Cultivation of Acetobacter xylinum for bacterial cellulose produc- tion in a modified airlift reactor. Biotechnol Appl Biochem 35 , 125–
Contreras-Zentella, M., Mendoza, G., Membrillo-Herna´ndez, J. and Escamilla, J.E. (2003) A novel double heme substitution produces a functional bo 3 variant of the quinol oxidase aa 3 of Bacillus cereus. J Biol Chem 278 , 31473–31478. Flores-Encarnacio´n, M., Contreras-Zentella, M., Soto-Urzu´a, L., Aguilar, G., Baca, B.E. and Escamilla, J.E. (1999) The respiratory system and diazotrophic activity of Acetobacter diazotrophicus PAL5. J Bacteriol 181 , 6987–6995. Gil, A., Kroll, R.G. and Poole, R.K. (1992) The cytochrome composition of the meat spoilage bacterium Brochothrix thermosph- acta: identification of cytochrome a 3 - and d-type terminal oxidases under various conditions. Arch Microbiol 158 , 226–233. Heo, M.S. and Son, H.J. (2002) Development of an optimized, simple chemically defined medium for bacterial cellulose production by Acetobacter sp. A9 in shaking culture. Biotechnol Appl Biochem 36 , 41–45. Hwang, J.W., Yang, Y.K., Hwang, J.K., Pyun, Y.R. and Kim, Y.S. (1999) Effects of pH and dissolved oxygen on cellulose production by Acetobacter xylinum BRC5 in agitated cultures. J Biosci Bioeng 88 , 183–188. Ju¨nemann, S. (1997) Cytochrome bd terminal oxidase. Biochim Biophys Acta 1321 , 107–127. Kelly, D.P., Lu, W.-P. and Poole, R.K. (1993) Cytochromes in Thiobacillus tepidarius and the respiratory chain involved in the oxidation of thiosulfate and tetrathionate. Arch Microbiol 160 , 87–
Kouda, T., Naritomi, K., Yano, H. and Yoshinaga, F. (1997) Effects of oxygen and carbon dioxide pressures on bacterial cellulose produc- tion by Acetobacter in aerated and agitated cultures. J Ferment Bioeng 84 , 124–127. Krystynowicz, A., Czaja, W., Wiktorowska-Jezierska, A., Goncalves- Miskiewicz, M., Turkiewicz, M. and Bielecki, S. (2002) Factors affecting the yield and properties of bacterial cellulose. J Ind Microbiol Biotechnol 29 , 189–195. Legge, R.L. (1990) Microbial cellulose as a speciality chemical. Biotechnol Adv 8 , 303–319.
Matsuoka, M., Tsuchida, T., Matsushita, K., Adachi, O. and Yoshinaga, F. (1996) A synthetic medium for bacterial cellulose production by Acetobacter xylinum subsp. sucrofermentans. Biosci Biotechnol Biochem 60 , 575–579. Matsushita, K., Tayama, K., Shinagawa, E., Adachi, O. and Ameyama, M. (1981) Purification and characterization of CO-binding cyto- chrome c 553 from the membrane of Gluconobacter suboxydans. FEMS Microbiol Lett 10 , 267–270. Molinari, J.L., Tato, P., Rodrı´guez, D., Solano, S., Rubio, M. and Sepu´lveda, J. (1998) Impairment of the inflammatory reaction on implanted Taenia solium metacestodes in mice by a T. solium RNA- peptide: a scanning electron microscopy study. Parasitol Res 84 , 173–180. Mormino, R. and Bungay, H. (2003) Composites of bacterial cellulose and paper made with a rotating disk bioreactor. Appl Microbiol Biotechnol 62 , 503–506. Muntyan, M.S., Ustiyan, V.S., Viryasov, M.B. and Skulachev, V.P. (1995) Terminal oxidases of the bb- and caa 3 -types in Bacillus sp. FTU. Biochem Biophys Res Commun 207 , 55–61. Naritomi, T., Kouda, T., Yano, H. and Yoshinaga, F. (1998a) Effect of ethanol on bacterial cellulose production from fructose in continuous culture. J Ferment Bioeng 85 , 598–603. Naritomi, T., Kouda, T., Yano, H. and Yoshinaga, F. (1998b) Effect of lactate on bacterial cellulose production from fructose in continuous culture. J Ferment Bioeng 85 , 89–95. Overkamp, K.M., Bakker, B.M., Kotter, P., van Tuijl, A., De Vries, S., van Dijken, J.P. and Pronk, J.T. (2000) In vivo analysis of the mechanisms for oxidation of cytosolic NADH by Saccharomyces cerevisiae mitochondria. J Bacteriol 182 , 2823–2830. Patchett, R.A. and Jones, C.W. (1986) The apparent oxidation of NADH by whole cells of the methylotrophic bacterium Methylo- philus methylotrophus. A cautionary tale. Antonie Van Leeuwenhoek 52 , 387–392. Ramana, K.V., Tomar, A. and Singh, L. (2000) Effect of various carbon and nitrogen sources on cellulose synthesis by Acetobacter. xylinum. World J Microbiol Biotechnol 16 , 245–248. Ross, P., Mayer, R. and Benziman, M. (1991) Cellulose biosynthesis and function in bacteria. Microbiol Rev 55 , 35–58. Son, J.H., Heo, M.S., Kim, Y.G. and Lee, S.J. (2001) Optimization of fermentation conditions for the production of bacterial cellulose by a newly isolated Acetobacter sp. A9 in shaking culture. Biotechnol Appl Biochem 33 , 1–5. Sutherland, I.W. (1998) Novel and established applications of micro- bial polysaccharides. Trends Biotechnol 16 , 41–46. Swissa, M., Aloni, Y., Weinhouse, H. and Benziman, M. (1980) Intermediary steps in Acetobacter xylinum cellulose synthesis: studies with whole cells and cell-free preparations of the wild type and a celluloseless mutant. J Bacteriol 143 , 1142–1150. Toyosaki, H., Naritomi, T., Seto, A., Matsuoka, M., Tsuchida, T. and Yoshinaga, F. (1995) Screening of bacterial cellulose producing Acetobacter strains suitable for agitation culture. Biosci Biotechnol Biochem 59 , 1498–1502. Verschuren, P.G., Cardona, T.D., Nout, M.J.R., De Gooijer, K.D. and Van den Heuvel, J.C. (2000) Location and limitation of cellulose production by Acetobacter xylinum established from oxygen profiles. J Biosci Bioeng 89 , 414–419.
G. X Y L I N U M R E S P I R A T O R Y S Y S T E M 1139
Watanabe, K. and Yamanaka, S. (1995) Effects of the oxygen tension in the gaseous phase on production and physical properties of bacterial cellulose formed under static culture conditions. Biosci Biotechnol Biochem 59 , 65–68. Williams, H.D. and Poole, R.K. (1987) The cytochromes of Acetobacter pasteurianus NCIB 6428. Evidence of a role for a cytochrome a 1 -like haemoprotein in electron transfer to cytochrome oxidase d. J Gen Microbiol 133 , 2461–2472.
Wood, P.M. (1984) Bacterial proteins with CO-binding b- or c-type haem. Functions and adsorption spectroscopy. Biochim Biophys Acta 768 , 293–317. Yamanaka, S. and Watanabe, K. (1994) Application of bacterial cellulose. In Cellulosic Polymers, Blends and Composites ed. Gilbert, R.D. pp. 207–215. Munich, Viena, New York: Carl Hanser Verlag.
1140 J. L. C H A´ V E Z - P A C H E C O ET AL.