¡Descarga Microbiología y más Apuntes en PDF de Biología solo en Docsity!
TEMA 8. REACCIONES DE ABASTECIMIENTO EN HETEROTROFOS
- Las Reacciones que permiten el abastecimiento de energía y carbono a los microorganismos quimioheterotrofos: 1.1. Degradación de polímeros extracelulares. 1.2Canalización de los monómeros a las rutas del metabolismo central: rutas glucolíticas (ruta KDPG o de Entner-Doudoroff, ruta Embden-Meyerhoff y ruta PP o de las Pentosas Fosfato), ciclo de Krebs, reacciones anapleróticas y ciclo del glioxilato, cadenas de transporte electrónico aerobias y anaerobias. Las oxidaciones sin aceptor externo de electrones: fermentacies, tipos, sustratos y productos residuales. Ejemplos de fermentaciones de carbohidratos, ácidos orgánicos y aminoácidos. 1.3 Respiraciones anaerobias facultativas y obligadas: la desnitrificación, la sulfatorreducción. Diferencias con la respiración aerobia. Subproductos y consecuencias. 1.4Fotoheterotrofía. 1. Las reacciones que permiten el abastecimiento de energía y carbono a los microorganismos quimioheterótrofos.
El metabolismo puede dividirse globalmente en dos partes fundamentales. En el catabolismo, moléculas grandes y complejas son descompuestas en moléculas más pequeñas y sencillas, liberándose energía en el proceso. Parte de esta energía es atrapada y está disponible para realizar trabajo, mientras que el resto se libera en forma de calor. La energía atrapada puede utilizarse en el anabolismo , la segunda parte del metabolismo.
En este tema nos centraremos en el metabolismo de los microorganismos quimioorganoheterotrofos. Estos microorganismos utilizan un sustrato que actúa como fuente de energía y poder reductor y que, a través de procesos de oxidación (completa o parcial), generan el ATP y los equivalentes reductores necesarios. Estos componentes serán utilizados posteriormente en el resto de procesos de biosíntesis que ocurren en el organismo.
La mayoría de estos procesos están acoplados a una cadena de transporte electrónico y, según el tipo de microorganismo, presentan un aceptor de electrones diferentes que pueden ser sulfato, CO2, aceptores orgánicos, etc. Según el tipo de aceptor de electrones podemos diferenciar varias formas de obtener energía:
1. En la fermentación el sustrato energético es oxidado y degradado sin la participación de un aceptor
externo de electrones. Normalmente, estos aceptores se tratan de compuestos intermedios como el piruvato que actúa como aceptor de electrones (este tipo de aceptores se denominan interno o endógenos). Las fermentaciones se producen habitualmente en condiciones anaerobias, aunque en ocasiones el oxígeno puede estar presente. En estos casos, el ATP se forma por fosforilación a nivel de sustrato y es el tipo de obtención de energía menos rentable ya que en estos compuestos intermediarios queda mucha energía retenida debido a que tienen el mismo estado de oxidación que el nutriente original.
2. El metabolismo productor de energía también puede utilizar aceptores externos de electrones. Este
tipo metabólico se denomina respiración y puede dividirse en dos tipos. En la respiración aerobia,
el aceptor final de electrones es el oxígeno, mientras que en la respiración anaerobia es un aceptor diferente del oxígeno. La cantidad de energía libre derivada de la respiración es mayor que en la
3. fermentación. Además, la respiración aerobia es la más rentable energéticamente por lo que será la
que los microorganismos llevarán a cabo siempre que puedan. Un poco menos rentable sería la respiración anaerobia.
FASES DEL METABOLISMO
Las tres etapas del metabolismo central de los microorganismos quimioheterótrofos son: rutas glicolíticas, ciclo de Krebs y oxidaciones .En estas tres rutas se sintetizan los 12 metabolitos necesarios para su crecimiento.
El primer estadio es la de despolimerización : los nutrientes que utilizan los microorganismos no vienen en forma de monómeros para ser directamente incorporados sino que lo más normal es encontrar nutrientes en forma de macromoléculas o formas más grandes como pueden ser partículas nutritivas en suspensión o adheridas a la superficie. Por ello, aprovechar estos nutrientes es necesario hidrolizar y degradar estas macromoléculas.
Puesto que estos microorganismos no poseen el mecanismo de tipo fagocítico, la hidrólisis de estas macromoléculas supone llevar a cabo una digestión extracelular. Para ello, vierten enzimas hidrolíticas al entorno, que son las encargadas de degradar las macromoléculas (proteínas, polisacáridos y lípidos) dando lugar a sus determinados monómeros (aminoácidos, monosacáridos, nucleótidos, etc). Una vez en forma de monómeros, los micro. los incorporan por transporte activo.
