¡Descarga Señalizacion celular y más Apuntes en PDF de Biología Celular solo en Docsity!
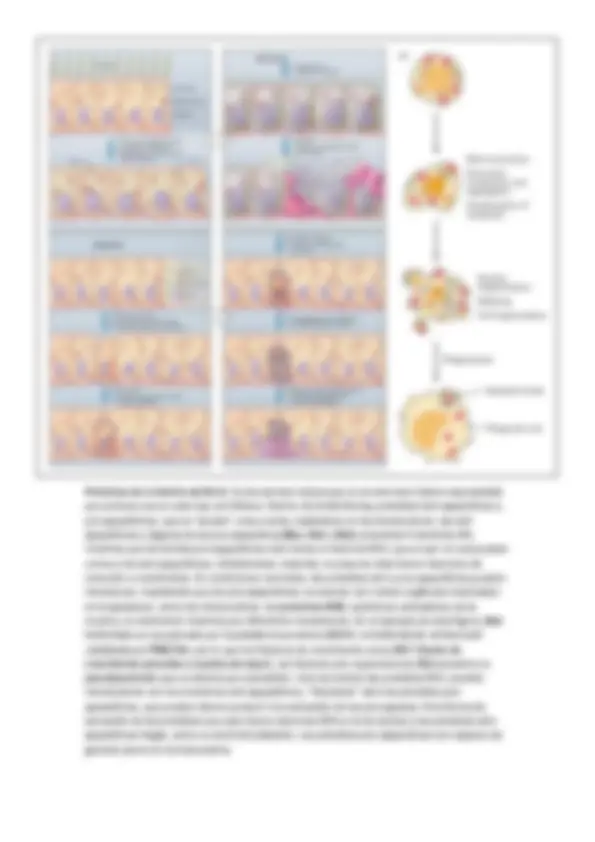
La señalización celular es la rama de la biología que estudia el mecanismo que tienen las células para responder al entorno y a los procesos intracelulares. Esto es así porque somos pura química. La señalización puede ser endógen a (respuestas intracelulares) y exógena (repuestas intercelulares). La comunicación puede ser endocrina (a larga distancia como las hormonas) o paracrina (a corta distancia como los neurotransmisores). * Turnover = vida media.
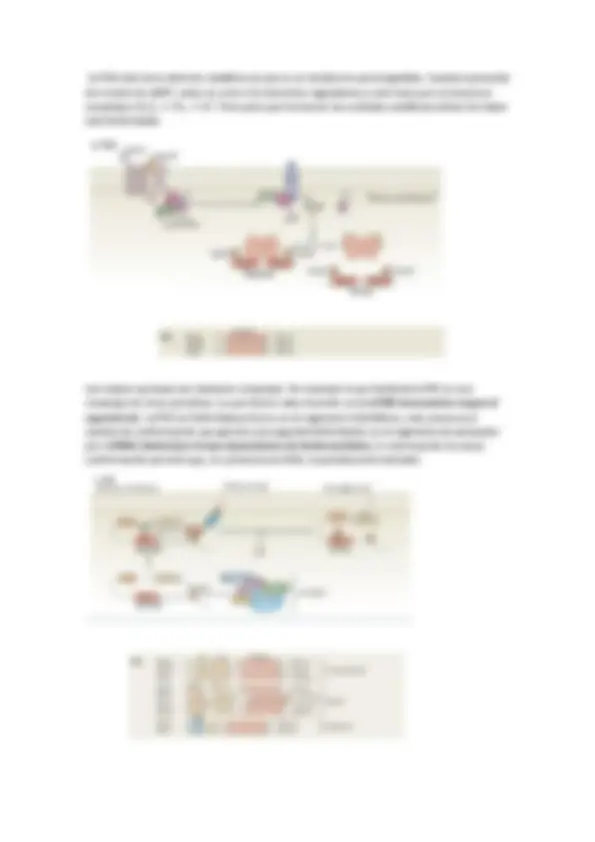
Un ejemplo de mensajero es la insulina. Esta es secretada por las células del páncreas ante un aumento del ATP producido por el incremento de concentración de glucosa en estas células. Ante este hecho, el páncreas produce la insulina que es liberada al torrente sanguíneo cuya diana son los hepatocitos y miocitos.
