¡Descarga Microbiología y más Apuntes en PDF de Biología solo en Docsity!
TEMA 10. LOS VIRUS
1.- Virología general 1.1.- Concepto de virus. Virus y viriones: tipos de viriones. Morfología y composición: ácidos nucleicos (tipo, estructura, número, polaridad) y capsómeros. Simetría de las nucleocápsidas. Enzimas. Viriones con envoltura. 1.2.- Crecimiento, detección y cuantificación de virus. Curvas de crecimiento: fase de eclipse, fase de estallido y rendimiento. 1.3.- Fases del ciclo general de replicación de un virus: entrada, decapsidación, expresión génica, replicación del genoma, ensamblaje y liberación. Modelos generales de replicación genómica en virus de DNA y RNA. Retrovirus. Clasificación de Baltimore. 2.- Bacteriofagos. 2.1.- Bacteriófagos: tipos. Fagos de arqueas. Características del ciclo lítico: adsorción específica y receptores. Regulación en fagos de RNA y DNA. Replicación. Ensamblaje y salida. 2.2.- Fagos atemperados y lisogenia: ciclos lisogénicos, regulación, inducción, inmunidad a la sobreinfección. Modelo en λ. Conversión lisogénica. Lisogenia en ambientes naturales. Transducción generalizada y especializada. 3.- Virus de eucariotas. 3.1.- Generalidades sobre virus animales: diferencias generales en su ciclo vital. Consecuencias de la infección vírica en células animales: infeciones latentes, persitentes y líticas. Transformación tumoral. 3.2.- Principales grupos de virus que afectan a humanos y otros vertebrados. Virus de invertebrados y virus de de microorganismos eucariotas. 3.3.- Virus vegetales: caracteres generales y principales grupos. 3.4.- Entidades subvíricas: Virus defectivos, viroides. Priones.
1. Concepto de virus. Virus y viriones.
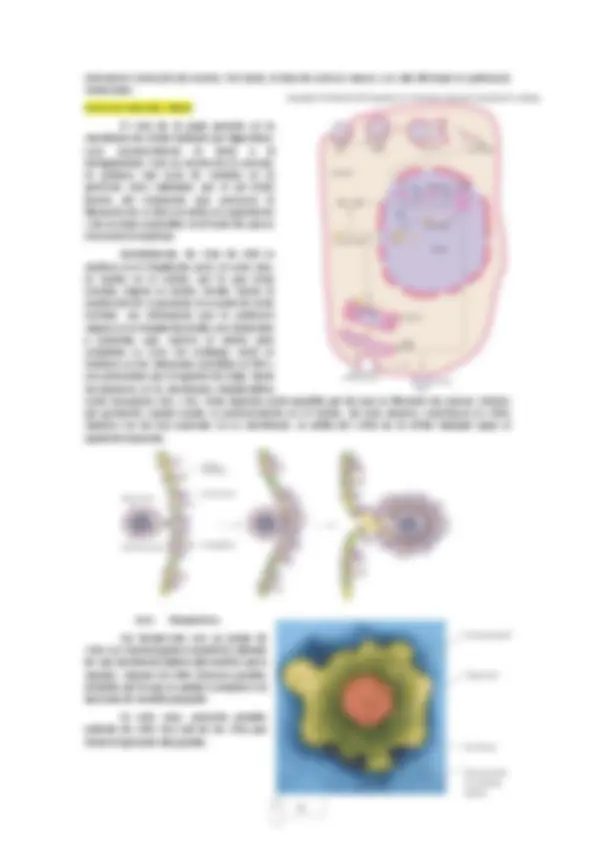
Los virus son entidades biológicas de naturaleza sencilla que no forman en ninguna fase de su desarrollo una célula viva, sino que son parásitos de otras células vivas de distintos tipos y que se comportan como parásitos intracelulares obligados de naturaleza no celular. De esta forma podemos afirmar que los virus no son células sino estructuras celulares. Por tanto, en su forma más sencilla, se tratan de partículas de pequeño tamaño constituidas por una envoltura proteica (cápsida) y una molécula de AN (DNA o RNA). Aunque no realiza las funciones de nutrición y relación, si son capaces de reproducirse. Su ciclo vital presenta dos fases:
Fase intracelular (activa): el virus se desensambla como partícula y solo encontramos sus componentes desasociados dentro de la célula. Fase extracelular (inerte): el virus es una partícula perfectamente definida (ácido nucleico envuelto como mínimo en una cubierta proteica). A esta partícula vírica que encontramos en la fase extracelular se le denomina virión. Un virión es la fase extracelular de un virus. El término virus incluye al virión y a la fase intracelular, pero el término virión solo es correcto para partículas víricas que viajan de una célula a otra (no incluye fase intracelular). Los virus envueltos presentan además una envoltura externa formada por fosfolípidos lo que puede modificar su morfología externa.