Sin embargo, el incorporar los nutrientes no resulta tan sencillo ya que tienen que competir con los microorganismos de su alrededor, puesto que mucho de ellos no poseen enzimas hidrolíticas para producir la degradación de las macromoléculas y aprovechan la presencia de un microorganismo que si puede hacerlo para poder así incorporarlas. Además, en algunos casos la degradación no da lugar a monómeros sino que puede dar lugar a péptidos (previa desaminación antes de incorporarlos), disacáridos o glicerol y ácidos grasos de cadena variable, los cuales serán aún más degradados en el interior del microorganismo.
El segundo estadio es aquel en el que los monómeros se incorporan como intermediarios en alguna ruta del metabolismo. Los aminoácidos, los monosacáridos, los ácidos grasos, el glicerol y otros productos resultantes de la primera etapa son degradados a una serie de moléculas más sencillas en la
menudo, se incorporan a la vía otros azúcares mediante su conversión en glucosa 6-fosfato o fructosa 6- fosfato. En esta etapa se consumen dos moléculas de ATP por cada molécula de glucosa.
La segunda etapa comienza cuando la fructosa 1,6-bisfosfato (6C) se escinde en dos mitades, cada una de ellas con un grupo fosfato (3C). Uno de los productos, el gliceraldehído 3-fosfato , se convierte en piruvato a través de otras reacciones. El otro producto, la dihidroxiacetona fosfato , puede transformarse fácilmente en gliceraldehído 3-fosfato y llevar a cabo el mismo proceso. En primer lugar, el gliceraldehído 3- fosfato se oxida con NAD+ como aceptor de electrones, incorporándose al mismo tiempo un grupo fosfato para formar una molécula de alta energía denominada 1,3-bisfosfoglicerato. El grupo fosfato de alta energía unido al carbono 1 es cedido posteriormente al ADP para formar ATP ( no importante saber cómo ocurre el proceso solo saber que se genera ATP ).
En este caso, la síntesis de ATP recibe el nombre de fosforilación a nivel de sustrato, debido a que la fosforilación del ADP está acoplada a la degradación exergónica de una molécula de sustrato de alta energía. Un proceso similar genera el segundo ATP por fosforilación a nivel de sustrato. El grupo fosfato del 3-fosfoglicerato pasa al carbono 2, y el 2- fosfoglicerato es deshidratado para formar una segunda molécula de alta energía, el fosfoenolpiruvato. Esta molécula cede su grupo fosfato al ADP para formar un segundo ATP y piruvato , el producto final de la vía. En conjunto, la vía glucolítica degrada una molécula de glucosa en dos moléculas de piruvato mediante la secuencia de reacciones anteriormente descrita. También se producen ATP y NADH. Esta producción puede calcularse considerando las dos etapas por separado. En la etapa de seis carbonos (etapa 1), se utilizan 2 ATP para formar fructosa 1,6-bisfosfato. Por cada gliceraldehído 3-fosfato transformado en piruvato, se forman 1 NADH y 2 ATP. Debido a que a partir de una glucosa se forman 2 gliceraldehídos 3-fosfato (1 a través de la dihidroxiacetona fosfato), la etapa de tres carbonos (etapa 2) genera 4 ATP y 2 NADH por molécula de glucosa. Si restamos el ATP utilizado en la etapa de seis carbonos al ATP producido en la etapa de tres carbonos obtenemos una producción neta de 2 ATP por molécula de glucosa. Por tanto, el catabolismo de la glucosa a piruvato en la glucólisis puede representarse por la siguiente ecuación:
GLUCOSA + 2ADP + 2Pi + 2NAD+ 2PIRUVATO + 2ATP + 2NADH + 2H+ 2H
2. Vía de Entner-Doudoroff: en un número apreciable de bacterias Gram negativas esta ruta
glucolítica clásica viene sustituida por una ruta alternativa, la ruta Entner-Doudoroff (KDPG
siglas del intermediario característico, el cual realiza la misma función que en la otra ruta tiene la fructosa-1,6- difosfato). Sin embargo, se ha encontrado en muy pocas bacterias Gram positivas.
Esta vía comienza con las mismas reacciones que la vía glucolítica. En ella se produce la formación de glucosa 6-fosfato (6C) que se transforma en 6-fosfogluconato (6C). El 6-fosfogluconato se deshidrata para formar 2-ceto-3-desoxi-6-fosfogluconato o KDPG , el producto intermedio clave de esta vía. El KDPG se esciende dando piruvato (3C) y gliceraldehído 3-fosfato (3C) que, siguen las mismas transformaciones que en la segunda etapa de la vía glucolítica para producir ATP por fosforilación a nivel de sustrato. Si la vía de Entner-Doudoroff degrada la glucosa a piruvato de esta forma, produce 1 ATP, 1 NADPH y 1 NADH por molécula de glucosa metabolizada.