En muchas ocasiones hay varios mensajeros a la vez, como en el caso de la regeneración tisular, en donde aparece una inhibición por contacto (por parte de las células vecinas) y una activación de la división, mitógenos (por parte de la propia célula). En estos casos las células se guían en función de la intensidad de la señal, es decir, de la concentración de esta.
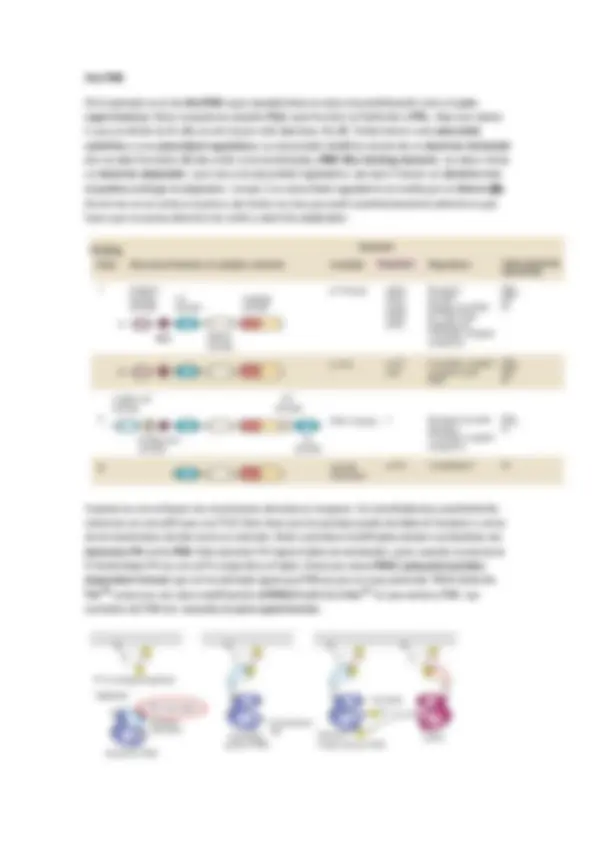
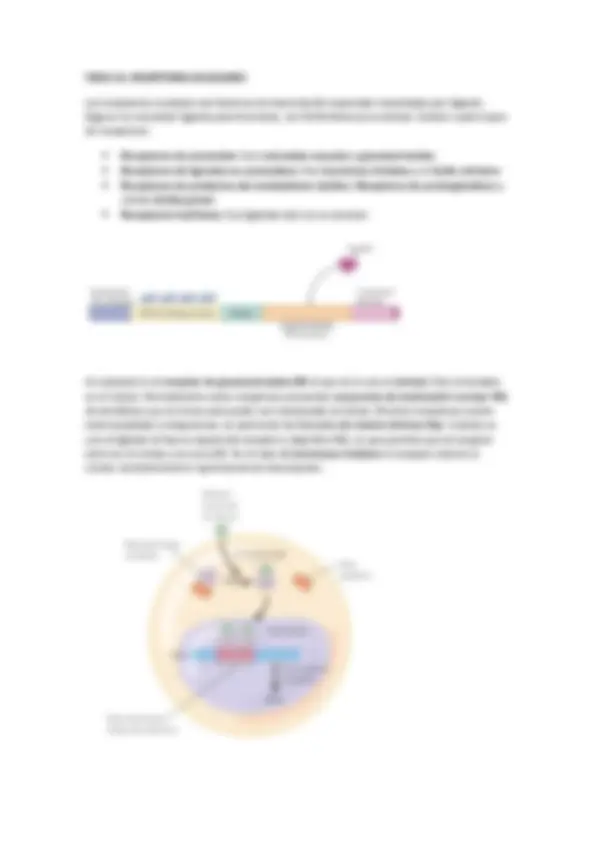
En general, una célula recibe un estímulo que genera la creación de una señal extracelular que viaja por el torrente hasta llegar a un receptor, que activa una señal, que puede ser aminoácidos, nucleótidos, lípidos, proteínas, fotones o gases. La señal llega hasta un receptor en la membrana de la célula diana, estos suelen ser canales iónicos (Na, Ca, K…), acoplados a proteínas G (PKC, AC…), con actividad enzimática intrínseca (TyrK…) o acoplados a proteínas con actividad enzimática (TNF, caspasas…). Así este receptor reacciona ante un ligando, en general, pequeño. Este ligando (la molécula señal o inhibidor o activador) puede ser agonista (si activa la señal) o antagonista (si bloquea a la señal). Si el receptor es activado provoca la aparición de una señal intracelular que suele ser un nucleótido cíclico, un lípido, [Ca2+], interacción proteína-proteína o fosforilación.