2. Tipos de viriones. Morfología y composición:
Los virus se clasifican en función de una serie de criterios:
Según el hospedador hablamos de
o Bacteriófagos (fagos) : virus que parasitan a procariotas (también afectan a arqueas). o Virus vegetales y virus animales : virus que parasitan a animales y plantas. o Micobirus : parasitan a hongos.
Además de estos tipos, existen virus que parasitan a algas y protozoos.
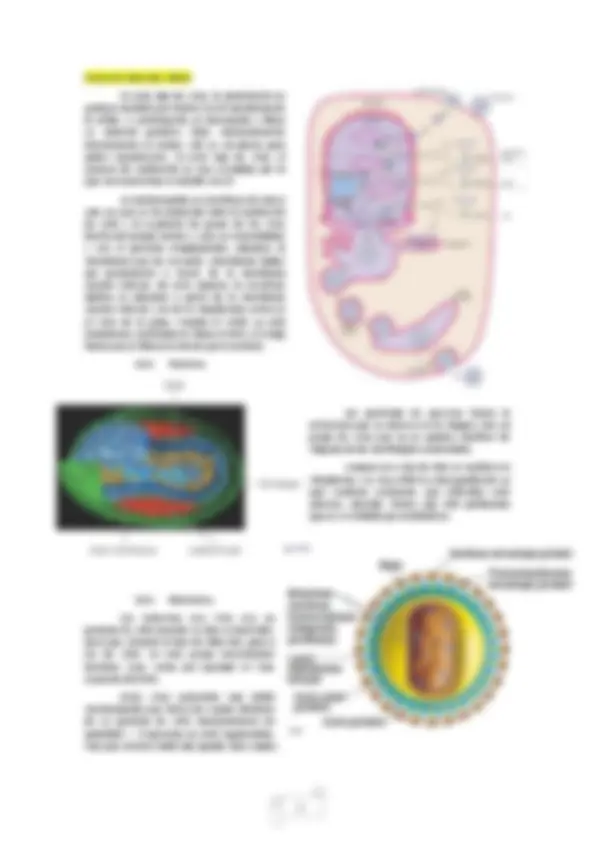
Características morfológicas de la partícula vírica (virión): Las cápsidas, en general, están constituidas por varias unidades proteícas denominadas capsómeros. De esta manera, existen virus que tienen todos los capsómeros iguales, pero hay otros con distintos tipos de capsómeros que se sitúan de forma diferente en esta envoltura. Además, al conjunto de cápsida y AN se le denomina nucleocápsida.
o Icosaédrica : es una de las dos morfologías básicas en viriones. Son virus casi esféricos rodeados por una cubierta de proteínas (cápsida) que contiene el ácido nucleico en su interior. Un ejemplo destacado es el virus de la gripe (c,h,i, j,k,l). o Filamentosa (helicoidal) : están formados por una molécula de ácido nucleico en forma de espiral a la que se arrollan los capsómeros, formando un filamento cuya longitud varía en función de la longitud del ácido nucleico. Un ejemplo conocido es el virus del mosaico del tabaco (TMV). Existen otros viriones que son rígidos y poco flexibles como el virus del évola, que es filamentoso y tiene un extremo enrollado como haciendo un nudo sobre sí mismo (a,b,d,e,m). o Bacteriofagos : son virus que combinan los dos tipos de estructuras básicos. Por un lado, la parte icosaédrica es la que constituye la cabeza donde se encuentra el AN y la parte helicoidal que constituye la cola del bacteriófago. Esta estructura es la que utiliza el virus para insertar el material génetico en el huespéd. A veces esta cola se acompaña de una placa basal y unas fibras que le sirven para reconocer y anclarse al huésped (f,g).
**DNA-RNA ss - ds lineal-circular segmentado o no
En esta imagen se representa la longitud del AN, el nº de pares de bases y el diámetro del virus de la gripe tipo A, el megavirus y, así, hasta llegar al pandoravirus que es el virus más grande que se ha encontrado. Este tipo de virus parasitan amebas y suelen confundirse con bacterias debido a su gran tamaño.
Sin embrago, hoy en día se siguen descubriendo virus de gran tamaño. Uno de estos, se ha descubierto al analizar muestras profundas de tundra en regiones que se están descongelando debido al calentamiento global.
Características de composición de los ANs que encierran esos viriones:
Los ANs pueden ser de ADN o ARN. No existen virus con ambos tipos dentro de la cápsida, pero algunos de ellos pueden alternan estas formas como material genético a lo largo de su ciclo vital.
El ácido nucleico puede ser monocatenario (ss) , una única cadena o bicatenario (ds) dos cadenas. Además, puede tener tanto forma tanto circular como lineal. Otro aspecto variable es que el genoma puede estar segmentado o no , es decir, que el virión tenga una o varias moléculas de AN. Por ejemplo, el virus de la gripe tiene el genoma segmentado y posee 8 moléculas independientes de ARN en las que cada una de ellas codifica para genes y proteínas diferentes, (no son copias idénticas). Además, los genomas segmentados son más proclives a combinar distintos fragmentos de virus diferentes por lo que aumenta su probabilidad de recombinación genética.