De esa manera, el rendimiento energético de esta ruta es justo la mitad que en la ruta clásica donde se generan 2ATP. Esto no supone un gran cambio, sin embargo en las bacterias fermentadoras que utilizan esta ruta tienen la mitad de energía que un fermentador que utilice la ruta clásica por lo que, bajo sí que le supone una cierta desventaja.
Debido a esto, existen pocas bacterias que lleven a cabo esta ruta pero, entre ellas, destacamos al género Zymomonas (fermentación alcohólica) que produce diverosos vinos en zonas tropicales.
3. Vía de las pentosas fosfato: además de estas dos rutas, también existe otra ruta presente en todas las bacterias y que se realiza en paralelo juanto a las otras dos rutas ya comentadas. Es una ruta muy complicada y compleja por lo que no entraremos en ella. por eso la señora dijo que no la daríamos en profundidad, jejeje que majica =)
La vía de las pentosas fosfato posee varias funciones catabólicas y anabólicas que se resumen de la siguiente forma:
- El NADPH procedente de la vía de las pentosas fosfato actúa como fuente de electrones para la reducción de moléculas durante la biosíntesis. El NADPH es el donador de H+ más común.
- La vía sintetiza azúcares de cuatro y cinco carbonos para diversas finalidades. El azúcar de cuatro carbonos eritrosa 4-fosfato se utiliza para sintetizar aminoácidos aromáticos y vitamina B6 (piridoxal). La ribosa 5-fosfato (pentosa) es un componente fundamental de los ácidos nucleicos, y la ribulosa 1,5- bisfosfato es el principal aceptor de CO2 en la fotosíntesis. La eritrosa 4-fosfato y la ribosa 5-fosfato son 2 metabolitos precursores y por tanto si no los incluye el medio tendrán que generarlos en esta ruta.
- Los productos intermedios de la vía de las pentosas fosfato pueden utilizarse para producir ATP. Además, parte del NADPH puede convertirse en NADH, el cual produce ATP al ser oxidado por la cadena transportadora de electrones (leer).
CICLO DE KREBS o CICLO DE LOS ÁCIDOS TRICARBOXÍLICOS:
Antes de comenzar este proceso, el piruvato sintetizado en la glucólisis se transforma en acetil-coA al pasar a la mitocondria. En este ciclo se generan los otros 3 metabolitos necesarios (oxalacetato, succinilCoA, alfa- cetoglutarato).
Este ciclo consiste en una cadena cíclica de varias reacciones catalizadas por diversas enzimas en las que se consume el acetil-coA. El ciclo puede dividirse en tres etapas basándose en el tamaño de sus productos intermedios. Las tres etapas están separadas entre sí por dos reacciones de descarboxilación (reacciones en las que se pierden grupos carboxilo en forma de CO2).
1.3 Respiraciones anaerobias facultativas y obligadas: la desnitrificación, la sulfatorreducción. Diferencias con la respiración aerobia. Subproductos y consecuencias.
Los electrones derivados de los azúcares y de otras moléculas orgánicas suelen ser cedidos a aceptores de electrones orgánicos endógenos (fermentación), o bien, al O2 mediante la cadena transporte electrónico (respiración aerobia). Sin embargo, numerosas bacterias tienen cadenas transportadoras de electrones que pueden operar con aceptores de electrones exógenos diferentes al O2. Este proceso se denomina respiración anaerobia. Los principales aceptores de electrones son el nitrato, el sulfato y el CO2, pero los metales y algunas moléculas orgánicas también pueden ser reducidos (TABLA). Podemos diferenciar entre: Respiraciones anaerobias facultativas (alternativa a la aerobia): pueden comportarse como aerobias (aquellas que utilizan O2 como aceptor de electrones, y cuyo producto reducido es el agua) pero también como anaerobias, es decir, pueden pasar a respirar con otro aceptor electrónico lo que les supone tener que cambiar la cadena de transporte electrónico. Esto les obliga a modificar su metabolismo pero los permite vivir en anaerobiosis. Como ejemplo, veremos el de las bacterias que emplean el NO 2 -^ como aceptor electrónico.
Respiraciones anaerobias obligadas: suelen utilizar sulfato o CO2 como aceptor electrónico. Solo lo llevan a cabo aquellos micro que solo pueden vivir de esta forma. Como ejemplos estudiaremos las bacterias sulfatoreductoras (emplean sulfato como aceptor) y las arqueas metanogénicas que emplean CO 2 como aceptor.
