Baixe Influence of Wing Kinematics on Aerodynamic Force Production in Flapping Wings e outras Notas de estudo em PDF para Engenharia de Produção, somente na Docsity!
To perform aerial maneuvers, insects must not only generate
sufficient lift to remain aloft, they must also manipulate flight
forces with great precision. Although insects are known to use
their legs and abdomen as control surfaces during flight (Arbas,
1986; Zanker, 1988; May and Hoy, 1990; Lorez, 1995), they
steer and maneuver largely by altering wing motion (Götz
et al., 1979; Ennos, 1989; Robertson and Johnson, 1993;
Wortmann and Zarnack, 1993). Thus, a central hurdle in
understanding how insects steer and maneuver is determining
how modifications in stroke kinematics alter the forces and
moments generated by flapping wings.
In a few cases, researchers have attempted to capture the
free-flight kinematics of maneuvering insects (Ennos, 1989;
Ruppell, 1989). While such analyses are essential because they
reveal what insects actually do with their wings when steering,
free-flight studies are limited because it is not yet feasible to
relate the changes in wing kinematics directly to changes in
instantaneous aerodynamic forces. An alternative approach is
The Journal of Experimental Biology 204, 2607–2626 (2001) 2607
Printed in Great Britain © The Company of Biologists Limited 2001 JEB
We used a dynamically scaled mechanical model of the
fruit fly Drosophila melanogaster to study how changes in
wing kinematics influence the production of unsteady
aerodynamic forces in insect flight. We examined 191
separate sets of kinematic patterns that differed with
respect to stroke amplitude, angle of attack, flip timing,
flip duration and the shape and magnitude of stroke
deviation. Instantaneous aerodynamic forces were
measured using a two-dimensional force sensor mounted
at the base of the wing. The influence of unsteady
rotational effects was assessed by comparing the time
course of measured forces with that of corresponding
translational quasi-steady estimates. For each pattern,
we also calculated mean stroke-averaged values of the
force coefficients and an estimate of profile power. The
results of this analysis may be divided into four main
points.
(i) For a short, symmetrical wing flip, mean lift was
optimized by a stroke amplitude of 180 ° and an angle of
attack of 50 °. At all stroke amplitudes, mean drag
increased monotonically with increasing angle of attack.
Translational quasi-steady predictions better matched the
measured values at high stroke amplitude than at low
stroke amplitude. This discrepancy was due to the
increasing importance of rotational mechanisms in
kinematic patterns with low stroke amplitude.
(ii) For a 180 ° stroke amplitude and a 45 ° angle of
attack, lift was maximized by short-duration flips
occurring just slightly in advance of stroke reversal.
Symmetrical rotations produced similarly high
performance. Wing rotation that occurred after stroke
reversal, however, produced very low mean lift.
(iii) The production of aerodynamic forces was sensitive
to changes in the magnitude of the wing’s deviation from
the mean stroke plane (stroke deviation) as well as to
the actual shape of the wing tip trajectory. However, in
all examples, stroke deviation lowered aerodynamic
performance relative to the no deviation case. This
attenuation was due, in part, to a trade-off between lift
and a radially directed component of total aerodynamic
force. Thus, while we found no evidence that stroke
deviation can augment lift, it nevertheless may be used to
modulate forces on the two wings. Thus, insects might
use such changes in wing kinematics during steering
maneuvers to generate appropriate force moments.
(iv) While quasi-steady estimates failed to capture the
time course of measured lift for nearly all kinematic
patterns, they did predict with reasonable accuracy
stroke-averaged values for the mean lift coefficient.
However, quasi-steady estimates grossly underestimated
the magnitude of the mean drag coefficient under all
conditions. This discrepancy was due to the contribution
of rotational effects that steady-state estimates do not
capture. This result suggests that many prior estimates of
mechanical power based on wing kinematics may have
been grossly underestimated.
Key words: flapping flight, quasi-steady force, unsteady aerodynamics, fruit fly, Drosophila melanogaster , added mass, delayed stall, rotational circulation, wake capture.
Summary
Introduction
THE CONTROL OF FLIGHT FORCE BY A FLAPPING WING: LIFT AND DRAG
PRODUCTION
SANJAY P. SANE* AND MICHAEL H. DICKINSON
Department of Integrative Biology, University of California, Berkeley, CA 94720, USA
*e-mail: [email protected]
Accepted 18 May 2001
2608
to measure instantaneous forces on tethered insects (Cloupeau
et al., 1979; Wilkin, 1990; Zanker, 1990b; Zanker and Götz,
1990; Dickinson and Götz, 1996). However, forces and stroke
kinematics measured on tethered insects may not accurately
represent those generated in free flight. Further, since tethered
flight force transducers measure whole-body forces, it is not
possible to resolve the instantaneous aerodynamic forces
generated by individual wings. A third approach is to calculate
the aerodynamic forces generated by arbitrary stroke
kinematics using computational fluid dynamics (Liu et al.,
1998; Wang, 2000). However, because of the critical role of
unsteady mechanisms and three-dimensional flow structure in
insect flight aerodynamics (Ellington et al., 1996; Dickinson et
al., 1999), theoretical or numerical approaches have, as yet,
offered only limited insight into the aerodynamics of steering.
Given the current limitations in studies of both real animals
and numerical simulations, we have chosen to study the
problem of maneuverability using a dynamically scaled model
of a flapping insect. Aerodynamic models have proved
valuable in the study of insect flight, particularly in the
identification and analysis of unsteady aerodynamics (Bennett,
1977; Maxworthy, 1979; Spedding and Maxworthy, 1986;
Dickinson and Götz, 1993; Ellington et al., 1996; Dickinson et
al., 1999). In large part through the use of mechanical models,
researchers have identified an array of mechanisms that
collectively account for the elevated aerodynamic performance
of flapping wings. These include the clap and fling (Spedding
and Maxworthy, 1986), dynamic stall (Dickinson and Götz,
1993; Ellington et al., 1996), rotational lift (Bennett, 1970;
Dickinson et al., 1999) and wake capture (Dickinson, 1994;
Dickinson et al., 1999). Now that the various mechanisms
responsible for the elevated aerodynamic performance of
insect wings have been identified, it is possible to tackle the
question of how animals manipulate such mechanisms to steer
and maneuver.
In this study, we use a dynamically scaled mechanical model
of Drosophila melanogaster to investigate how changes in
wing kinematics affect the production of aerodynamic forces.
In particular, we explore the influence of five behaviorally
relevant kinematic parameters: stroke amplitude, angle of
attack, the timing and duration of wing rotation and stroke
plane deviation. We chose this particular set of parameters
because fruit flies actively vary them during flight maneuvers
(Götz et al., 1979; Zanker, 1990a; Dickinson et al., 1993;
Lehmann and Dickinson, 1998). However, the goal of this
project is not to replicate the precise kinematics of free flying
insects per se , but rather to map aerodynamic forces within a
broad parameter space that encompasses the variation seen
among insects.
From the instantaneous force records, we calculate time-
averaged aerodynamic force coefficients, lift-to-drag ratios and
other measures of aerodynamic performance. The resultant
data set is useful in identifying the kinematic parameters that
most influence the magnitude and direction of aerodynamic
forces generated by flapping wings. In a companion paper
(S. P. Sane and M. H. Dickinson, in preparation), we will
extend the analysis by considering the instantaneous and time-
averaged force moments generated about the yaw, pitch and
roll axes. The comprehensive parameter maps generated in
these studies should be of help to biologists who wish to know
the aerodynamic consequences of observed changes in wing
kinematics as well as to engineers who wish to optimize the
performance of small biomimetic flying robots. In addition,
these data provide experimental validations for numerical
simulations of the fluid motion around flapping wings.
Materials and methods
Most of the instruments and procedures used in these
experiments have been described elsewhere (Dickinson et al.,
1999). We fashioned the wings from 2.3 mm thick acrylic
sheets using an isometrically enlarged planform of a
Drosophila melanogaster wing. The proximal end of the wing
was attached to a two-dimensional force transducer that
measured the forces normal and parallel to the wing surface.