Si se trata de un genoma monocatenario, podemos clasificarlo según la secuencia de nucleótidos que contiene en:
o Polaridad + : el genoma tiene la misma secuencia de nucleótidos que tiene el mensajero del virus para codificar esa o esas proteínas (no es necesario leer su secuencia). o Polaridad - : el genoma tiene la secuencia complementaria a la que tiene el mensajero por lo que se requiere que sea transcrito por la hebra molde para promover su síntesis. El virus de la rabia tiene un genoma de ARN monocatenario -, por lo que el mensajero que se forma tendrán la secuencia complementaria a la del AN que tenemos en la cápsida. En cambio, aquellos virus que tengan polaridad +, pueden entrar en la célula y funcionar directamente como mensajeros. Combinando todos estos factores, podemos clasificar a los virus tal y como representa este tabla:
3. Enzimas: Dentro de los viriones, a parte del AN también podemos encontrar una gran variedad de enzimas necesarias en el momento de entrada o infección del huésped. La forma más sencilla de encontrar estas enzimas son empaquetadas con el genoma vírico. Un ejemplo de estas enzimas son la replicasa o la retrotranscriptasa. También existen virus con metilasas o los fagos también pueden tener enzimas con actividad icolisocimas que al reconocer el virus a la bacteria hospedadora, forma un agujero para inyectar el genoma del virus a través de las colas de los fagos binarios por lo que es necesario romper la pared celular para producir la penetración de su material genético. 4. Fases del ciclo vital de virus: Penetración. Esta fase engloba dos procesos: adsorción específica y pérdida de cubierta. Por un lado, en la mayoría de los virus se produce la adsorción o reconocimiento específico a través de la interacción con unos receptores que se sitúan en la superficie de la célula hospedadora. En muchos casos, se trata de un paso imprescindible como en los fagos y virus de animales donde se produce la interacción de un componente de la cápsula o de la membrana del virus con un componente de la superficie de la célula huésped. Es un reconocimiento de la misma especificidad de la interacción antígeno-anticuerpo.
Si no se produce este reconocimiento, no se produce la infección por más que el virus pueda reproducirse pero es capaz de penetrar en ella. Sin embargo, si desaparece el receptor, no puede penetrar y la célula se vuelve inmune a él.
Una vez que el virus ha sido reconocido, debe penetrar en el interior de la célula lo que implica la pérdida de la cubierta como paso previo a la entrada. En los fagos, el virus no entra totalmente a la célula sino que únicamente penetra el ácido nucleico y algunas enzimas pero la cápsida queda fuera. Sin embargo, en otros virus, penetra todo o casi todo el virión y se produce la perdida de la cubierta a través de la apertura y desintegración de los capsómeros. A continuación, los viriones se desensamblan y liberan sus componentes llevando a cabo una serie de reacciones (paso característica que los diferencia de los parásitos).
Procesos víricos intracelulares : dentro de la célula se producen unos procesos que consisten en 3 fases: o Replicación de genoma viral (veremos a continuación). o Expresión de genes víricos y regulación : simultáneamente se produce una expresión y regulación de los genes virales. En algunos casos puede ocurrir antes o después de la replicación del genoma. o Ensamblaje de los componentes que formarán las partículas víricas (Cápsida, envoltura y AN).
Salida/ liberación de nuevos viriones : se puede producir por destrucción de la célula en la que se encuentran los virus (lisis) o pueden salir de la célula sin romperla a través de distintos mecanismos.
4.1. Modelos generales de replicación genómica en virus de ADN y ARN. Retrovirus:
Hay mucha diversidad de comportamientos en la replicación del genoma, en función de que tipo se trate:
-Virus de DNA:
Si son lineales: se produce la circularización , que puede tener lugar por la existencia de extremos cohesivos que se asocian entre sí porque son complementarios y se cierran mediante una ligasa o bien porque el genoma lineal tenga redundancia terminal, de forma que se recombinan los dos extremos de la cadena por lo que el genoma se vuelve circular.
Si son monocatenarios: se convierte en bicatenarios utilizando habitualmente la actividad enzimática de la célula hospedadora.
En la clase IV hay virus con genoma de ARN de cadena simple y polaridad +.
En la clase V encontramos virus de ARN monocatenario de cadena -, tenemos el ejemplo del virus de la gripe o de la rabia.
En la clase VI, encontramos virus de ADN de cadena simple que se replican con intermediarios de cadena de ADN, retrovirus.
En la clase VII, son virus que contienen ADN bicatenario pero se replican usando un ARN intermediario monocatenario. Los grupos 6 y 7 hacen lo mismo, el ciclo es similar, pero unos encapsidan ADN bicatenario y otro ARN. Además, ambos usan la retrotranscriptasa para mediar en el ciclo de replicación.