DESNITRIFICACIÓN : proceso mediante el cual algunas bacterias utilizan nitrato (NO 3 - ) como aceptor de electrones. A menudo se suele denominar reducción desasimilatoria de nitrato. Este proceso se lleva a cabo por bacterias que, en condiciones aerobias emplean el 0 2 pero que, en condiciones anaerobias, emplean el nitrato como aceptor modificando su metabolismo. El nitrato puede ser reducido a nitrito (NO 2 - ) por la nitrato reductasa , que sustituye a la citocromo oxidasa. Se trata de una enzima citoplasmática, soluble e inhibida en presencia de sales amónicas. Está regulada por la presencia de O2, si hay O2 no necesita esa nitrato reductasa ya que llevará a cabo la respiración aerobia.
Sin embargo, la reducción del nitrato a nitrito no es una forma especialmente eficaz de producir ATP, ya que se requiere una gran cantidad de nitrato para el crecimiento (una molécula de nitrato sólo aceptará dos electrones) y además, el nitrito es muy tóxico. Por ello, en la mayoría de bacterias, el nitrato se reduce aún más hasta dar N 2 (gas nitrógeno), proceso denominado desnitrificación. Cada nitrato aceptará cinco electrones, y el producto no será tóxico. En este caso, cada reacción es llevada a cabo por enzimas ligadas a
membrana o periplásmicos que reducen el nitrato en el medio hasta nitrógeno molecular (mayor variabilidad). De esta manera, por cada dos moléculas de nitrato se emplean 10 electrones y 12H+ para obtener N2 y 6 moléculas de agua.--> que se nos quede el proceso.
Cuando estos micro., se quedan expuestos al O 2 pasan a realizar un metabolismo aerobio.
SULFATORREDUCCIÓN: ocurre en un grupo de bacterias que únicamente realizan la respiración anaerobia con sulfato (anaerobios estrictos). A este tipo de bacterias se las llama sulfatorreductoras y viven de la oxidación de compuestos orgánicos, etanol, hidrógeno, etc que genera su propio metabolismo.
Utilizan el sulfato (SO 4 2-) como aceptor electrónico. Sin embargo es necesaria una previa activación de éste para que pueda actuar como aceptor de electrones (que se transforme en un primer intermediario, el APS y, de ahí, a sulfito), proceso requiere un alto nivel de energía. La APS reductasa es la encargada de reducir el sulfato a sulfito (SO 3 2-) liberando con ello AMP. A su vez, este sulfito actúa como aceptor de electrones en otras 3 reacciones reductivas hasta llegar a dar ácido sulfhídrico a través de la sulfito reductasa. Finalmente, este residuo generado se secreta al medio.
Este tipo de bacterias son tolerantes a niveles elevados de H 2 S pero puede tener graves consecuencias para el medio en el que viven ya que pueden producir precipitación si hay componentes metabólicos en el medio. Además el H 2 S es muy corrosivo y resulta tóxico para muchos micro.
- Aminoácidos (obtenidos al degradar proteínas)
En algunos casos, otras bacterias utilizan la ruta de las pentosas fosfato en su fermentación pero, en general, se suele llevar a cabo la vía glucolítica.
TIPOS DE FERMENTACIONES (Sustrato: azúcares)
En la fermentación existe un aceptor de electrones endógeno que permite la reoxidación del NADH. Cuando este sustrato es un azúcar, sigue las rutas glucolíticas dando lugar a piruvato. Las rutas fermentativas van a diferir según como actúe el receptor endógeno.
En este esquema, podemos ver que todas las rutas fermentativas parten del piruvato, pero como hemos visto no todas parten de este sustrato sino que pueden utilizar también ácidos grasos o aminoácidos. Tipos:
Fermentación homoláctica : Utiliza el piruvato para reoxidar NADH y convertirlo en lactato (ver más).
Fermentación alcohólica : convierte el piruvato en etanol, en eucariotas es la fermentación más usual, permite la obtención de bebidas alcohólicas, combustibles, etc.
Fermentación propiónica : el piruvato es convertido a propionato, acompañado de la producción de CO2. Es importante en la maduración de los quesos, el CO2 es el causante de los agujeros de algunos tipos de quesos.
Fermentación butanodiólica : con esta fermentación obtenemos como producto 2,3-butanodiol.
Fermentación ácidomixta : Este tipo de fermentación la llevan a cabo bacterias entéricas. Se obtiene como producto ácido fórmico.
Fermentación butírica: producto final butirato, se trata de una fermentación compleja. Forzando los procesos podemos llegar a formar butanol en lugar de butirato.
Existen fermentaciones que dan lugar a ácido acético, fermentación homoacética.