Each force channel measured the shear encountered by two
parallel phosphor-bronze shims equipped with four 350 Ω
strain gauges wired in full-bridge configuration. This design
rendered the sensor nearly insensitive to the position of the
force load on the wing as well as to moments around its central
axis. Forces generated by calibration weights placed at the tip,
base, trailing edge and leading edge differed by less than 5 %.
The final calibration was based on a point load at the wing’s
center of area. The proximal end of the force transducer was
attached to a gearbox capable of three degrees of rotational
motion (Fig. 1A). The distal tip of the wing was located 25 cm
from the center of the gearbox. The gearbox was driven via
three coaxial shafts by three stepper motors. The stepper
motors were attached to the shafts by pulleys and timing belts
with a 1:10 gear reduction, such that each 4.5 ° step of the
motor produced a 0.45 ° rotation of the wing. The wings, force
sensor and gearbox were immersed in a tank of mineral oil with
a viscosity of 120 cSt at room temperature (approximately
25 °C). The viscosity of the oil was chosen to achieve a
Reynolds number in the range of 10 2 , although the exact value
varies according to the kinematics for each trial. Since the
forces on the wing are directly proportional to the density of
the surrounding medium, the oil also serves to increase forces
on the wings and to decrease the signal-to-noise ratio of the
force measurements. Mineral oil provides an additional
advantage of electrically and thermally isolating the sensor and
thus reducing noise fluctuations.
Dynamic scaling
To obtain accurate dynamic scaling of an insect, it is
necessary to keep the values of both the Reynolds number
( Re =4Φ nR^2 ν−^1 AR −^1 , where Φ is stroke amplitude, n is wingbeat
frequency, R is wing length, ν is kinematic viscosity, aspect ratio
AR is 4 R^2 S −^1 and S is the surface area of a wing pair; Ellington,
1984c) and the reduced frequency parameter (body
velocity/wing velocity) constant (Spedding, 1993). For hovering
animals as well as the model fly, the reduced frequency
S. P. S ANE AND M. H. D ICKINSON
2610
parameter is zero by definition, since their body velocity is
zero. The free-flight cruising velocity of D. melanogaster is
approximately 20 cm s−^1 (David, 1978), while the mean velocity
of the wing tip is 280 cm s−^1 (Lehmann and Dickinson, 1997).
Thus, even while flying forwards, the reduced frequency
parameter is less than 0.1, indicating that the effect of free-
stream velocity on force generation should be of secondary
importance to the velocity generated by flapping. For these
reasons, our experiments in still fluid, which match the hovering
case, should also serve as a fair approximation for moderate
forward speeds. Thus, the reduced frequency parameter is not
significantly different for cruising D. melanogaster and our
static, hovering model fly. To obtain the correct range of
Reynolds numbers, we used an isometrically enlarged wing
planform of an actual D. melanogaster wing to ensure that the
shape parameters (Ellington, 1984a) were identical to those
of D. melanogaster. Using available data on D. melanogaster
morphology and kinematics (Lehmann and Dickinson, 1997),
we estimated that a wing length of 0.25 m, a surface area of the
wing pair of 0.0334 m^2 (or 0.0167 m^2 for a single wing), a
kinematic viscosity of 120 cSt and a wingbeat frequency of
0.168 Hz would allow us to achieve Reynolds numbers in the
same range as those of D. melanogaster.
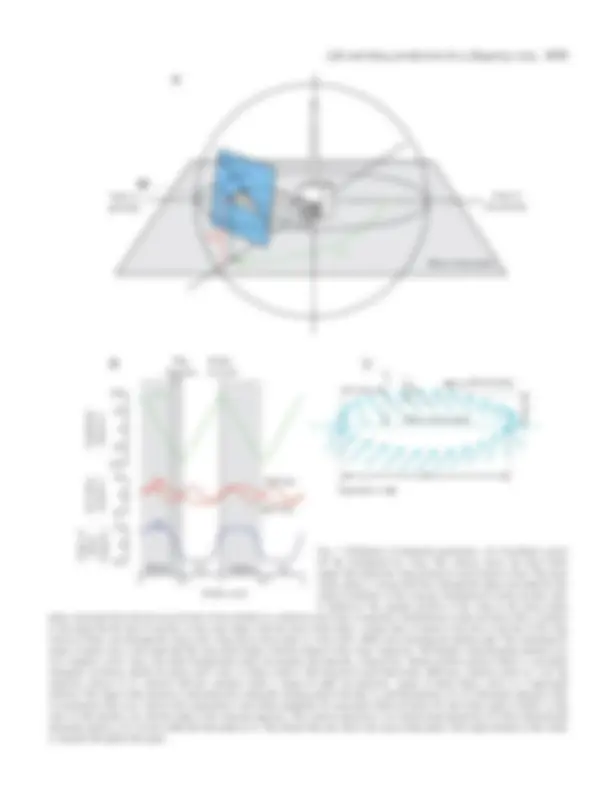
Stroke kinematics
In the absence of wing deformation, the kinematics of the
wings may be uniquely described by specifying the time course
of three angles: stroke position, φ( t ), angle of attack, α( t ), and
stroke deviation, θ( t ) (Fig. 1B). In all experiments, the angular
position of the wing within the stroke plane was described by a
triangular waveform, which maintains a constant translational
velocity throughout each half-stroke. The waveform was
smoothed to minimize inertial accelerations during stroke
reversal and to match more closely published stroke kinematics
from a variety of insects (Ellington, 1984b; Zanker, 1990a).
For smoothing, we filtered the triangular waveform using a
zero-phase-delay low-pass two-pole Butterworth filter with a
cut-off frequency equal to 10 times the stroke frequency of
0.17 Hz. The peak-to-peak amplitude of the stroke angle
waveform could be varied in each experiment. The angle of
attack was described by a trapezoidal wave function, which
maintained a constant angle of attack during each half-stroke
and constant rotational velocity during stroke reversal. The
shape of this waveform in each experiment was determined by
setting the mid-stroke angles of attack during the upstroke and
downstroke and by specifying the starting and stopping points
for wing rotation. The resulting function was then smoothed
using a low-pass filter with identical characteristics to that used
for the stroke position waveform. We used two functions to
describe stroke deviation: an ‘oval’ pattern in which the wing
tip deviated from the stroke plane according to a half-sine-
wave per stroke period and a ‘figure-of-eight’ pattern in which
the stroke deviation varied as a full sine-wave. These patterns
were chosen because they roughly approximate patterns
described for a variety of insects (Ellington, 1984b; Zanker,
1990a).
To create the kinematic patterns used in this study, we varied
any or all of six parameters: (i) the stroke amplitude, (ii) the
mid-stroke angle of attack during upstroke and downstroke,
(iii) the timing of wing rotation at dorsal and ventral reversal,
(iv) the duration of the stroke reversal, (v) the shape of the wing
tip trajectory (‘oval’ or ‘figure-of-eight’) and (vi) the angular
deviation from the mean stroke plane during the upstroke
and downstroke (Fig. 1C). In most of the experiments, the
deviation amplitude was set to zero, such that the wing tip
remained within the stroke plane throughout the cycle. Under
these conditions, the kinematics of the wing stroke were
symmetrical such that the upstroke and downstroke were
mirror images of one another. Only in trials using ‘oval’ stroke
deviations were the kinematics of the two strokes not identical.
The frequency of the wing stroke (0.17 Hz) remained constant
in all experiments, as did the upstroke-to-downstroke duration
ratio, which was fixed at 1. We constructed the kinematic
patterns using a custom-designed MATLAB program
(Mathworks) to convert the angular trajectories into a series of
stepper motor commands.