Como vemos, según esta clasificación en función del tipo de genoma que encapsidan, tienen distintas formas de formar el mensajero. La estrategia para la producción de mensajeros es similar en virus de clase I, II y VII. Los de clase I y VII son virus que encapsidan moléculas de ADN doble mientras que los virus de clase II encapsidan ADN monocatenario + por lo que necesitan la síntesis de la cadena complementaria, actividad producida por las enzimas de la célula hospedadora y, cuando tenemos en forma bicatenario, la transcripción de la hebra - da igual al ARNm.
Por otro lado, los virus de la clase 3, los que tienen genoma de ARN bicatenario en el que la hebra positiva (tono más oscuro) es usado para copiar el complementario mientras que la hebra la hebra – se transcribe para producir así el ARNm. Los virus de la clase IV usan su genoma directamente como mensajero. Los virus de la clase V tienen que replicarse con ayuda de la replicasa y la copia de la hebra - produce el mensajero.
Por último, los virus de la clase VI tienen que llevar a cabo la transcripción reversa como paso previo para producir los genes virales y que se expresan. En este proceso transforma el RNA en ADN bicatenario. Una vez así, sigue el mismo camino que el de las clases I, II y IV en el que se transcribe la hebras - por la ARN polimerasa y produce una hebra + del RNA que actúa como mensajero.
5. Crecimiento, detección y cuantificación de virus. Hoy en día, se conocen métodos para analizar la cinética de multiplicación de los virus, es decir, como se incrementan las poblaciones virales durante la infección de la célula hospedadora. Por ello, para detectar las partículas víricas se utiliza el microscopio electrónico. Además, se requiere un hospedador adecuado para que se produzca la infección. Por lo tanto tenemos que ofrecer al microorganismos unas condiciones óptimas de nutrientes para que pueda desarrollarse y conseguir así, más adelante, que lo viriones puedan desarrollarse. Por todo esto, es necesario cultivar a las células hospedadoras antes de introducir los virus.
De esta manera, conseguiremos una capa continua de células del hospedador (césped bacteriano) y, una vez que se ha producido el crecimiento de esta capa, los viriones son introducidos. Estos viriones provocarán la lisis de un cierto número de células y, en las siguientes generaciones de virus, estos entraran en las células vecinas y seguirán lisándolas hasta que, finalmente, se forma una zona donde todas las células han sido lisadas y, por tanto, no observamos la opacidad típica del césped del huésped. Estas zonas se conocen como calvas (“plaques” en inglés).
En la foto de la derecha podemos ver el efecto que tienen sobre el césped distintas concentraciones de viriones. De esta manera, podemos observar que los que tienen tasas de dilución altas (10-5) apenas hay césped (muchas calvas y las células vecinas pueden crecer si son fagos atemperados) mientras que en las más diluidas (10-8) el efecto es menos acusado (menos calvas, más césped). De esta manera, podemos enumerar las calvas existentes en una placa para poder determinar su crecimiento y el número de partículas infectivas que teníamos.
Las calvas no pueden regenerarse ya que al intentar volver a crecer en este lugar las células hospedadoras se vuelven a lisar por los viriones. Por lo tanto, en estas zonas no es posible el recrecimiento de la calva porque está repleta de viriones infectivos, a no ser que sean mutantes no reconocidas por los fagos.
Una condición que debe de cumplir el césped es que se encuentre en crecimiento exponencial ya que los virus son parásitos que carecen de la capacidad para generar energía o realizar su biosíntesis y, por tanto, es necesario que lo obtengan a partir del huésped, excepto su información genética y lo que esto codifica. Por lo tanto, las células que forman el césped tienen que ser células que crecen exponencialmente ya que cuando las células del hospedador entran en la fase estacionaria, no son capaces de proporcionarles lo necesario para crecer.
Los virus no siempre producen la lisis de las células que los hospedan. En algunos casos, cuando se liberan, no la destruyan y de esta forma no se podrían detectar los virus contando las clavas formadas.
5.1. Crecimiento de virus animales:
Para ver el crecimiento de virus de animales necesitamos una célula animal y no una bacteria (más común) y que el cultivo presente una línea celular del hospedador adecuada para ser empleada como césped. Uno de los problemas de estos virus, es que se desconoce las líneas celulares de algunos viriones. Por ello, en muchos casos, se suele trabajar con organismos enteros (huevos embrionarios).
donde se produce la infección, no podemos detectar ninguna partícula vírica infectiva ya que, en esta fase, el virus no está como partícula vírica infectiva. En esta fase es donde se produce la replicación y la expresión del genoma vírico. o Maduración : fase en la que se empieza a ensamblar los componentes de viriones y donde se sintetizan las proteínas que formarán parte de la cápsida.
2º etapa: en este período, los virus crecen de forma brusca alcanzando inmediatamente el número máxima de viriones, valor a partir del cual se estabiliza. Este tipo de crecimiento, llamado incremento o crecimiento en escalón, nos da lo que se llama el tamaño de la explosión , número medio de viriones que se produce por cada célula infectada con un solo virión. En muchos casos este valor ronda los 100 (libera 100 viriones por célula infectada por un solo virión).