La mayoría de fermentaciones dan lugar a ácidos orgánicos, pero no todas ellas, algunas pueden dar lugar a productos neutros, etanol, butanol, etc. Un ejemplo curioso son las bacterias que
producen el vinagre, se trata de bacterias acéticas que utilizan etanol como sustrato y lo convierten en ácido acético. No se trata de bacterias fermentativas. Son aerobias estrictas (necesitan el CO2) con un metabolismo estrictamente respiratorio. Son oxidadores incompletos, se les acumula el acetato porque carecen del ciclo de Krebs o funciona de una forma insuficiente.
TIPOS DE FERMENTACIONES EN BASE A OTROS SUSTRATOS:
● SUSTRATO: CARBOHIDRATOS
FERMENTACIONES LÁCTICAS (Bacterias del ácido láctico, BAL). Este tipo de bacterias tienen un gran interés en la microbiología de alimentos. Incluyen un grupo grande de bacterias Gram positivas que se comportan como anaerobias tolerantes. Sus procesos energéticos derivan de la degradación de carbohidratos. Dentro de las fermentaciones lácticas podemos diferenciar tres grupos:
1. Fermentación homoláctica :
Sigue la glucólisis clásica, dando lugar, a partir de una glucosa, a 2 piruvatos los cuales van a ser convertidos a 2 moléculas de lactato. Por tanto, en este tipo de fermentación se obtiene un rendimiento energético neto de 2 moléculas de ATP y 2 moléculas de ácido láctico a partir de una sola molécula de glucosa.
2. Fermentación heteroláctica :
Utiliza una ruta para degradar los azúcares que no es la glucólisis clásica. En ella, el 6-fosfogluconato es descarboxilado al tiempo que se oxida hasta la ribulosa 5-fosfato desprendiendo CO2. Con ello se forman pentosas (xilulosa 5-fosfato), la cual va a sufrir una rotura incorporando un grupo fosfato del medio, dando lugar a dos tipos de molécula: una es el gliceraldehido 3-fosfato que va a sufrir el mismo destino que en la ruta homoláctica y, por otro lado, el Acetil-fosfato (molécula ricoenergética) que se utiliza para reoxidar el NAD+. De esta manera, se pierde el fosfato que posee de alta energía por lo que no se utiliza para crear energía. El acetil-fosfato va a pasar a convertirse en etanol.
En conclusión en este tipo de fermentación se obtiene: CO2, ácido láctico y etanol. El rendimiento neto energético es solo de una molécula de ATP procedente del gliceraldehído-3P.
3. Fermentación de bifidobacterias :
● SUTRATO: Aminoácidos
FERMENTACIÓN DE AMINOÁCIDOS (EJ: Clostridium)
Se trata de fermentaciones de parejas de aminoácidos en los que una parte se oxida y la otra se reduce. Se utilizan parejas de aminoácidos porque la oxidación de uno de ellos costa de la reducción del otro. Ejemplo: la Alanina sufre una desaminación y forma piruvato. Éste sufre una descarboxilación al mismo tiempo que se oxida el piruvato dando lugar a Acetil-coA, en ese momento entra un fosfato y se forma Acetil-P, se libera ATP gracias a ese fosfato de alta energía liberando como producto final acetato.
Alanina + 2Glicina 3acetato + CO2 + 3NH4 + 3ATP
Se trata de una reacción de Stickland en la que un aminoácido es oxidado y un segundo aminoácido actúa como aceptor de electrones.
1.5 Obtención del ATP
En general, todos los organismos dependen en último término (a pesar de que algunos obtengan la energía por fosforilación a nivel de sustrato) de una ATPasa para obtener la energía. Este complejo se encuentra en la membrana y convierte el gradiente de protones (o Na+) en energía en forma de ATP.
a) Propionigenium
Este grupo de bacterias tienen un metabolismo sencillo. Utiliza succinato presente en el medio, es capaz de incorporarlo al interior celular, una vez ahí es descarboxilado por la descarboxilasa (enzima integral de membrana) a propianato. Por cada molécula de succinato que descarboxila expulsa un Na+. Por lo
que se crea un gradiente de Na+ debido a esta descarboxilasa. Hay una Na-ATPasa que transporta el Na+ liberando ATP (gradiente iónico de Na+). Esta bacteria vive energéticamente de este proceso. Es difícil de clasificar porque no llevar a cabo ni respiración ni fermentación.
b) Oxalobacter
Este grupo de bacterias utilizan el ácido oxálico del medio, el cual penetra en el interior y es descarboxilado a ácido fórmico. El ácido fórmico es expulsado de la célula a partir de un transportador antiporte formato/oxalato. De esta manera, se crea un gradiente de H+ y lo aprovecha la ATPasa para producir ATP.Es un mecanismo muy potente por el que la bacteria puede obtener todo el ATP que necesita. No podemos clasificarlo, porque ni llevar a cabo respiración ni fermentación.