Force measurements
Signals from the two-dimensional sensor were acquired
using a National Instruments data-acquisition board (model
BNC 2090) in a PC running custom-designed software written
in MATLAB. Data were filtered on-line with an active four-
pole Bessel filter with a cut-off frequency of 10 Hz and off-line
with a zero-phase-delay low-pass digital Butterworth filter
with a cut-off frequency of 3 Hz, which was 17.6 times the
wing stroke frequency. Apart from increasing the high-
frequency components resulting from motor jitter, increasing
the cut-off frequency of the filter did not alter the time course
of the force traces.
Each experiment consisted of one burst of four consecutive
wing strokes following pre-programmed kinematics. The wing
begins the first downstroke in still fluid, whereas during the
subsequent strokes it moves through a wake created by the
preceding strokes. As a result, the time course of forces
generated during the first stroke is markedly different from
those of subsequent strokes. For this reason, the data from the
first stroke were excluded from this analysis, while those from
the three subsequent strokes were averaged. Thus, each
presented trace represents an average of three force records.
After subtracting gravitational forces, the forces measured
from the normal and parallel channels were transformed into
lift, drag, thrust and radial components.
Added mass
The measured force at the wing base consists of
gravitational, inertial and aerodynamic components. The
gravitational contribution of the sensor and wing mass to the
total force signal was easily calculated and subtracted from the
measured force traces. The inertial components represent the
acceleration forces on the mass of the sensor and wing as well
as the added mass of the fluid around the wing. To examine
the contribution of the inertial effects of the wing mass and
S. P. S ANE AND M. H. D ICKINSON
Lift and drag production by a flapping wing 2611
sensor, we replaced the wing surface with an aerodynamically
neutral inertial model of the wing. The aerodynamically neutral
model was essentially a brass knob with the same mass and
center of mass inside the oil as the Plexiglas wing. Because
of its low surface area, the brass knob generated negligible
aerodynamic forces compared with the Plexiglas wing. For any
arbitrary kinematic pattern, the resulting force traces for the
brass knob could be entirely accounted for by gravity. Thus,
the inertial forces generated by flapping this brass model, and
therefore the Plexiglas wing, were negligible and have been
ignored. Compared with gravity and wing inertia, the non-
circulatory forces due to added mass are more difficult to
measure because the fluid acceleration induced by a moving
wing changes dynamically as the wing rotates, decelerates or
accelerates (Daniel, 1984).
To estimate the magnitude of added mass, we used an
approximation derived for motions of an infinitesimally thin
two-dimensional plate in an inviscid fluid (Sedov, 1965).
Using blade element method, we adapted it to the case of a
three-dimensional wing rotating around an axis located at one-
quarter chord length from the leading edge. The force
contribution normal to the wing surface due to the added mass
inertia is given by:
where ρ is the fluid density, R is the wing length, c–^ is the mean
chord length, r ˆ and cˆ ( r ˆ) are the non-dimensional radial position
along the wing and non-dimensional chord length, respectively
(for nomenclature, see Ellington, 1984a), φ is the angular
position of the wing and α is the angle of attack. Using equation
1, we calculated an estimate of added mass inertia for each set
of kinematics. As illustrated by the representative trace in
Fig. 2, the absolute contribution of added mass to net forces on
the wing was quite small in all cases. Further, by comparing
equation 1 and equation 3 (see below), it can be seen that the
added mass forces (≈ρ R^2 c–^2 ) scale in proportion to aerodynamic
forces (≈ρ R^3 c– ) for geometrically similar wings. Thus, for
identical kinematics and geometry, added mass will have the
same physical effect on a model wing as on the wing of a fly,
provided that the Reynolds number is the same. Because both
added mass and aerodynamic contributions are biologically
relevant, we chose not to subtract the inertial estimates from the
presented force traces.
Experimental procedures
To examine the influence of the six kinematic variables on
aerodynamic forces, we divided our analysis into three sets
of experiments. First, we held the values of flip timing
(symmetrical around stroke reversal), flip duration (16 % of
stroke cycle period) and stroke deviation (0 °) constant, while
we systematically varied stroke amplitude and angle of attack.
For each amplitude from 60 to 180 ° in 20 ° increments, we
varied the angle of attack at mid-stroke from 0 to 90 ° in steps
of 10 °. From the results of this 7×10 set of experiments, we
determined the combination of stroke amplitude and angle of
attack that generated maximum mean lift. Using these locally
optimizing values of stroke amplitude and angle of attack, we
systematically varied the values of flip start and flip duration.
Values for flip start indicate when in the stroke cycle the wing
begins to rotate relative to stroke reversal and are expressed as
a fraction of total cycle time (Fig. 1C). Thus, a value of −0.
indicates that the wing begins rotation 50 % of a stroke period
prior to stroke reversal, whereas a flip start value of 0 indicates
that the wing begins rotation at the instant of stroke reversal.
Values of flip duration, the total time it takes the wing to
complete wing rotation, are also represented as a fraction of
total cycle time. Thus, a value of 0.2 indicates that a flip
requires 20 % of the stroke cycle to complete. In these
experiments, we set the flip to start at various points within the
stroke from −0.5 to 0 in steps of 0.05. For each value of flip
start, we varied the flip duration from 0.1 to –0.5, also in steps
of 0.05. Flip timing, τf, which describes when the mid-point of
a flip occurs within the stroke, may be calculated from:
τf = τ 0 + 0.5∆τ , (2)
where τ 0 is flip start and ∆τ is flip duration. As before, we
1
0
rˆcˆ^2 ( rˆ )d rˆ −
F a,N = ρ R^2 c–^2 (φ¨ sinα + φ¨^ α˙ cosα)
α¨ρ c–^3 R
1
0
cˆ^2 ( rˆ )d rˆ ,
Normal force (N)
Added mass inertia
Added mass inertia subtracted
Added mass inertia included
Stroke cycle
Fig. 2. Contribution of added mass inertia to total measured aerodynamic force. The traces shown were taken from a representative kinematic pattern (stroke amplitude Φ=180 °, maximum stroke deviation Θ=0, angle of attack α=45 °, flip start, τ 0 =0.05, flip duration ∆τ=−0.1). The blue trace represents the total normal force measured on the wing. The static gravitational component has been subtracted. The black trace represents the added mass inertia estimated using equation 1. Added mass inertia is zero throughout most of the stroke because the linear velocity of the wing is constant. The red trace represents the total measured force after subtracting added mass inertia. The contribution of added mass inertia to the measured aerodynamic forces is small, as indicated by the similarity of the red and blue traces.
Lift and drag production by a flapping wing 2613
Fig. 3. Sample instantaneous forces for various combinations of total stroke amplitude Φ and mid-stroke angle of attack α. In each case, the wing rotation parameters were kept constant (flip duration ∆τ=0.16, flip start τ 0 =−0.08, flip timing τf=0). Each panel (A–H) shows a plot of measured drag (solid red line), the quasi-steady estimate of drag (broken red line), measured lift (solid blue line) and the quasi- steady lift (broken blue line). Since the radial forces were zero for all these kinematic patterns, they are not plotted. A two- dimensional diagram of the wing kinematics is plotted above each set of traces using the convention described in Fig. 1C. The wing chord is shown in light blue, and the superimposed black vector indicates the magnitude and direction of the instantaneous aerodynamic force. For convenience, the kinematic values of stroke amplitude and angle of attack used in each trial are printed in the upper left of each panel. Values for the measured mean force coefficients ( C – D^ and C – L^ ) are printed adjacent to each set of traces. Axis labels given in A apply to all panels. (A,B) Forces generated at a 90 ° angle of attack with no wing rotation (α=90 °) for a short stroke amplitude (A, Φ=60 °) and a long stroke amplitude (B, Φ=180 °). Note the enormous transients in drag at the start of each stroke due to wake capture. (C,D) Forces generated at a 50 ° angle of attack for a short stroke amplitude (C, Φ=60 °) and a long stroke amplitude (D, Φ=180 °). The contribution of rotational circulation is apparent at the end of each stroke. (E,F) Forces generated at a 30 ° angle of attack for a short stroke amplitude (D, Φ=60 °) and a long stroke amplitude (F, Φ=180 °). (G,H) Forces generated at a 0 ° angle of attack for a short stroke amplitude (G, Φ=60 °) and a long stroke amplitude (H, Φ=180 °).
