



Retículo
endoplasmático













Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: BCHA/HA, Profesor: Laura Tussell, Carrera: Microbiologia, Universidad: UAB
Tipo: Apuntes
1 / 22

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
El retículo endoplasmático (RE):
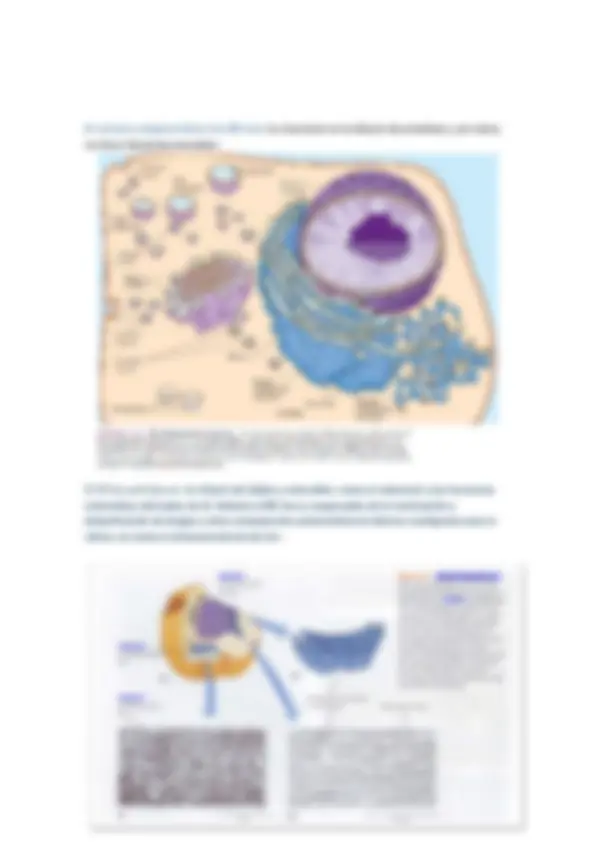
Es una red continua de sáculos aplanados, túbulos y vesículas, que se distribuye por todo el citoplasma de las células eucariotas. (Endoplasmático significa “dentro del plasma” y retículo, significa red pequeña)
Los sacos denominados cisternas del RE y el espacio es conocido como luz o lumen del RE. En una célula típica de mamífero del 50 al 90% del componente membranoso está representado por el RE.
Los dos tipos básicos de retículo endoplasmático difieren en su estructura y función.
Se distinguen por a presencia o ausencia de ribosomas enanclados a la cara externa de su membrana.
El retículo endoplasmático rugoso (RE rugoso) tiene ribosomas unidos en la cara citosólica (externa) de su membrana.
El retículo endoplasmático liso (RE liso) tiene tal apariencia por la ausencia de ribosomas en su membrana.
Ambos retículos, pueden distinguirse además por su morfología, las membranas del RE rugoso, forman en general, sacos aplanados, mientras que las del RE liso tienden a formar túbulos.
La presencia o ausencia de ribosomas, refleja la diferencia funcional entre los dos tipos de retículos de las células eucariotas. Las células que llevan a cabo una importante actividad de síntesis de proteínas, suelen tener una red muy desarrollada de retículo rugoso. Por otra parte, en las células que producen hormonas esteroícas, hay un claro predominio del RE liso.
En el REL se lleva a cabo la hidroxilación de diversas sustancias, como fármacos, drogas… La hidroxilación aumenta la solubilidad en agua de la mayoría de las drogas hidrofóbas. Esta modificación es critica pues la mayoría de los compuestos hidrófobos son solubles en los lípidos de las membranas y son, por tanto retenidos por el organismo, mientras que los compuestos hidrosolubles se eliminan fácilmente, primero hacia la sangre, y luego hacia el exterior (excreción).