TEMA 9. REACCIONES DE ABASTECIMIENTO EN AUTOTROFOS
1.- Las reacciones de abastecimiento de carbono en autotrofos: Rutas de fijación de CO2 (Calvin-Benson, Krebs reductivo, ruta del acetil-CoA y ruta del hidroxipropionato), distribución. 2.- Las reacciones de abastecimiento de energía y poder reductor en quimiolitotrofos:
_- Sustratos oxidables, subproductos, cadenas de transporte electrónico y aceptores, transporte electrónico inverso. Metanogénesis. 3.- Las reacciones de abastecimiento de energía y poder reductor en fotolitotrofos y fotoorganotrofos oxigénicos y anoxigénicos.
- Fotosistemas. Fotofosforilación cíclica. Donadores electrónicos en fototrofos anoxigénicos. Fototrofos anoxigénicos aerobios. Fototrofía con rodopsinas procarióticas. 4.- El abastecimiento de otros elementos biogénicos:
- Nitrógeno: asimilación de amonio, nitrato y nitrógeno molecular. Fijación simbiótica de N2. Azufre y fósforo. Asimilación de hierro: sideróforos. 5.- Biosíntesis: esquema general y particularidades en las rutas biosintéticas microbianas._
Las reacciones de abastecimiento de carbono en autotrofos: Rutas de fijación de CO2 (Calvin- Benson, Krebs reductivo, ruta del acetil-CoA y ruta del hidroxipropionato), distribución.
Los organismos autótrofos son aquellos que obtienen su fuente de C a partir del carbono inorgánico totalmente oxidado procedente del CO 2 atmosférico mediante el proceso de fijación del carbono para la síntesis de los metabolitos precursores. Existen dos modalidades de autotrofía:
- Fotolitoautotrofía : obtienen la energía y el poder reductor (NADH o Ferredoxina reducida) a partir de la radiación luminosa en la fotosíntesis. Generan O 2 como residuo inorgánico oxidado.
- Quimiolitoautotrofía : obtienen energía y poder reductor a partir de compuestos inorgánicos. Generan compuestos inorgánicos aún más oxidados como azufre elemental y (SO4)- consecuencia de los diferentes compuestos metabolizados.
Los heterótrofos, en cambio, obtienen la energía necesaria y sus metabolitos precursores a partir de compuestos orgánicos en los que el C ya está reducido. Generan como residuo compuestos totalmente oxidados (CO 2 y el O 2 ) o residuos fermentativos que pueden ser reducidos por otros organismos. En este tema nos centraremos en los quimiolitoautotrofos, cuya diversidad metábolica es muy variada.
Rutas de Fijación de CO 2 en los microorganismos :
Ciclo de Calvin
Este ciclo es el más extendido en la biosfera ya que es utilizado por la totalidad de los eucariotas autótrofos y muchos procariotas. Se trata de un ciclo complejo que combina reacciones de las rutas glucolíticas típicas pero que presenta dos reacciones exclusivas de autótrofos.
Fase de carboxilación : donde el aceptor del CO 2 , la ribulosa 1,5-bifosfato se carboxila y se escinde en dos moléculas de 3-Fosfoglicerato (triosas fosfato). Esta reacción está catalizada por
Una de las tres enzimas exclusivas es una Citratoliasa, que incorpora el Coenzima A al Citrato producido a partir de una serie de reacciones desde oxalacetato. Otra enzima exclusiva de este metabolismo lleva a cabo la carboxilación del Sucinil CoA interviniendo como donador electrónico la ferredoxina oxidada. Una tercera enzima exclusiva de esta ruta metabólica de fijación del Co2 cataliza la carboxilación de Acetil CoA en piruvato interviniendo también como donador de electrones la Ferredoxina oxidada (leer).
El resto de enzimas de esta ruta, son las mismas en ambos casos pero actúan en dirección contraria, de manera reductiva.
Este ciclo lo llevan a cabo bacterias hipertermófias quimiolitoautótrofas, arqueas con ese mismo metabolismo con las que comparten hábitat; y bacterias fotolitoautótrofas anoxigénicas verdes.
Ruta de la Acetil-CoA o de la CO deshidrogenasa
Se trata de una ruta no cíclica. El enzima característico de esta ruta es la Monóxido de carbono deshidrogenasa (que le da nombre a la ruta) , capaz de unir C en forma de grupo monóxido e incorporar sobre él un grupo metilo generado a través de una serie de reacciones, metanogénicas o no, desde otro Co2. En este proceso se consume una gran cantidad de energía y poder reductor. A partir de este producto se sintetiza Acetil CoA, la primera molécula con carbono orgánico de la ruta y se forma piruvato al incorporar una tercera molécula de Co2.