α = 50 °
C D = 6. 19
C^ —L = 0. 01
A
0. 5 C
L =^1.^39
C^ —D = 4. 12
C
C^ —D = 3. 16
C^ —L = 1. 87
D
B
C^ —D = 4. 57
C^ —L = 0. 03
E
C^ —D = 3. 17
C^ —L = 0. 61
F
C L= 1. 46
C D = 1. 93
C L = - 0. 73
C^ —D = 3. 03
G
D
rag
(N)
L
ift
(N)
C^ —L = 0. 20
C^ —D = 1. 14
Downstroke Upstroke
H
α = 90 °
α = 90 °
α = 50 °
α = 30 °
α = 30 °
α = 0°
α = 0°
Stroke cycle
Stroke cycle
D
rag
(N)
Lift
(N)
D
rag
(N)
L
ift
(N)
D
rag
(N)
Lift
(N)
2614
The measured force trajectories display prominent peaks at
the beginning and end of each translational stage that are
absent or smaller in the quasi-steady predictions. In isolating
the mechanisms responsible for these transient effects, it is
useful to consider cases in which there is no wing rotation,
because this removes the potential contribution of rotational
circulation. For example, in Fig. 3A,B, the wing translates
back and forth at a 90 ° angle of attack without rotating. At the
start of each stroke, measured drag far exceeds the quasi-steady
predictions. The fact that there is no corresponding dip in drag
at the end of translation indicates that the prominent drag
transient is not due to acceleration of inertial mass. This
interpretation is consistent with estimates of added mass inertia
using equation 1 (data not shown). Instead, the elevation in
drag at the start of each stroke in Fig. 3A,B is probably due to
wake capture as the wing intercepts the fluid flow field induced
by the shed vorticity of the previous stroke.
The influence of rotational circulation is most easily seen
when the angle of attack during the upstroke and downstroke
is zero (Fig. 3G,H). Under these conditions, the vorticity
generated during the translational phase of the stroke is
minimal and, thus, the magnitude of the wake capture effect
should be small. The influence of the wake is not entirely
absent, however, because the process of rotation generates and
sheds vorticity through which the wing must translate at the
start of each stroke. Further, the vorticity created by rotation is
particularly strong when the translational angle of attack is
zero, because the wing must flip over by 180 ° during stroke
reversal, making the angular velocity of the wing particularly
large. At the end of the each stroke in Fig. 3G,H, the influence
of rotational circulation is manifest as a transient increase in
lift and drag that exceeds the quasi-steady prediction. After
stroke reversal, the continuing rotation of the wing generates
a pressure force with opposite polarity, resulting in negative
lift. The time course of this rotational effect is complicated by
the presence of an added mass inertia and a modest amount of
wake capture at the start of each stroke. The influence of these
multiple mechanisms is manifest by the positive peak in lift
immediately following stroke reversal due to wake capture,
which is followed by the negative peak due to rotational
circulation. The pattern of an early positive peak in lift
followed by a later negative peak is seen throughout the traces
in Fig. 3.
The rest of the traces in Fig. 3 illustrate the complex
interactions among delayed stall, rotational circulation and
wake capture that result from changes in angle of attack and
stroke amplitude. At angles of attack of 30 and 50 °, the wing
generates lift throughout the stroke due to delayed stall
(Fig. 3C–F). The influence of rotational lift is reduced as the
angle of attack increases, however, because the wing flips over
a smaller arc with lower angular velocity. This effect can be
seen by comparing the relative magnitude of the force peaks
at the end of each stroke in Fig. 3B,D,F,H. In contrast, the
influence of wake capture is greater at higher angles of attack
because the vorticity shed into the wake at the end of the stroke
is stronger. This effect can be seen by comparing the relative
size of the force transients at the start of each stroke in the same
panels. Thus, changing stroke amplitude and angle of attack
has a complex but interpretable influence on the magnitude of
the different unsteady mechanisms. The kinematics that
optimize the aerodynamic performance of the wing will reflect
these complex interactions. The maximum mean lift-to-drag
ratio (0.8) occurred at an angle of attack of 30 ° and amplitude
S. P. S ANE AND M. H. D ICKINSON
Angle of attack (degrees)
Stroke a
mp
lit
ude (degrees)
Net aerodynamic force (N)
A
Angle of attack (degrees)
Stroke a
mp
lit
ude (degrees)
Net force coefficient
0 6.^5
B
Fig. 4. Parameter maps of net aerodynamic force and net force coefficient as functions of stroke amplitude and mid-stroke angle of attack. For fixed values of wing rotation (flip duration ∆τ=0.16; flip start τ0=−0.08, flip timing τf=0), stroke amplitude was varied from 60 to 180 ° and angle of attack was varied from 0 to 90 °. In each diagram, the small open circles indicate the positions of actual measurements. Values between these measured points have been interpolated using a cubic spline. Values are encoded in pseudocolor according to the scales shown beneath each plot. This same format is used in Fig. 5, Fig. 7 and Fig. 10. (A) Net aerodynamic force, the vector sum of lift and drag, increases monotonically with increasing angle of attack and stroke amplitude. (B) Net aerodynamic force coefficient increases with angle of attack, but decreases with stroke amplitude.
2616
amplitude. The maximum value of C L
/ C D
(0.8) occurred at
stroke amplitude of 180 ° and an angle of attack of 30 °. The
corresponding quasi-steady-state estimates of C L
/ C D
are
independent of stroke amplitude, with a maximum of 1.1 at
an angle of attack of 20 °. Thus, the quasi-steady model
significantly overestimates aerodynamic efficiency and fails to
account for its dependence on stroke amplitude.
The effects of flip timing and flip duration
In all subsequent experiments, we set the stroke amplitude
to 180 ° and the mid-stroke angle of attack to 45 °, which are
the values that maximized lift production in the first set of
experiments. Next, we systematically varied the timing and
duration of wing rotation to examine their effects on force
production. Sample force traces selected from 99 pairs of flip
start and flip duration are shown in Fig. 6A–H, with the
corresponding values of flip timing, τf, given in the upper left
corner of each panel. A comparison of Fig. 6A,C,D illustrates
that a long-duration flip, ∆τ=0.5, can produce quite different
forces depending on when the flip occurs. If the flip occurs
symmetrically about the stroke reversal (τf=0, Fig. 6A), C L
is
quite large ( C L
=1.54) and the time course is well approximated
by the quasi-steady predictions. An advanced flip (τf=−0.25,
Fig. 6C) results in very low mean lift ( C L
=0.36), but produces
a fairly prominent wake capture peak. In contrast, a rotation
after stroke reversal (τf =+0.25, Fig. 6D) results in mean
negative lift ( C L
=−0.28) because of the adverse effects of
rotational circulation following stroke reversal.
Fig. 6 F–H shows the effects of a fast flip (∆τ=0.1) at
different flip times. As flip duration decreases, the aerodynamic
performance of the wing generally rises. Symmetrical and
advanced flips yield nearly identical mean lift ( C L
symmetrical; C L
=1.87, advanced), whereas a delayed flip
generates somewhat lower lift ( C L
=1.52). These differences
are due primarily to the amount of lift produced at the start of
each stroke. Early and symmetrical flips (Fig. 6F,G) result in
a substantial wake capture peak at the start of translation.
However, if the flip occurs very early in the stroke (τf=−0.45,
Fig. 6E), the wing translates through most of the stroke at
negative angles of attack, leading to a large value of negative
lift ( C L
=−1.41; Fig. 6E). When rotation is delayed, the wake
capture peak is missing, revealing two negative peaks at the
start of translation, an early small peak due to added mass
inertia and a later more prominent peak due to rotational
circulation (Fig. 6H).