7. Bacteriófagos: Los bacteriófagos, como ya hemos visto, se pueden clasificar en función de su morfología o del ácido nucleico que tenga. De esta manera, pueden ser de ARN o ADN, de cadena simple o doble. Además, podemos clasificar a los fagos en dos tipos básicos en función de su comportamiento cuando infectan una célula:
Fagos virulentos : desencadenan siempre el ciclo lítico, es decir, siempre que entran en célula hospedadora se reproducen rápidamente produciendo una nueva generación de viriones que se liberan al exterior provocando o no la lisis de la célula. Un ejemplo es el fago T4.
Fagos atemperados : estos fagos penetran en célula hospedadora y se establece como unidad genética autoreplicativa pudiendo formar parte del cromosoma de la bacteria o no (plásmido). Además de poder realizar esto, también pueden producir un ciclo lítico, al igual que los fagos virulentos, cuando reciben un estímulo. Sin embargo, se diferencia ya que este proceso no ocurre de manera inmediata y no tiene por qué llevarse a cabo en todas las células. Un ejemplo es el fago lambda. Los fagos más conocidos son aquellos que tienen estructura binaria, aunque también hay icosaédricos o filamentosos. Dentro de fagos con estructura binaria, se distinguen en dos tipos:
Fagos con cola muy corta (T3,T7) Fagos con cola larga que puede ser contráctil como los fagos de la serie tepar (tiene dos cilindros, el exterior es contráctil y se contrae al introducir el material).
CICLO LÍTICO. 1. Penetración 2. Síntesis del genoma y de las proteínas víricas 3. Maduración y ensamblaje 4. Liberación fagos virulentos y temperados
CICLO LISOGÉNICO fagos atemperados únicamente.
7.1. Fago T4: Como ejemplo de fagos virulentos, estudiaremos el fago T4. Para que se produzca la infección es necesario que una parte del fago sea reconocida por una molécula de superficie de célula hospedadora. Esta unión se trata de una unión específica. En el caso del fago T4, el reconocimiento se produce entre la placa basal de la cola del fago y las fibras del lipopolisacárido de E. coli (célula huésped).
Una vez que se ha producido el reconocimiento, la vaina externa de la cola se contrae y el cilindro interno penetra a través de las envolturas mediante una actividad de tipo lisozima formando un pequeño agujero en la parte del peptidoglucano de la pared. De esta manera, se inyecta a presión el ADN de doble cadena que se contiene en el fago. Así, el ADN entra totalmente desnudo y el resto de componentes del virión quedan fuera.
En este tipo de infecciones solo penetra el ADN y, a lo sumo, alguna enzima, pero nunca la cola ni la cápsida ni el resto de componentes que se quedan totalmente fuera.
7.2. Receptores de fagos:
Además de los lipopolisacáridos los fagos pueden presentar otros receptores. Cualquier molécula superficial lo suficientemente grande puede servir como receptor de fagos. Además, existen bastantes fagos que puede usar como receptores el flagelo, el pili sexual u otra estructura parecida.
Como vemos en la tabla, en E.coli podemos encontrar varios tipos de fagos tanto filamentosos como icosaédricos que reconocen y utilizan como receptores proteínas o glicoproteínas que constituyen el pili sexual. Otros emplean proteínas que forman parte de la membrana externa de las Gram -, ácidos teitocicos en Gram +, pero, sin duda, la mayoría de ellos son reconocidos por lipopolisacáridos. VER TABLA. En los virus envueltos, se produce fusión de membranas entre la envuelta del virus y de la célula o puede incorporarse por endocitosis.
Esto nos permite observar la gran especificidad que existe en el reconocimiento de fagos con los receptores de las células hospedadoras (similar al que existe entre antígeno-anticuerpo).
En una especie como la Salmonella enterica tiene varios miles de serotipos caracterizados cada uno en función de sus antígenos. Serotipado (el receptor es de una bacteria) y fagotipatos (el receptor es de un fago).
En la imagen se nos muestra varios viriones adosados a la superficie de la célula hospedadora y se puede apreciar que algunos tienen la cabeza opaca (no han vaciado su contenido) y otros transparentes (ya han liberado su material genético en el citoplasma del huésped). En ella, también podemos la cola contraída de 3 viriones, el cilindro externo rígido.
El ADN dentro de estos fagos está a presiones muy elevadas de tal forma que, en el momento que se abre, el ADN sale con gran presión en el citoplasma de la célula hospedadora (espacio viscoso). Al salir la cabeza queda transparente.
capaces de seleccionar los fragmentos de ADN que penetran en la célula que son reconocidos por los ARNs sin necesidad de etiquetado produciendo la transcripción y degradándolo sin necesidad de que haya este proceso de enzimas de restricción.