Los microorganismos que presentan esta ruta metabólica son: arqueas metanógenas, la ruta forma parte del propio proceso de metanogénesis, bacterias sulfatorreductoras quimiolitoautótrofas, bacterias acetógenas (algunas heterótrofas), bacterias del grupo Anammox , oxidadoras de amonio.
Ruta del hidroxipropionato
Se trata de una ruta en la que se producen incorporaciones sucesivas de moléculas de Co2 con gasto energético para la síntesis de Glicoxilato como forma de C orgánico, molécula con 2 carbonos reducidos. Este ciclo solo lo lleva a cabo un único grupo de procariotas (bacterias fotosintéticas verdes no dependientes de azufre. Como vemos en su rendimiento, también se requiere poder reductor y energía en forma de ATP.
Ciclo del 3 hidroxipropionato/ hidroxibutirato
Hasta hace poco, solo se conocían estas 4 formas de incorporar el Co2. Pero, desde hace poco, se ha descurbierto otra forma de fijar el Co2 en arqueas. (no hablaremos de ella).
Las reacciones de abastecimiento de energía y poder reductor en quimiolitotrofos:
.1. Sustratos oxidables, subproductos, cadenas de transporte electrónico y aceptores, transporte electrónico inverso.
Estos organismos obtienen energía y poder reductor de cualquier compuesto inorgánico reducido susceptible de oxidación. Entre los sustratos oxidables más importantes encontramos:
Compuestos reducidos del N (NH3,NH4,NO2) son metabolizados por bacterias nitrificantes aerobias y anaerobias oxidadoras de nitrito y amonio. También las del grupo anammox (anaerobias estrictas). Utilizan como aceptores electrónicos oxígeno molecular (aerobias) y nitrito, produciendo residuos de nitrógeno oxidados. Azufre elemental y compuestos reducidos del azufre como tiosulfato o sulfhídrico, oxidados por bacterias del azufre en condiciones anaerobias y aerobias. El oxígeno molecular y el nitrito son también los aceptores electrónicos en este caso. Hierro ferroso, es metabolizado en condiciones aerobias por bacterias y en condiciones de anaerobiosis por arqueas. H2, es el sustrato más demandado ya que presenta una gran cantidad de aceptores además del O2. Por ello, multitud de grupos de procariotas compiten por él. Esto se debe a que la reacción de oxidación del H2 es la que produce más energía de entre las de oxidación de compuestos inorgánicos. Es metabolizado tanto en condiciones de hipertermofília como de mesofília así como en anaerobiosis y aerobiosis. Cuando se oxida con Co2 ( metanogénesis) se convierte en un reservorio de C, causante del 100% del metano terrestre actual que proviene de origen biológico. CO, el monóxido de carbono solo es metabolizado por carboxidobacterias y en condiciones aerobias ya que solo el O2 puede actuar como aceptor de electrones.
La mayoría de las reacciones de estos micro. son exergónicas y ocurren de manera espontánea. Sin embargo, su rendimiento energético es mucho menor que el de los quimiolitoheterótrofos ya que la oxidación del O2 da mucha más energía (bajo rendimiento químico).
La producción de nitrito como residuo produce la acidificación del medio donde se desarrollan estos microorganismos y dado que son sensibles o poco tolerantes a condiciones ácidas deben crecer en un ambiente abierto que presente renovación continua para no verse inhibidas en cuanto a crecimiento por los residuos de su propio metabolismo.
Las de tipo II presentan un metabolismo más sencillo en el que solo interviene una única enzima: la nitrito oxidoreductasa (NOR) una enzima integral de membrana que produce la oxidación de nitrito a nitrato con cesión de e- que se emplean en el bombeo de protones para generar un gradiente y que la ATP sintasa pueda producir ATP (no hay que olvidar que parte de ese gradiente de protones se utilizara en vez de sintetizar ATP por medio de la ATPsintasa para obtener poder reductor a través de la cadena de transporte electrónico inverso).
En condiciones de anaerobiosis estrictas encontramos un grupo de bacterias capaces de oxidar el amonio con nitrito como aceptor de e- que integran al grupo Anammox. Son difíciles de estudiar ya que es difícil reproducir las condiciones del medio donde se desarrollan. En su interior presentan una gran vacuola central llamada anammoxoma cuya membrana está formada por unos lípidos especiales, los laderanos, que le otorgan una gran de impermeabilidad a la membrana que solo puede ser atravesada mediante enzimas específicas. Sirve, principalmente, para almacenar la hidracina, un intermediario oxidativo muy tóxico que queda retenido aquí.
No se han conseguido cultivar de manera aislada y los reactores que las contiene toman un color rojizo debido a la gran cantidad de citocromos que presentan para llevar a cabo su proceso oxidativo.