The maps of mean force coefficients as a function of flip
duration and flip start are shown in Fig. 7A,C,E, with
comparable quasi-steady, translational predictions shown in
Fig. 7B,D,F. Flip timing, the non-dimensional time when the
mid-point of the flip occurs, is indicated by the inclined parallel
lines on each graph. Both C L
and C D
are strongly influenced
by flip timing and duration. For example, at a flip duration of
0.1, C L
varies with flip timing from as low as −1.5 to as high
as +2. The comparable values of the quasi-steady translational
estimate, C L,t
—
, also vary, but over a smaller range (from −1 to
For all values of flip timing, C D
increases monotonically
with flip duration (Fig. 7C). Measured values range from 2.
and 4.1, representing a somewhat smaller variation than was
seen with C L
. However, unlike the case with lift, the
discrepancy between measured C D
and the quasi-steady
estimate, C D,t
—
, is substantial. In addition to generally
underestimating the magnitude of drag, the quasi-steady
predictions fail to observe the local rise in drag along the −0.
flip timing iso-line. Since the range of variation for drag is less
than that for lift, the measured lift-to-drag ratio map resembles
the lift coefficient map (Fig. 7A,C,E). Further, because the
quasi-steady translational predictions underestimate lift and
drag by approximately the same proportion, the predicted lift-
to-drag ratio map is quite similar to the measured map
(Fig. 7E,F). The map for the net force coefficient (Fig. 7G)
resembles the drag map (Fig. 7C), which is expected since the
values for the mean drag coefficient are, on average, twice
those for the lift coefficient at comparable points on the
kinematic surface.
The effects of stroke plane deviation
Using kinematic values for stroke amplitude, angle of attack,
flip duration and flip start that maximized lift production
(Φ=180 °, α=45 °, ∆τ=0.1 and τf=−0.05), we tested how forces
vary with deviation from the mean stroke plane in a set of 22
experiments. The peak-to-peak magnitude of stroke deviation
was varied from −50 to +50 ° in 10 ° steps for both the half-
sine (‘oval’) and full-sine (‘figure-of-eight’) patterns. It is
worth noting that, in the oval pattern, an upward deviation at
the start of the downstroke requires a downward deviation at
the start of the upstroke and vice versa. This is not the case for
the figure-of-eight pattern, in which the two half-strokes are
mirror images of one another.
Fig. 8 shows a selection of force traces resulting from
different patterns of stroke deviation. In general, the figure-of-
eight pattern had a more profound influence on the magnitude
and time course of force production than did the oval pattern.
In both cases, however, the direction of stroke deviation at the
start of each translational phase greatly influenced the
magnitude of the force transient at the start of the stroke. For
example, in the figure-of-eight pattern shown in Fig. 8C, each
stroke begins with an upward motion, and the lift and drag
transients at the start of each stroke are quite small. In contrast,
the comparable kinematic pattern that starts with a downward
motion (Fig. 8D) generates sizeable force peaks. A similar
trend is seen in the oval patterns (Fig. 8A,B). The upstroke in
Fig. 8A and the downstroke in Fig. 8B, which both start with
a downward motion, are marked by sizeable force peaks at the
start of translation, whereas the strokes that begin with upward
motion are not. This dependence of the early force transient on
the direction of deviation is explained in part by an increase in
the aerodynamic angle of attack caused by the downward
motion of the wing. However, the measured force peaks are
much greater than the quasi-steady estimates, which take into
account this effect, suggesting that there is a substantial wake
effect at the start of each stroke. The influence of the wake is
S. P. S ANE AND M. H. D ICKINSON
Lift and drag production by a flapping wing 2617
Fig. 6. Sample instantaneous forces for various combinations of flip start τ 0 and flip duration ∆τ. In all kinematic patterns, stroke amplitude was 180 ° and angle of attack was 45 °. The format for each panel is that described for Fig.
- As in Fig. 3, the radial forces for all these kinematics are zero and have not been plotted. (A) Forces generated with a slow flip (∆τ=0.5), symmetrical with respect to stroke reversal (τ0=−0.25, flip timing τf=0). Under these conditions, the quasi-steady model (broken lines) accurately predicts measured lift, but not drag. (B) Forces generated with moderate flip duration (∆τ=0.25), advanced with respect to stroke reversal (τ0=−0.25, τf=−0.125). With these kinematics, the augmentation of lift by rotational circulation and wake capture is evident. (C) Forces generated with a long, advanced flip (∆τ=0.5; τ0=−0.5, τf=−0.25). This pattern of kinematics produced elevated drag due to wake capture at the start of each stroke. (D) Same kinematics as in C, but with a delayed flip (∆τ=0.5; τ0=0, τf=+0.25). The delay in flip timing causes a small decrease in mean drag, but an enormous decrease in lift. (E–H) The influence of rotational timing on a short- duration flip. (E) Forces generated by a short flip advanced by almost a full half-cycle with respect to stroke reversal (∆τ=0.1; τ0=−0.5, τf=−0.45). Note that the angle of attack is negative during most of translation because the wing flips much too soon. As a consequence, the pattern generates negative lift. (F) Forces generated by a slightly advanced short flip (∆τ=0.1; τ0=−0.1, τf=−0.05). This near- optimal pattern augments lift by both rotational mechanisms. (G) Forces generated by a short symmetrical flip (∆τ=0.1; τ0=−0.05, τf=0). (H) Forces generated by a slightly delayed short flip (∆τ=0.1; τ0=0, τf=0.05). The small delay of 0.05 decreases the mean lift coefficient by 20 % compared with the symmetrical case shown in G. C – D^ , mean drag coefficient; C – L^ , mean lift coefficient.
1 C
L =^1.^47
C D = 3. 28
C L = -0. 28
C D = 3. 87
1 C
L =^1.^87
C D = 2. 85
C L = 1. 9
C D = 2. 66
C D = 2. 66
C L = 1 .5 2
C L = 1 .5 4
C D = 3. 82
A
0.5 C
L =^ -^1.^41
C D = 2. 64
E
G
B
D
F
H
1 C
L =^ 0.^36
C D = 4.0 6
C
τ 0 = – 0. 25 ∆τ = 0. τf = 0
τ 0 = – 0. 25 ∆τ = 0. 25 τf = – 0. 125
τ 0 = – 0. ∆τ = 0. τf = – 0. 25
τ 0 = 0 ∆τ = 0. τf = +0. 25
τ 0 = – 0. ∆τ = 0. 1 τf = – 0. 45
τ 0 = – 0. 1 ∆τ = 0. 1 τf = – 0.
τ 0 = – 0. ∆τ = 0. 1 τf = 0
τ 0 = 0 ∆τ = 0. 1 τf = 0.
Downstroke Upstroke 0 0.5 1
Li
ft^
(N)
D
rag
(N)
Li
ft^
(N)
D
rag
(N)
Li
ft^
(N)
D
rag
(N)
L
ift
(N)
D
rag
(N)
Stroke cycle
Stroke cycle
Lift and drag production by a flapping wing 2619
Fig. 8. Sample instantaneous forces for various combinations of stroke deviation and tip trajectory. All other kinematic variables were constant (amplitude Φ=180 °, angle of attack α=45 °, flip duration ∆τ=0.5, flip timing τf=−0.05). The format for each panel is that described for Fig. 3 and Fig. 6, except for an additional panel showing the instantaneous radial forces (green). (A) Forces generated with a large oval deviation in which the downstroke starts with upward motion and the upstroke starts with downward motion D dev =+25 °, U dev=−25 °, where D dev is equal to the maximum angle of downstroke deviation and U dev indicates maximum angle of upstroke deviation. Both lift and drag transients are higher at the start of the upstroke when the wing travels downwards than at the start of the downstroke. The absolute average radial forces are also correspondingly large. (B) Reversed condition compared with A, the downstroke starts with downward motion and the upstroke starts with upward motion ( D dev=−25 °, U dev=+25 °). Lift and drag transients are now much larger at the start of the downstroke. (C) Forces generated with a figure-of- eight deviation in which both strokes start with upward motion ( U dev= D dev =+25 °). Both lift and drag are low at the start of each stroke, but reach elevated values at midstroke. (D) Reversed condition compared with C, both strokes start with downward motion ( U dev= D dev=−25 °). Both strokes now start with large transients in both lift and drag. (E) Forces generated by comparable kinematic pattern to those in A–D but with no stroke deviation.