En los seres vivos pluricelulares la señalización más importante es la de las hormonas. Requiere la coordinación de múltiples señales. Las características generales de la señalización son:
Laespecificidad de un receptor. La unión del ligando es especifica del dominio de la proteína. La transducción de la señal está unido a un proceso de amplificación de la señal. La existencia de procesos en cascada tiene un efecto de amplificación que aumenta la intensidad del proceso. Así las señales iniciales suelen estar en concentraciones muy pequeñas. Integración de la información. Las señales codificadas por diferentes mensajeros regulan distintos sistemas intracelulares de modo que la respuesta vendrá determinada por el conjunto de señales que reciben
Es importante también la duración ( temporalidad ). Depende del primer componente, es decir, de la actividad del receptor: afinidad del ligando. En general, como en el caso de proteínas G, ellas mismas pueden auto modularse mediante su actividad GTPasa. Cuanto más rápida sea la hidrolisis más lenta será la duración de la señal. Si la señal intracelular mantiene una concentración alta, más larga es la duración; por lo tanto además de señal suele aparecer un elemento que degrada la señal como las fosfodiesterasas. Un ejemplo es la adrenalina , que activa el receptor -adrenérgico , este activa una proteína G que hidroliza GTP, cuando la concentración de este disminuye se activa la adenilato ciclasa (un efector). Este cicla el ATP formando AMPc. Al mismo tiempo se producen fosfodiesterasas y moduladores de este. El
aumento de [AMPc] genera la fosforilación de la PKA que fosforila diferentes sustratos. Por lo tanto también aparecen fosfatasas y moduladores de esta. También es importante la locación de los sustratos, por ejemplo la PKC fosforila sustratos de la membrana. Esta se modula por alosterismo de Ca y fosforilación/defosforilación.
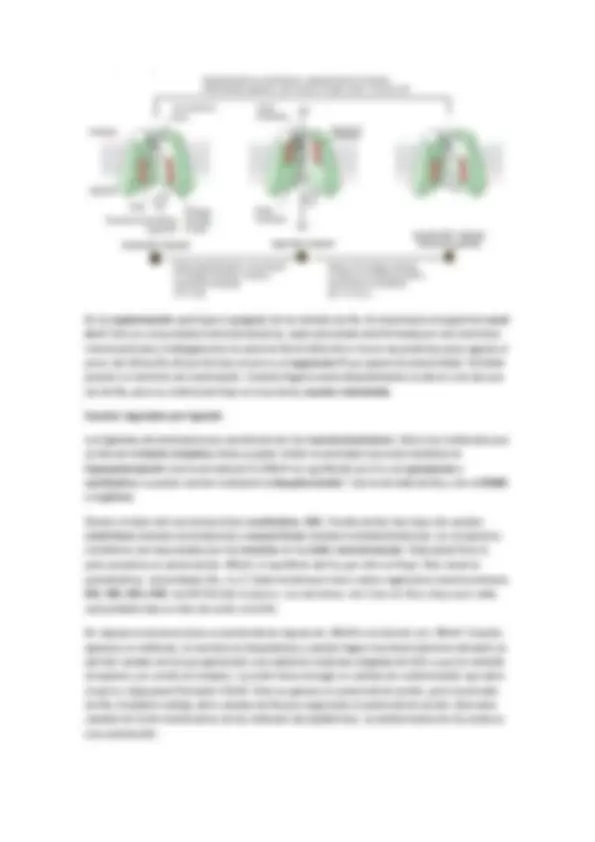
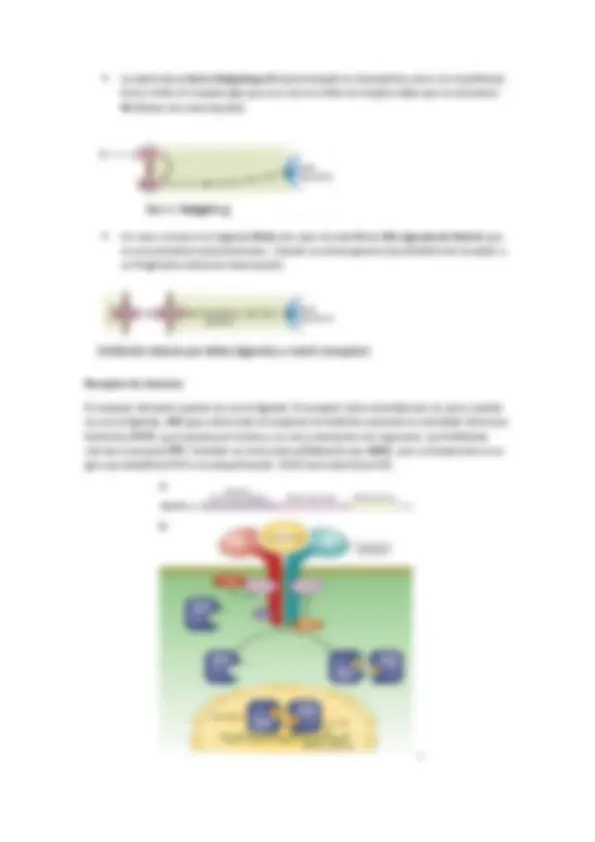
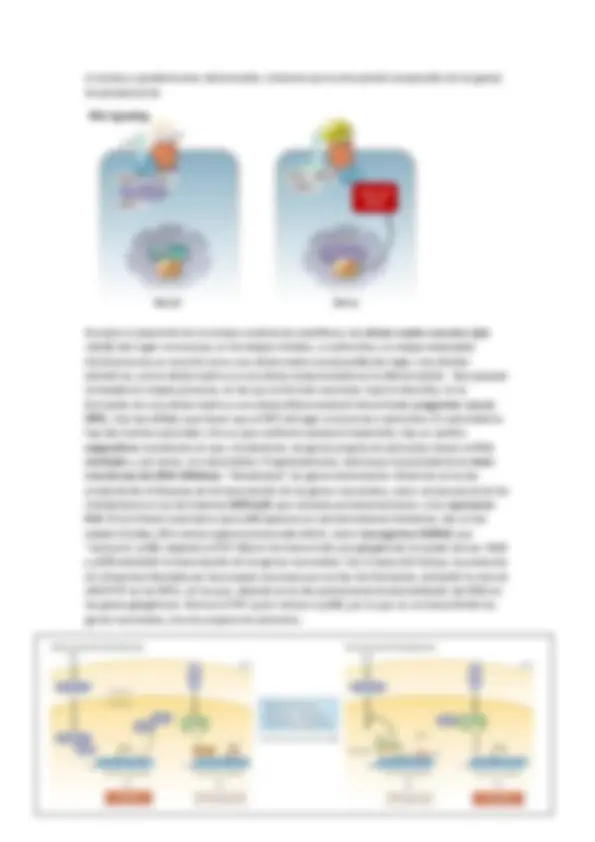
REGULACION DEL RECEPTOR : En la regulación también depende la sensibilización del receptor. La duración está relacionada con la Kd (disociación). El receptor cuando se activa también puede auto regularse mediante auto fosforilación. Son mecanismos de adaptación de modo que la activación del receptor atenúa posteriores activaciones. También puede ocurrir que la propia cascada produzca un compuesto que sea alostérico del receptor primario y puede inactivarlo o internalizarlo en la membrana y así se produce desensibilización o tolerancia.