Esta es una forma de memoria de los ADN con los que han tenido contacto la célula. Este sistema combina la acción de las enzimas CAS y los ARN producidos por la traducción de CRISPR para evitar la infección y degradar los ANs de los viriones.
9. Expresión del genoma vírico: Si el DNA no es degradado rápidamente, comienza la replicación y la síntesis de nuevos fagos. Poco después de la penetración, empiezan a expresarse los genes tempranos. Entre estos genes encontramos los que codifican los genes de ARN temprano, que contienen nucleasas que degrada el ADN bacteriano, impidiendo que pueda reproducirse y además se generan precursores para la síntesis del fago como nucleótidos para poder sintetizar la nueva cadena. En el caso del fago T4 solo usan 3 nucleótidos, la citosina debe de sufrir una transformación, por lo que las enzimas necesarias para esto, se encuentran también en este ARN temprano. También, se produce otras enzimas que formarán parte del repliosoma y otras que modifican la especificidad de ARN polimerasa haciendo que se una únicamente a elementos víricos.
En la etapa media, se sintetizan otras proteínas y enzimas que participan en diversas actividades propias del virus pero algunos de los productos de estos genes son nuevas subunidades sigma codificadas en el genoma del virus que permitirán la expresión de los genes tardíos que son los últimos en sintetizarse y fabricar mensajeros y proteínas. Estos genes tardíos solo se expresarán cuando se hayan formado estas subunidades que modifican la especificidad de la RNA polimerasa. Por tanto, como vemos existe una cierta temporalidad en la expresión génica de virus durante la infección.
En la última parte, el genoma del virus ya se ha copiado, por lo que los genes tardíos sintetizan todos los componentes necesarios para formar los viriones. Se trata de un gran grupo de proteínas (son
- muy variable y diferente. Una vez se ha producido el ensamblaje de la partícula vírica, se expresan los genes que le ayudan a ser expulsado que comienzan a acumular lisozima para favorecer su lisis y poder liberar los fagos al exterior.
10. Ensamblaje y salida de los viriones:
El ensamblaje tiene 3 líneas de montaje. Por un lado, se inserta la cabeza en forma de procabeza vacía, luego se empaqueta con ayuda de un motor que mete el materia genético a presión llenando las cabezas y sellándolas y, por último, se produce su ensamblaje (se ensambla la placa basal, la cola y se incorporan ambas cosas. Lo último en unirse son las fibras de la cola a la placa basal). En este proceso, también intervienen una gran cantidad de proteínas y enzimas específicas.
Mientras se está produciendo este ensamblaje, se va produciendo y acumulando lisozima para que se pueda producir la lisis de la célula y se liberen los viriones.
Hay otras fagos, sobre todo los filamentosos, que una vez completado su ciclo de expresión y replicación del genoma se ensamblan formando filamentos al tiempo que salen de la célula sin lisarla. Estas células crecen más lentamente pero sintetizan todo lo necesario para abastecer al fago de sus necesidades. Una vez que el fago presenta todos los componentes necesarios para su desarrollo, sale al exterior.
El fago tiene ADN de cadena simple circular cerrado y su filamento es asimétrico ya que los capsómeros se disponen de forma asimétrica (diferentes tipos de capsómeros). En un extremo tiene los capsómeros tipo A y en el otro, los capsómeros tipo C. Esto nos permite distinguir entre la cabeza y la cola y los capsómeros se incorporan en la membrana citoplásmica de la bacteria a la vez que sale de la bacteria.
Sale encabezada por el sitio donde tiene el capsómero tipo A y conforme sale se le van añadiendo el tercer tipo de capsómeros hasta que al final del proceso sale el extremo C. Sin embargo, cuando parasita, la proteína C es la que marca la entrada (al revés).
Como sabemos, en estos casos, la salida del virión no lisa ni destruye la célula por lo que esta sigue siendo activa y puede estar emitiendo estos filamentos (partículas viricas) por lo que no produce calvas en el césped donde estén los virones ya que no las lisan, pero si se ve que son zonas donde hay menos densidad de cultivo (el cultivo es más tenue y el crecimiento más lento da lugar a una menor opacidad en el césped).
11. Fagos atemperados y lisogenia: Los fagos atemperados tienen la capacidad de establecerse de una forma reversible como parte del genoma de la célula a la que han infectado y pueden realizar tanto el ciclo lítico como el lisogénico. Estos fagos solamente pueden ser fagos de ADN. Este tipo de ciclos, en los que el genoma del virus se integra en el genoma del hospedador, se llama lisogenia. El termino lisogénico se reserva para cepas bacterianas que son portadora del profago (fago es atemperado).