D dev =–25° U dev =+ 25 °
C L = 1.
C D = 2 .4 7
D dev=+ 25 ° U dev=+ 25 °
C L = 1. 34
C D = 2. 23
D dev=–25° U dev=–25°
C L = 1. 27
C D = 2 .1 8
D dev =+ 25 ° U dev =–25°
C L = 1.6 2
C D = 2 .6 8
L
ift
(N)
C L = 1. 88
C D = 2. 88
D dev = 0 ° U dev = 0 °
D
rag
(N)
A
B
C
D
E
Downstroke Upstroke 0 0.^5 Stroke cycle
R
ad
ia
l^ (N)^ | C R|^ =^0.^38
| C R| = 0
| C R| = 0 .4 8
| C R| = 0. 37
| C R| = 0. 53
L
ift
(N)
D
rag
(N)
R
ad
ia
l^ (N)
Lift
(N)
D
rag
(N)
R
ad
ia
l^ (N)
2620
during the upstroke are equal and opposite to the radial forces
during the downstroke. In the ‘figure-of-eight’ kinematics,
however, the upstroke and downstroke are identical and, hence,
the radial forces during the upstroke are identical to the radial
forces during the downstroke.
Fig. 9 summarizes the effects of stroke deviation on the
time-averaged measured forces and quasi-steady predictions.
While the influence of stroke deviation on the time course of
the aerodynamic forces is large, its impact on the mean force
coefficients is surprisingly small. This indicates that the
differences in the dynamics of force production noted in Fig. 8
tend to average out over the stroke. For both the ‘oval’ and
‘figure-of-eight’ deviation trajectories (Fig. 9A), the mean lift
and drag coefficients decreased with increasing absolute
deviation (Fig. 9C,D). The changes in average performance for
the ‘oval’ deviation pattern should be symmetrical around zero
deviation, since oval patterns with positive and negative
deviations are mirror images of one another. Thus, the
downstroke in Fig. 8A should resemble the upstroke in
Fig. 8B, and the upstroke in Fig. 8A should resemble the
downstroke in Fig. 8B. The asymmetry in these measurements
results from the mechanical play in the gear mechanism of the
robot. However, the asymmetry in the performance of the
‘figure-of-eight’ patterns around zero deviation represents, in
part, a real aerodynamic effect (Fig. 9B,C,D). In this case, a
positive deviation will result in a downward motion at the start
of both the upstroke and downstroke, whereas a negative
deviation indicates upward motion at the beginning of both
strokes. Downward deviation should enhance wake capture, as
described above. Values of C L
and C D
fall off faster with
S. P. S ANE AND M. H. D ICKINSON
E
Net force coefficient
0
D
Lift coeffiecient
L
ift
/d
rag
ra
tio
D
rag
coefficient
A
B
Net deviation (degrees)
Positive deviation
Negative deviation
C
Oval
Figure-of- eight
Net deviation (degrees)
Net deviation (degrees)
Net deviation (degrees)
Downstroke
Upstroke
R
ad
ia
l/d
rag
ra
tio
Fig. 9. Effects of stroke deviation on mean force coefficients. (A) Cartoon illustrating the kinematic patterns used to study stroke deviation. Throughout the figure, blue data points refer to oval kinematic patterns, and red data points refer to figure-of-eight patterns. Measured and quasi- steady predicted values are given by filled and open circles, respectively. (B) The magnitude of the mean net aerodynamic force coefficient is only mildly influenced by stroke deviation. (C) Measured mean lift coefficient decreases with stroke deviation. For the figure-of-eight patterns, the drop in performance with increasing deviation is greater for strokes that begin with upward motion. (D) Drag coefficient decreases with stroke deviation. Note the large discrepancy between measured values of mean drag and quasi-steady predictions. (E) Ratio of lift-to-drag and radial- to-drag forces as a function of stroke deviation. Radial-to-drag forces for oval kinematic patterns are represented by filled black circles and for figure-of-eight patterns by open black circles. Because deviation affects lift and drag almost equally, the influence on their ratio is quite small. The radial forces are dependent on the sine of stroke deviation angle. Because of the linearity of sine functions for small angles, the values of radial-to-drag force increase linearly with increasing absolute deviation.
2622
0.37< C D,t <3.47). At present, the reason for this discrepancy
is not clear.
The relative importance of unsteady mechanisms
The relative contributions of delayed stall, rotational
circulation and wake capture to total force production vary
with the precise kinematics of the stroke. In general, the
importance of delayed stall increases with stroke amplitude
because the wing can integrate the influence of the leading
edge vortex over a greater distance. In contrast, rotational
S. P. S ANE AND M. H. D ICKINSON
Angle of attack (degrees)
Stroke a
mp
lit
ude(degrees)
D
Non-dimensional flip duration
N
on
im
ens
ional fl
ip
start
E
Net deviation (degrees)
C
N
on
im
ens
ional fl
ip
timi
ng
0 0.^3
Net deviation (degrees)
Angle of attack (degrees)
Stroke a
mp
lit
ude(degrees)
A
Non-dimensional flip duration
N
on
im
ens
ional fl
ip
start
B
N
on
im
ens
ional fl
ip
timi
ng
P pro (kg m^2 s–^3 )
F
L / P pro (s m-^1 )
___ _^ ___
L^
/^ P
pro
(s
m
-^1
)^
___
P pro _
(kg
m
2 s
-^3
)^
___
Fig. 10
Lift and drag production by a flapping wing 2623
effects become more important with lower stroke amplitude.
For kinematic patterns mimicking those of hoverflies with a
60 ° stroke amplitude, rotational effects account for more than
half the total force (Dickinson et al., 1999). Because of these
differences, quasi-steady translational estimates should more
closely resemble the measured values for kinematic patterns
with large stroke amplitudes and deviate from measured values
for those with lower stroke amplitudes. These predictions are
borne out by the force traces shown in Fig. 3 and the maps of
C L
in Fig. 5A,B. The quasi-steady translational model predicts
maximum lift at an angle of attack of 45 °. While this is close
to the maximum value for the measured forces at high stroke
amplitude, the measured lift maximum is shifted to higher
angles of attack (60 °) at low stroke amplitude.
There are two possible explanations for this shift. First,
rotational circulation might make a greater contribution to
mean lift at low stroke amplitude. However, this is unlikely to
be true for symmetrical flips, for which rotational circulation
enhances lift only before stroke reversal, but attenuates it after
stroke reversal (Dickinson et al., 1999). Also, at stroke
reversal, the wing rotates less for a 60 ° angle of attack than
it does for a 45 ° angle of attack. Thus, the proportional
contribution due to rotational lift is further minimized. Second,
the shift in the lift maximum may reflect an increasing
importance of wake capture at low stroke amplitude. The large
contribution of wake capture can be easily seen in the forces
generated by hoverfly-like patterns (Dickinson et al., 1999) as
well as in the drag traces in Fig. 3. The increasing importance
of wake capture also explains the changes in drag coefficients
at low stroke amplitude. While the translational quasi-steady
model predicts that drag should be independent of amplitude
(Fig. 5D), the measured mean drag coefficient clearly
increases at smaller stroke amplitudes.