La oxidación anaerobia del amonio no se produce en la membrana citoplasmático sino que se lleva a cabo en la membrana del anammoxoma. El compuesto que actúa como intermediario entre los dos enzimas integrales de membrana que intervienen en el proceso metabólico es la hidracina que debido a su alta toxicidad es retenida en el anammoxoma para que no quede libre en el citoplasma.
NH 4 +^ + NO 2 - --------------> N 2 + 2H 2 O
Pueden ser consideradas a la vez como bacterias nitrificantes y desnitrificantes (respiran nitrito).
Microorganismos oxidantes de compuestos del azufre
Son mucho más diversas en cuanto a variedad de metabolismos que las nitrificantes. Utilizan una gran cantidad de sustratos oxidables (cualquier compuesto de S lo suficientemente reducido como para que la oxidación sea rentable) y el sulfato suele ser el residuo final. Sin embargo, no todas son capaces de oxidar su sustrato azufrado hasta sulfato (menor rendimiento energético).
En algunos casos, como en Thiomargarita namibiensis , se acumula S elemental en el interior (o exterior) de la célula en invaginaciones del espacio periplásmico, formando gránulos de S (bacterias que oxidan el H 2 S en S). Por ello, en ocasiones, se suelen adherir otras células a estos cúmulos para completar su oxidación. Esta misma bacteria también almacena el nitrato (NO3-) en vez de 02 que utiliza como aceptor electrónico en una gran vacuola central que ocupa gran parte de su citoplasma.
Otras como Thioploca forman filamentos compuestos por grandes células que forman masas visibles en la superficie de los sedimentos someros, presentan una respiración anaerobia y degradan residuos orgánicos.
Los residuos de sulfato elemental de estas dos bacterias sulfatoreductoras en disolución acuosa forman sulfhídrico que es usado por otras bacterias como sustrato, transformando el nitrato que usan como aceptor de electrones en amonio.
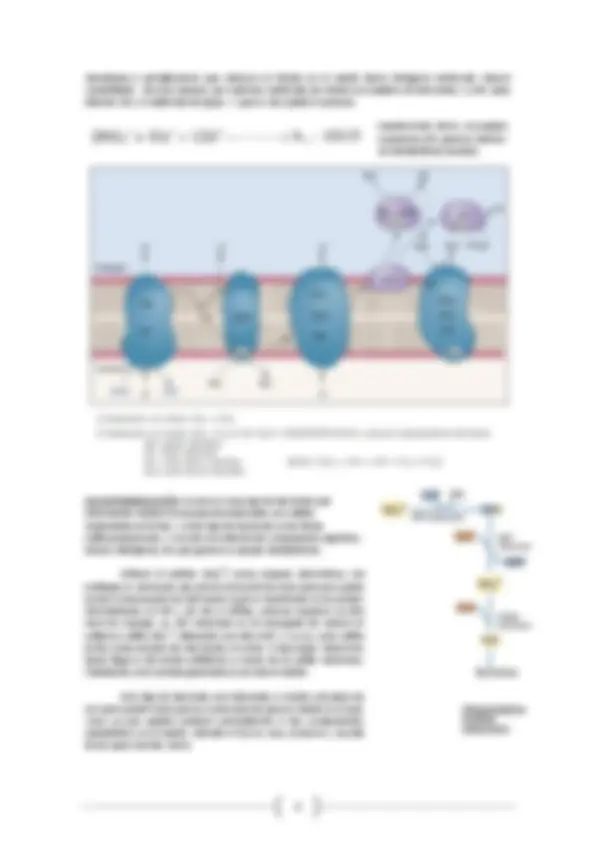
Dependiendo de cuál sea el sustrato oxidable que utilicen estos microorganismos los electrones ingresarán en la cadena de transporte electrónico por una vía diferente (VER FOTO), siendo el rendimiento energético mayor en aquellas bacterias que consuman un compuesto más reducido del azufre ya que al estar acoplado el bombeo de protones y a la formación de un gradiente a través de la membrana cuanto más largo sea el camino recorrido por los electrones en la cadena de transporte mayor translocación de H+ generarán. Además también será más corto el recorrido de electrones por la cadena de transporte inverso cuanto más alejado del citocromo C y del oxidador terminal se incorporen los electrones, teniendo que gastar menos gradiente de protones para obtener poder reductor. EJEMPLOS: si oxidan HS-, entra en una flavoproteína (FP) que se transforma en NADH o Q (más rentable energéticamente ya que requiere menor energía); si oxidan S 2 O 3 -^ o S (entran a la altura del cyt c por lo que requieren más energía para llevar a cabo el proceso y su rendimiento químico en menor).