El estado en el que el material genético está establecido de manera reversible al genoma del hospedador se llama profago o pro-virus. Este profago se replica cada vez que la célula bacteriana lo hace. Las modalidades en la que podemos incorporar y estabiliar el profago en el cromosoma del huésped son dos:
Por inserción en el cromosoma de la bacteria. Este proceso lo pueden hacer de dos maneras: incorporando su material genético en lugares específicos que son reconocidos por los fagos. Esta forma es la más común y se en fagos como el fago lambda de E.coli que se ensambla entre operones gal y bio de E.coli. Por otro lado, también encontramos fagos que incoporan su AN en una zona al azar. Un ejemplo de este tipo de fagos es el fago Mu (de mutagénico), que se inserta en el genoma del huésped al azar Por mantenimiento a modo de plásmido. En este caso, los fagos estabilizan su material genético a través de un plásmido autoreplicativo que se hereda como otro plásmido de la bacteria a sus descendientes. Ambos tipos de fagos, los que se insertan al azar y los que se quedan como plásmidos, no pueden dar lugar a procesos de transducción especializada.
Los profagos producen de manera continuada represores que bloquean todos los promotores de los genes implicados en el desarrollo de un ciclo lítico y en la producción de ciertos viriones por lo que se impide la inducción al ciclo lítico ya que se impide la expresión de los genes implicados en este proceso.
En algunos casos, la cepa portadora de un profago no se ve afectada ni tiene ninguna propiedad extraña excepto la inmunidad a la sobreinfección. Sin embargo, en otros casos, la cepa es capaz de desarrollar capacidades que no tienen las bacterias no infectadas. En estos casos, el fago dota a la bacteria de unas características especiales que modifican el comportamiento del fago ( conversión lisogénica o conversión fágica ). Este proceso consiste en la adquisición de una propiedad que esta codificada por el genoma del fago pero expresada por parte de la bacteria que es portadora del profago.
Uno de los ejemplos más característicos y destacados consiste en la adquisición de la bacteria para producir toxinas, proceso importante en el desarrollo de una enfermedad, la toxina diftérica solamente la produce la cepa de Colina bacterium que son portadoras de un fago atemperado llamado fago beta. En muchos casos, la bacteria que lleva el profago es mucho más virulenta que el profago aislado, (leer) la toxina no está en genoma de la bacteria, solamente la posee aquellas que tienen un fago beta integrado. Esta toxina es muy potente y es capaz de paralizar la síntesis proteica de toda una célula por una modificación en los factores de elongación de células eucariotas y arqueas. Esta proteína no le hace falta al fago, pero la proteína que produce hace que la infección sea más grave y virulenta, pero al profago no le soporta ningún beneficio.
La toxina botulínica esta codificada por una bacteria esporulada anaerobia estricta que son toxinas más potentes del mundo bacteriano, también algunos de los tipos, al menos 2 o 3 están codificadas en genoma de profagos La toxina del cólera también esta codificada en el genoma de un fago filamentoso y se establece en ellas cepas del vibrio cólera que son patógenas y producen el cólera (leer).
A veces la presencia del fago dota a la bacteria de un arma adicional para defenderse de otros fagos diferentes a él mismo, dota de un sistema de detección y modificación que detecta genes externos y los destruye (otras características distintas a las toxinas).
12. Virus de eucariotas: Los virus que parasitan células eucariotas parasitan células que se encuentran muy compartimentadas y cuya expresión génica también puede estar muy compartimentarizada. En estos casos, los virus de ADN es normal que se replican y expresen en el núcleo mientras que los que tienen un genoma de ARN se replican y se expresan en el citoplasma (hay excepciones).
Centrándonos en los virus animales, los virus se clasifican en virus de DNA o RNA y con o sin envolturas. Los más comunes son aquellos que presentan envolturas lipídicas que provienen del huésped pero cuyas proteínas son sintetizadas por el propio virus.
En cuanto al primer paso en el proceso de infección, en los virus animales es necesario el reconocimiento específico del receptor. Si la célula o tejido no tiene esos receptores específicos, la célula no será infectada por el virus animal. De esta manera se establece los tejidos que va a sufrir infección y cual no. En el caso de los poliovirus solamente encuentra receptores específicos en las células de la mucosa nasofaríngea, en el intestino, y en las células del asta interior de la médula espinal. Sin embargo hay otros como el virus que usa el sarampión que usa un receptor que está en todos los tejidos de organismos, esto lo determina la existencia de glicoproteínas que usan el virión como receptor específico.