Separating the effects of wing rotation from wake capture
Since rotational circulation, wake capture and added mass
usually occur together during stroke reversal, it is often
difficult to separate these effects. After estimating the effect of
added mass on force traces before and after subtraction
(Fig. 2), we concluded that, at these Reynolds numbers, the
magnitude of added mass is small compared with rotational
circulation or wake capture. To isolate these two mechanisms,
it is helpful to focus on kinematic patterns in which either the
entire flip occurs prior to stroke reversal (Fig. 6B,E,F) or the
wing does not flip at all (Fig. 3A,B). In the case of an advanced
flip, the force peak that exceeds the translational estimate prior
to stroke reversal may be attributed to rotational circulation,
while the force peak after stroke reversal is due to wake
capture. In the case of no flip, the large drag peak at the start
of each stroke is due to wake capture, in which vorticity shed
from the previous stroke elevates force by inducing an increase
in flow velocity towards the wing (Dickinson, 1994; Dickinson
et al., 1999). Because of the squared dependence of forces on
relative velocity, even small changes in flow can cause a large
elevation in force.
The influence of wake capture should be reduced when the
wing translates at a 0 ° angle of attack during the previous
stroke. Under these circumstances, the wing sheds less
vorticity at the end of each stroke, since it did not create a
leading edge vortex during translation. There is some vorticity
present, however, due to rotational circulation at the end of the
stroke. Nevertheless, in these cases (Fig. 3G,H), wake capture
is reduced, which partially isolates the influence of rotational
circulation. The results indicate that a wing rotating early
enhances lift, but that if the rotation continues after stroke
reversal the force dips below the quasi-steady-state prediction
because, after changing direction, rotational circulation will act
to counter the circulation produced by translation.
By systematically varying the kinematic parameters, we
determined that lift was optimized at a 45 ° angle of attack,
180 ° stroke amplitude, 10 % flip duration and approximately
−5 % flip advance. Even subtle deviations from the optimal
combination drastically decreased lift. A 5 % delay in flip
timing relative to the optimal conditions (Fig. 6H) is less
effective in generating rotational lift and decreases the average
coefficient by as much as 25 %. Small decreases in flip duration
cause comparable changes. These results suggest that the
control of flip timing and duration provides an easy and
powerful way of modulating the forces on two wings, as is
required to alter force moments during steering. Tethered flies
advance wing rotation on the outside of a turn in response to
a turning visual stimulus (Dickinson et al., 1993). Since early
rotation enhances whereas delayed rotation attenuates lift,
early wing rotation on the outside of a turn should produce a
greater force on that side, thus creating an appropriate turning
moment in the opposite direction. While flip advance is usually
coupled with an increase in stroke amplitude, flies do possess
the ability to decouple the modulation of rotation and
amplitude (Dickinson et al., 1993). A more thorough analysis
of steering must consider the influence of wing kinematics on
force moments, which will be the topic of a separate paper (S.
P. Sane and M. H. Dickinson, in preparation)
In nearly all the experiments presented here, the kinematics
of the upstroke and downstroke were identical. Under these
Fig. 10. The effects of wing kinematics on profile power P – pro^ and the lift-to-power ratio L^ – / P – pro^. Procedures for plotting data are as described in Fig. 4, Fig. 7 and Fig. 9. The pseudocolor scale for A and B is shown below B, and the scale for D and E is shown below E. (A) Profile power as a function of stroke amplitude and angle of attack. (B) Profile power as a function of flip start and flip duration. Flip timing is shown on the right axis. Profile power varies by a factor of 2 within the parameter space, indicating that flip kinematics are important determinants of flight cost. (C) Profile power as a function of stroke deviation for oval (blue) and figure-of-eight (red) patterns. Stroke deviation has only a minor effect on profile power. (D) The ratio of mean lift to mean profile power, L^ – / P – pro^ , as a function of stroke amplitude and angle of attack. Like the mean lift coefficient (Fig. 5A), there is a single angle of attack that maximizes L^ – / P – pro^ for each value of stroke amplitude. (E) The ratio of mean lift to mean profile power as a function of flip start, flip duration and flip timing. (F) The ratio of mean lift to mean profile power as a function of stroke deviation for oval (blue) and figure-of-eight (red) deviation.
Lift and drag production by a flapping wing 2625
on a flapping wing. In the current study, this equation predicts
a C
— D,pro
—
value of approximately 0.7 for large stroke amplitudes,
which is less than 25 % of the measured values for kinematic
patterns capable of sustaining flight ( C D,t
—
≈3). One obvious
reason for this large discrepancy is that the process of wing
rotation generates large instantaneous values of drag which, even
though lasting only a small portion of the stroke, substantially
increase the mean values. However, even the drag contributed
by delayed stall during translation (≈2) is higher than that
predicted by equation 7. These large values for the mean drag
coefficient have several important implications for flight
energetics and muscle physiology. First, profile power, which
scales linearly with the mean drag coefficient, may be several
times greater than predicted in estimates using equation 7. Thus,
under many conditions, profile power, and not induced power,
may represent the dominant component in aerodynamic power.
Further, estimates of total mechanical power based solely on
induced power are likely grossly to underestimate flight costs.
The elevated values of drag also call into question previous
attempts to estimate elastic storage, which depend upon inertial
power costs being high relative to aerodynamic costs (Dickinson
and Lighton, 1995). If the ratio of aerodynamic cost to inertial
cost is high, the energetic benefits of elastic storage are greatly
reduced. Second, because previous estimates of mechanical
power may be several times too low, calculations of muscle
efficiency, based on the ratio of mechanical power to total
metabolic power, may be substantially lower than actual values.
Such estimates for the muscle efficiency of asynchronous flight
muscle range from 8 to 12 %. When corrected for the
underestimate of drag, we find that previous estimates of muscle
efficiency presented for fruit flies (Lehmann and Dickinson,
1997) should be adjusted to 20–25 %. If correct, these adjusted
values help to solve a perplexing problem in insect flight
physiology. Because Ca2+^ cycling represents a sizeable fraction
of the energetic cost in cyclic contractions, stretch-activated
muscle should accrue an energetic savings and concomitant
increase in mechanical efficiency. This prediction was not
supported, however, by the previous estimates of efficiency.
After correction with more accurate values of drag, the estimates
of mechanical efficiency in asynchronous muscle are now
consistent with the reduced cost of Ca2+^ cycling.
In addition to indicating that the mean level of drag may be
higher than previously expected, the results also show that the
value of drag is quite sensitive to subtle changes in wing
kinematics (Fig. 5C, Fig. 7C, Fig. 9D). Mapping an estimate of
profile power indicates that this sensitivity of the drag
coefficient to wing kinematics translates into a sensitivity of
flight cost as well (Fig. 10A,B,C). This result again has
consequences for estimates of flight costs and mechanical
efficiency from free-flight kinematics based solely on measures
of stroke frequency and amplitude (Ellington, 1984d). In
particular, for a given stroke amplitude, profile power varies
depending upon the timing and duration of wing rotation
(Fig. 10B). Thus, without knowledge of the precise stroke
kinematics, estimates of free-flight energetics must be viewed
with caution.
Concluding remarks
The data presented in this paper represent a comprehensive
analysis of the effects that wing kinematics have on the
generation of aerodynamic forces in flapping flight. The results
reveal a complex system in which subtle alterations in stroke
kinematics can have quite large effects on force production. In
future studies, it will be an important challenge to determine
how real insects use this sensitively to adjust the balance of
forces and force moments as they steer and maneuver through
their environment.
List of symbols
AR aspect ratio
c–^ mean chord length
cˆ ( r ˆ) non-dimensional chord length
C
— D,pro
—
mean profile drag coefficient
C D
mean drag coefficient
C D,t translational drag coefficient
C F
mean force coefficient
C L
mean lift coefficient
C L,t translational lift coefficient
C N
mean total aerodynamic force coefficient
C R
mean radial coefficient
dφˆ/d t ˆ
mean non-dimensional angular velocity of the wing
D drag
D dev maximum angle of downstroke deviation
D ( t ) instantaneous drag
F
force coefficient averaged over a stroke
F a,N added mass inertia normal to the wing surface
F N
mean total aerodynamic force
L lift
n wingbeat frequency
P pro
mean profile power
R wing length
Re Reynolds number
r ˆ non-dimensional radial position along wing length
r ˆ 22 ( S ) non-dimensional second moment of wing area
S surface area of a wing pair
t time
T stroke period
U dev maximum angle of upstroke deviation
v wing ( t ) instantaneous wing velocity
α angle of attack
α( t ) instantaneous angle of attack
∆τ flip duration
Φ total stroke amplitude
φ( t ) instantaneous stroke position
ν kinematic viscosity
θ( t ) instantaneous stroke deviation
Θ maximum stroke deviation
ρ density of fluid
τf flip timing
τ 0 flip start
This work was supported by grants from the NSF (IBN-
2626
9723424), Defense Advanced Research Projects Agency and
the Office of Naval Research (FDN00014-99-1-0892). We
wish to thank Fritz Lehmann, Claire Balint, Namrata
Gundiah, Jim Birch and two anonymous reviewers for helpful
comments.