En las membranas del REL se sintetizan todos los lípidos con dos excepciones:
Ácidos grasos (citosol) 2 fosfolípidos mitocondriales ( cardiolipina y fosfatidiglicerol ) en la mitocondria
También ocurre la biosíntesis de fosfolípidos, como la fosfatidilcolina, que tiene lugar en la monocapa citosólica del REL, ya que las 3 enzimas que intervienen tienen un centro activo en este lado de la membrana. Estas tres enzimas son:
Los lípidos intermediarios se pueden difundir de una monocapa a otra.
Los fosfolípidos son añadidos en la capa C de la membrana del REL, pero nunca en la capa E. Las flipasas específicas trasladan determinados fosfolípidos de la capa C a la capa E de la membrana para conservar la distribución inicial. Aquí se forma la asimetría de la membrana.
Adquisición de la topología (asimetría) de lípidos de membrana
Gracias a las lipasas específicas. Todos los lípidos son creados en la capa C, las flipasas específicas solo trasladan determinados fosfolípidos a la capa E.
Cuando las vesículas formadas en el RE se fusionen con otros orgánulos del sistema endomembranas, esta distribución diferencial establecida en el RE liso, se transferirá a las otras membranas celulares.
El movimiento de los fosfolípidos desde el RE a las mitocondrias y los cloroplastos entraña una dificultad añadida. A diferencia de los orgánulos del sistema de endomembranas, las mitocondrias y los cloroplastos no crecen por fusión con vesículas derivadas del RE. En lugar de eso, en el citoplasma existen unas proteínas de intercambio de fosfolípidos (o proteínas de transferencia de fosfolípidos), que llevan a los fosfolípidos desde la membrana del RE, a las membranas externas de las mitocondrias y los cloroplastos.
Cada tipo de proteína de intercambio reconoce un tipo específico de fosfolípido, que extrae de una membrana y lo lleva a través del citosol, a la otra membrana. Esta transferencia contribuye también al movimiento de fosfolípidos desde el RE a otras membranas de la célula, incluyendo la membrana plasmática.
Los ribosomas unidos a la cara citosólica de la membrana del RE rugoso, son los responsables de la síntesis de proteínas, tanto solubles, como de membrana. Estas proteínas suelen incorporarse en los sistemas de endomembranas y en la membrana plasmática, o bien son exportados como productos de secreción.
¿Cómo entran las proteínas en el sistema de endomembranas? Muchas de ellas son descargadas hacia la luz del RE rugoso durante su propia síntesis. Después de la biosíntesis las proteínas de membrana quedan ancladas a la membrana del retículo por sus regiones hidrófobas o por uniones covalentes de lípidos de membrana. Las proteínas solubles y la mayoría de secreción se vierten a la luz del RE.
Además de la síntesis de polipéptidos , el RE es el lugar donde se producen las primeras etapas de adición y procesamiento de los grupos hidrocarbonados de las glicoproteínas, el plegamiento de proteínas, el reconocimiento y eliminación de polipéptidos mal plegados y el ensamblaje de proteínas multiméricas.
Además de facilitar el plegamiento y ensamblaje de las proteínas, el RE es el lugar de control de calidad.
Síntesis de proteínas en el RER (Seminario)
Resumen de requerimientos para que una proteína continúe síntesis en RER:
Todas las proteínas empiezan su síntesis en el citosol. Las proteínas que tengan una secuencia señal de importe al RE se dirigirán a las membranas del RE rugoso para continuar su síntesis. Estas señales suelen estar situadas en el extremo N-terminal de la proteína (inicio). Estas señales son muy heterogéneas (contienen entre 15-30 aminoácidos; 4-13 hidrofóbico).
La secuencia señal de importa al RE es reconocida cuando emerge del ribosoma por la partícula SRP (o partícula de reconocimiento de la secuencia señal). Para que la proteína sea reconocida tiene que tener unos 70 aminoácidos (30 aa en el interior del ribosoma y de 15- aa de la secuencia señal de importe). La partícula SRP dirige a la proteína (y ribosomas unidos a ella) a un receptor específico situado en la membrana del RER ( receptor de SRP ). El SRP tiene dos dominios: uno de unión a la secuencia señal y ribosoma y el otro de unión al receptor de SRP. Tiene una forma similar a un ARNt y está compuesto por 1 molécula de ARN ( nucleótidos) y 6 unidades proteicas.