REGULACIÓN DE LA ACTIVIDAD ENZIMÁTICA : Pueden ser:
A corto plazo : Son procesos rápidos (de segundos a minutos) y suelen cambiar la actividad de las moléculas pre-existentes. Ejemplos de estos mecanismos son: Alosterismo : Regulación a través de un lugar diferente al centro activo. La unión del ligando puede provocar una sobre estimulación de la proteína o una inactivación. Ejemplos son la calmodulina con el Ca2+^ y los dedos de Zn2+ Modificación covalente reversible : Aquí entra la fosforilación/desfosforilación por parte de PKs y PPasas. Es un proceso dinámico y sensible. Muchas veces aparece convergencia en la regulación por diferentes rutas. Actúan sobre aminoácidos concretos (tirosina, treonina y serina) dirigidos por secuencias específicas consenso. Interacción proteína-proteína : En general inducida por procesos alostéricos o de fosforilación. Suele ser la formación de agregados como es el caso de la ACC. A largo plazo : Son procesos lentos (de horas a días) y suelen cambiar los niveles de enzima. La regulación de los niveles de proteína/mensajero se da en: Cambios de la expresión génica y en la vida media proteínas (turnover). La regulación de la expresión génica es esencial en: Desarrollo tejidos y órganos y respuesta a factores ambientales/extracelular. La vida media de una enzima se define como la diferencia entre la degradación en el proteasoma y su transcripción y posterior traducción.
Otro ejemplo de la regulación es la presencia de isoformas , que son proteínas con las mismas propiedades que proceden de distintos genes. Al ser en general agregados son más fácilmente regulables. Las distintas isoformas difieren en la afinidad y condicionan el distinto funcionamiento celular. Un ejemplo es la LDH que difiere en los tejidos astrocitos-neurona o fibras blancas-corazón. La Km de la M 4 es menor frente al piruvato mientras que la H 4 es mayor.
NATURALEZA DE LOS MENSAJEROS QUÍMICOS : Los segundos mensajeros son de seis tipos:
Aminoácidos : Como el ácido glutámico o la glicina. Nucleótidos cíclicos : Como GMPc o AMPc Lípidos : Como las PGs o tromboxanos. Cationes : Como Ca2+^ o K+ Aminoácidos modificados : Como el GABA o la serotonina. Gases : Como el NO
Las subunidades también tienen funciones como la regulación de canales iónicos o la regulación del tiempo de duración mediante activación GRKs ( G receptor kinase ) que fosforila GPCRs que genera una disminución de la afinidad del receptor por el ligando o puede que active la endocitosis del receptor (recordad que el N-terminal estaba hacia fuera de la célula y ahora estará en el lumen de la vesícula). En general la función es la de desensibilización.
Existen unos 35 genes que codifican para proteínas G:
: 16 genes en cuatro tipos de s y tres tipos de i : 5 genes : 14 genes
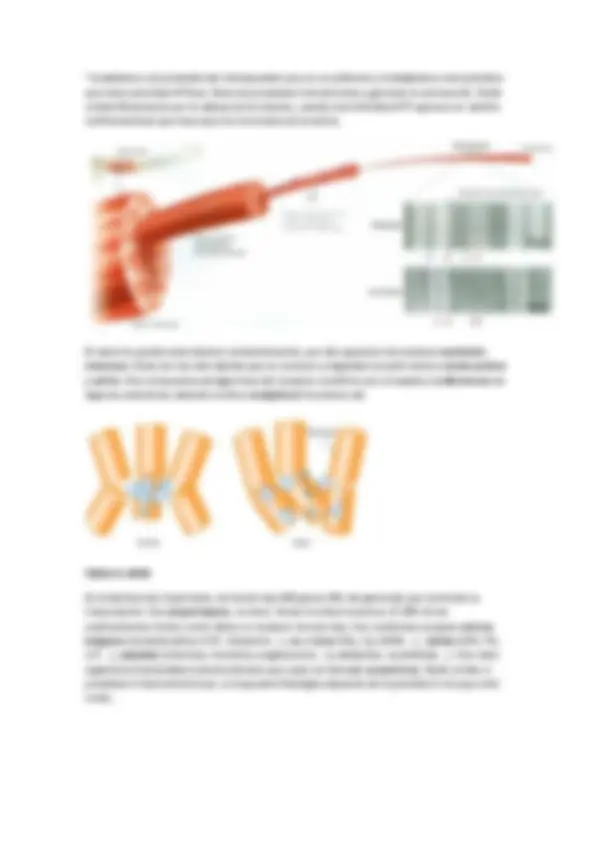
La asociación entre diferentes tipos de cadenas se define en el tipo de regulación que se necesita. Existe un denominado lazo que está oculto en la estructura , cuando se disocia, el lazo queda expuesto y por ello comienza la cascada. La actividad de la señal depende directamente sobre el tipo de proteína G este unida. Por ejemplo los receptores de las catecolaminas que en función de la proteína G tienen una función u otra: 1 +q (aumenta a concentración de Ca2+)
Existen análogos del GTP como el GppNHp ( guanosina-5’-( -imino)trifosfato )que no es hidrolizable, lo que genera es que la proteína G este constitutivamente activada. Otro ejemplo es el GTP- -S^35 ( guanosina-5’-O-3-tiotrifosfato ) que nos permite ver por autorradiografía que partes de un tejido son activadas. Muchas bacterias producen toxinas que afectan a proteínas G mediante ADP ribosilación : un NAD+ se une a una arginina de la subunidad S, soltando nicotinamida. La toxina colérica produce en el intestino una diarrea mortal mediante un flujo continuo de iones cloro y la toxina pertussis ( tos ferina ) ADP ribosila a una i generando una inactivación constitutiva que provoca una deficiencia pulmonar.