References
Arbas, E. (1986). Control of hindlimb posture by wind-sensitive hairs and antennae during locust flight. J. Comp. Physiol. A 159 , 849–857. Bennett, L. (1970). Insect flight: lift and the rate of change of incidence. Science 167 , 177–179. Bennett, L. (1977). Clap and fling aerodynamics – an experimental evaluation. J. Exp. Biol. 69 , 261–272. Cloupeau, M., Devillers, J. F. and Devezeaux, D. (1979). Direct measurements of instantaneous lift in desert locust; comparison with Jensen’s experiments on detached wings. J. Exp. Biol. 80 , 1–15. Daniel, T. L. (1984). Unsteady aspects of aquatic locomotion. Am. Zool. 24 , 121–134. David, C. T. (1978). The relationship between body angle and flight speed in free flying Drosophila. Physiol. Ent. 3 , 191–195. Dickinson, M. H. (1994). The effects of wing rotation on unsteady aerodynamic performance at low Reynolds numbers. J. Exp. Biol. 192 , 179–206. Dickinson, M. H. and Götz, K. G. (1993). Unsteady aerodynamic performance of model wings at low Reynolds numbers. J. Exp. Biol. 174 , 45–64. Dickinson, M. H. and Götz, K. G. (1996). The wake dynamics and flight forces of the fruit fly Drosophila melanogaster. J. Exp. Biol. 199 , 2085–2104. Dickinson, M. H., Lehmann, F.-O. and Götz, K. G. (1993). The active control of wing rotation by Drosophila. J. Exp. Biol. 182 , 173–189. Dickinson, M. H., Lehmann, F.-O. and Sane, S. P. (1999). Wing rotation and the aerodynamic basis of insect flight. Science 284 , 1954–1960. Dickinson, M. H. and Lighton, J. R. B. (1995). Muscle efficiency and elastic storage in the flight motor of Drosophila. Science 128 , 87–89. Dickinson, M. and Tu, M. (1997). The function of dipteran flight muscle. Comp. Biochem. Physiol. 116 A, 223–238. Ellington, C. P. (1984a). The aerodynamics of hovering insect flight. II. Morphological parameters. Phil. Trans. R. Soc. Lond. B 305 , 17–40. Ellington, C. P. (1984b). The aerodynamics of hovering insect flight. III. Kinematics. Phil. Trans. R. Soc. Lond. B 305 , 41–78. Ellington, C. P. (1984c). The aerodynamics of hovering insect flight. IV. Aerodynamic mechanisms. Phil. Trans. R. Soc. Lond. B 305 , 79–113. Ellington, C. P. (1984d). The aerodynamics of hovering insect flight. VI. Lift and power requirements. Phil. Trans. R. Soc. Lond. B 305 , 145–181. Ellington, C., Vandenberg, C., Willmott, A. and Thomas, A. (1996). Leading-edge vortices in insect flight. Nature 384 , 626–630. Ennos, A. R. (1989). The kinematics and aerodynamics of the free flight of some Diptera. J. Exp. Biol. 142 , 49–85. Götz, K. G., Hengstenberg, B. and Biesinger, R. (1979). Optomotor control of wing beat and body posture in Drosophila. Biol. Cybernetics 35 , 101–112.
Jensen, M. (1956). Biology and physics of locust flight. III. The aerodynamics of locust flight. Phil. Trans. R. Soc. Lond. B 239 , 511–552. Lehmann, F.-O. and Dickinson, M. H. (1997). The changes in power requirements and muscle efficiency during elevated force production in the fruit fly Drosophila melanogaster. J. Exp. Biol. 200 , 1133–1143. Lehmann, F. and Dickinson, M. (1998). The control of wing kinematics and flight forces in fruit flies ( Drosophila spp.). J. Exp. Biol. 201 , 385–401. Lehmann, F. and Götz, K. (1996). Activation phase ensures kinematic efficacy in flight-steering muscles of Drosophila melanogaster. J. Comp. Physiol. A 179 , 311–322. Liu, H., Ellington, C. P., Kawachi, K., VandenBerg, C. and Willmott, A. P. (1998). A computational fluid dynamic study of hawkmoth hovering. J. Exp. Biol. 201 , 461–477. Lorez, M. (1995). Neural control of hindleg steering in flight in the locust. J. Exp. Biol. 198 , 869–875. Marden, J. H. (1987). Maximum lift production during takeoff in flying animals. J. Exp. Biol. 130 , 235–258. Maxworthy, T. (1979). Experiments on the Weis-Fogh mechanism of lift generation by insects in hovering flight. Part 1. Dynamics of the ‘fling’. J. Fluid Mech. 93 , 47–63. May, M. L. and Hoy, R. R. (1990). Leg-induced steering in flying crickets. J. Exp. Biol. 151 , 485–488. Robertson, R. and Johnson, A. (1993). Collision avoidance of flying locusts: steering torques and behaviour. J. Exp. Biol. 183 , 35–60. Ruppell, G. (1989). Kinematic analysis of symmetrical flight manoeuvres of Odonata. J. Exp. Biol. 144 , 13–43. Sedov, L. I. (1965). Two-Dimensional Problems in Hydrodynamics and Aerodynamics , pp. 20–30. New York: Interscience Publishers. Spedding, G. R. (1993). On the significance of unsteady effects in the aerodynamic performance of flying animals. Contemp. Math. 141 , 401–419. Spedding, G. R. and Maxworthy, T. (1986). The generation of circulation and lift in a rigid two-dimensional fling. J. Fluid Mech. 165 , 247–272. Tu, M. S. and Dickinson, M. H. (1994). Modulation of negative work output from a steering muscle of the blowfly Calliphora vicina. J. Exp. Biol. 192 , 207–224. Vogel, S. (1967). Flight in Drosophila. III. Aerodynamic characteristics of fly wings and wing models. J. Exp. Biol. 46 , 431–443. Wakeling, J. E. and Ellington, C. P. (1997). Dragonfly flight. III. Lift and power requirements. J. Exp. Biol. 200 , 583–600. Wang, J. (2000). Vortex shedding and frequency selection in flapping flight. J. Fluid Mech. 410 , 323–341. Wilkin, P. J. (1990). The instantaneous force on a desert locust, Schistocerca gregaria (Orthoptera: Acrididae), flying in a wing tunnel. J. Kansas Ent. Soc. 63 , 316–328. Wortmann, M. and Zarnack, W. (1993). Wing movements and lift regulation in the flight of desert locusts. J. Exp. Biol. 182 , 57–69. Zanker, J. M. (1988). How does lateral abdomen deflection contribute to flight control of Drosophila melanogaster. J. Comp. Physiol. A 162 , 581–588. Zanker, J. M. (1990a). The wing beat of Drosophila melanogaster. I. Kinematics. Phil. Trans. R. Soc. Lond. B 327 , 1–18. Zanker, J. M. (1990b). The wing beat of Drosophila melanogaster. III. Control. Phil. Trans. R. Soc. Lond. B 327 , 45–64. Zanker, J. M. and Götz, K. G. (1990). The wing beat of Drosophila melanogaster. II. Dynamics. Phil. Trans. R. Soc. Lond. B 327 , 19–44.
S. P. S ANE AND M. H. D ICKINSON