En el citosol, la unión del SRP al péptido señal y ribosoma causa una pausa momentánea en la traducción. En la membrana del RER, habrá un receptor del ribosoma, un receptor del SRP y un complejo de translocación (proteínas Sec61). El SRP se une a su receptor. El SRP se liberará de su receptor cuando la proteína quede unida al complejo de translocación (Sec61), donde se continúa la síntesis de la proteína. La unión y liberación del receptor del ribosoma sigue los mismos pasos del receptor del SRP.
Modificaciones químicas de proteínas del RER
-Inicio de la N-glucosilación de proteínas
-Rotura proteolítica específica
Modificaciones químicas de proteínas del RER
-Inicio de la N-glucosilación de proteínas (importante)
Plegamiento de proteínas al RER
Es un proceso cotraducional (simultáneamente a la síntesis de las proteínas). Intervienen las proteínas plegadoras (o chaperonas o proteínas Bip de la familia hsp 70 ) y enzimas. Tiene lugar en el lumen (por proteínas globulares) o en la membrana del RER (por proteínas transmembranales). Las proteínas Bip se separan de la proteínas una vez esta está plegada correctamente.
El tipo de enlace más frecuente en el plegamiento son los puentes de disulfuro. Los puentes S- S estabilizan la estructura tridimensional de la proteína. Estos enlaces solo se pueden formar a partir de residuos de cisteína. Se forman en el lumen del RER (ambiente oxidativo), NO al
Las glucosilaciones están presentes en la mayoría de las proteínas transportadas desde el RER al AG. Estos glúcidos siempre están orientados a la cara e (lumen de compartimentos o exterior de la célula), hay excepciones como la glucoproteína O-derivada de los poros nucleares. Las células invierten mucha energía en glucosilar por que tiene funciones muy importantes:
Proteínas de cavidad de RE, otros compartimentos del SMI y secreción celular.
El transporte a los diferentes destinos se hará a través de señales de reconocimiento específico. Hay tres medios de transporte mayoritarios:
Secreción regulada hacia el exterior Secreción constitutiva Dirección a los lisosomas
Todas aquellas proteínas que no se hayan de quedar en el RE, porque tienen su función allí, lo que hacen es ir al A.G; a través de vesículas.
Cuando salen proteínas del RE al A.G. si están muy cerca el RE y el AG, las vesículas llegan directamente al A.G. donde se fusionan. Si por el contrario, están lejos el uno del otro, aparece la vesícula tubular dubster (compartimento intermedio entre el RE y el AG), las vesículas se fusionan entre ellas y formando nuevas vesículas llegan al AG.
Las proteínas que tienen que ir o no al A.G. deben ser previamente seleccionadas.
-mecanismos de distinción:
Las proteínas que tienen que permanecer en el RE bien sea para formar parte de la membrana o como proteínas solubles disueltas en el lumen del RE, forman agregados difíciles de introducir en vesículas.
Las proteínas con la secuencia KDEL en el extremo C-terminal que se escapan del RER en una vesícula de transporte que no correspondía, vuelven de los compartimentos de Golgi a través de un mecanismo de retorno.
También lo hacen aquellas que presentan una secuencia kkxx en el extremo C-terminal, la diferencia entre las proteínas con secuencia KDEL y KKXX, ambas residentes del RE, se diferencian en que las de KDEL son proteínas residentes solubles, mientras que las KKXX son proteínas residentes de la membrana del RE.
Cuando una de las proteínas de secuencia KDEL es enviada al A.G., son reconocidas por receptores KDEL y forman nuevas vesículas que se devuelven al retículo.
Modulan las interacciones entre el receptor y el sustrato, haciendo cambiar o bien la conformación del receptor o del sustrato.