12.1. Diferencias generales en su ciclo vital.
En virus animales hay 4 modalidades de infección:
Modalidad equivalente al ciclo lítico : multiplicación rápida del virus en la que, una vez que se ha reproducido, provoca la muerte y la lisis de la célula, liberando los viriones formados. Este tipo se conoce como infección aguda, lítica o citocida. La muerte de la célula puede deberse a:
o Inhibición de los procesos biosintéticos de la célula animal. El virus inhibe la síntesis del ARN o del ADN y aprovecha los recursos de la célula para completar su desarrollo. o Formación de cuerpos de inclusión que son agregados de gran tamaño formados por fragmentos de partículas víricas, ribosomas, fragmentos de cromatina,.. Además, pueden formar lisosomas y hacerlos más permeables acumulando sustancias tóxicas del citoplasma que pueden destruir a la célula. La célula puede morir también porque, como parte del proceso de infección, se introducen proteínas víricas en la membrana de la célula que son reconocidas como extrañas por el propio sistema inmunitario y son destruidas. Infección latente : el virus penetra en la célula, libera su ADN al descapsidarse y no se produce ninguna alteración ya que el virus se encuentra en forma inactiva ya que su material genético no se inserta en el genoma de la célula. Lo que sí que ocurre es que esa célula es portadora del virus inactivo y pueden permanecer años con este virus inactivo pero pueden producirse fenómenos que permitan la activación de éstos y pasar así a desarrollar un ciclo como el descrito anteriormente. Un ejemplo destacado es el herpesvirus. Infección crónica : la célula infectada produce de forma más o menos lenta la liberación progresiva de virones, pero sin lisar la célula. En muchas ocasiones, este tipo de infección se ha asimilado al ciclo lisogénico que experimentan los fagos de bacterias. Transformación tumoral o malignación de células infectadas por virus(transformación en célula maligna) : en algunos casos, estas infecciones provocan que las células se vuelvan tumorales ya que pierde los controles de su propio crecimiento bien porque el virus introduce un oncogen o porque el virus afecta el sistema de regulación del oncogen de la célula y esta empieza a expresarse transformando la célula en tumoral (división rápida e incontrolada).
**13. Principales grupos de virus que afectan a humanos y otros vertebrados:
- Virus de invertebrados y virus de microorganismos eucariotas:**
Virus importantes para la salud humana (Tabla-ResumenIMPORTANTE conocer sobre todo ejemplos y enfermedades que producen). 1. Picornavirus. Son los virus más pequeños.
14.1. Rhadovirus
Fotografías con microscopia electrónica donde vemos su forma característica (virus helicoidal). Dentro de la cápsula tiene una molécula de RNA monocatenaria – por lo que requiere distintas enzimas para completar su ciclo vital como la replicasa.
14.2. Virus de la gripe El virus de la gripe consta de 8 fragmentos de ARN monocatenario – en los que, cada uno de ellos, codifica para 1 o 2 proteínas (en total codifica 10 proteínas). El núcleo se encuentra rodeado por una envoltura proteica (cápsida o matriz forma icosaédrica) que, a su vez, está rodeada por una membrana lipídica que derivan de la membrana citoplásmica de la célula parasitada pero que contiene dos espículas diferentes, la neuroaminidasa (NA) y la hemaglutina (HA) que son las estructuras encargadas de mediar el reconocimiento de la célula que va a ser infectada (determinan sus propiedades antigénicas). Los números de la gripe hacen referencia al tipo de espículas que tienen los virus en su superficie y nos permite determinar el grado de virulencia de cada virus (gripe N1H1). Los ortomixovirus no solo tienen humanos como hospedadores, sino que también pueden actuar sobre animales como aves, cerdos,...
Los virus con DNA son menos reactivos (menor capacidad para mutar) por lo que presentan menos capacidad para matar. Además, presentan la DNA polimerasa que tiene actividad exonucleasa que participa en la corrección de errores. Por otro lado, los virus de RNA mutan más rápidamente (son más reactivos) y esto se debe a que la RNA polimerasa no tiene esa actividad por lo que no presentan un
sistema de corrección de errores. Por tanto, la tasa de error es mayor y es más fácil que se produzcan mutaciones.
CICLO DE VIDA DEL VIRUS :
El virus de la gripe penetra en la membrana de célula huésped por fagocitosis, cuyo reconocimiento se debe a la hemaglutinina. Una vez dentro de la vesícula, se produce una serie de cambios en la partícula vírica mediados por el pH ácido dentro del endosoma que provocan la liberación de su RNA envuelto en capsómeros y las enzimas contenidas en él entre las que se encuentra la replicasa.
Normalmente, los virus de ARN se replican en el citoplasma, pero, en este caso, se replica en el núcleo, por lo que estas enzimas migran al núcleo, donde inician la replicación de su genoma con ayuda de estas enzimas. Los mensajeros que se producen migran en el citoplasma donde son traducidos a proteínas que vuelven al núcleo para completar su ciclo. Sin embargo, otros se traducen en los ribosomas asociadas al RER y son procesados por el aparato de Golgi hasta incorporarse en la membrana citoplasmática como receptores (NA y HA). Estas regiones serán aquellas por las que se liberarán los nuevos viriones por gemación cuando acabe su procesamiento en el núcleo. De esta manera, constituyen el virión maduro con las dos espículas en su membrana. La salida del virión de la célula huésped sigue el siguiente esquema:
14.3. Herpesvirus. Los herpesvirus son un grupo de virus con nucleocapsida icosaédrica rodeada de una membrana lipídica pleomorfica (poco regular). Algunos de ellos alcanzan grandes tamaños por lo que se pueden comparar con bacterias de tamaño pequeño.
En este caso, presenta grandes cadenas de ADN. Son uno de los virus que tienen el genoma más grande.