El tiempo de duración depende del tiempo que esté unido el ligando al receptor, es decir, el tiempo que esté activo la GPCRs más la actuación de los GEFs. La actividad intrínseca GTPasa se regula por GAP. Un ejemplo es la regulación de la actividad PKA : La proteína G activa la AC que genera AMPc que se une a las dos unidades reguladoras de PKA, que provoca su disociación, esto hace que las dos subunidades catalíticas fosforilen proteínas. Las subunidades catalíticas también activan las PDE que generan una disminución de la señal. Una vía de la PKA es la regulación de la expresión génica (transcripción) mediante la modificación covalente de la
CREBP. Cuando aumenta el cAMP el cofactor CBP se une al factor de transcripción
CREBP que hace que el elemento de respuesta CRE ( cAMP response element ) se una
al DNA haciendo que se transcribe un gen ya que es una acetilasa.
Las proteínas G monoméricas.
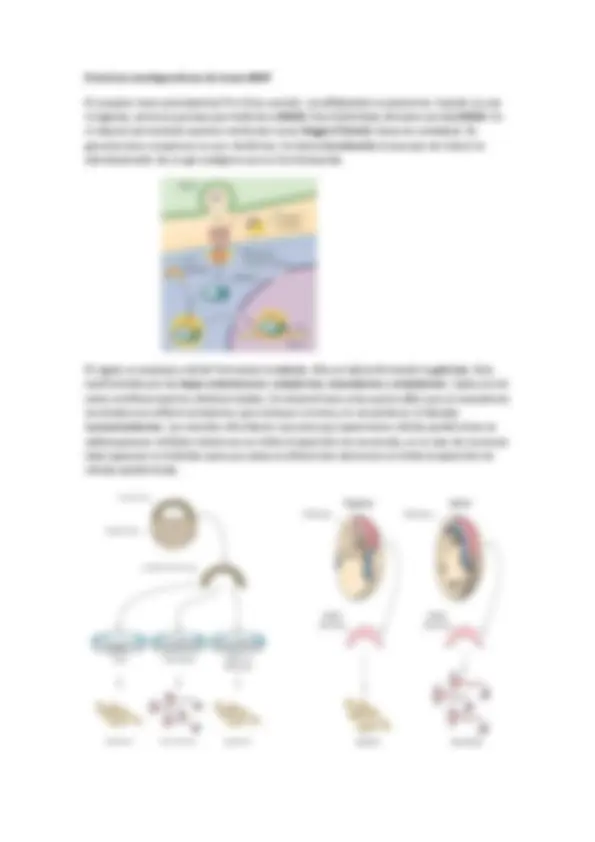
Mariano Barbacid descubrió el primer protooncogen: Ras , que tiene tres familias: H-Ras , N- Ras (acilada y prenilada) y K-Ras (prenilada y dominio polibásico). Las proteínas G monoméricas se unen a la membrana por vía farnesilo. Solo tienen una cadena y es anterior evolutivamente a las heterotriméricas. Oscila su peso alrededor de 21kDa.
Su activación depende de receptores TyrK (un único segmento transmembrana y en general receptan factores de crecimiento , GF como el NGF , EGF , PDGF ). Se basa en un dímero con cuatro partes, dos hacia adentro y dos hacia afuera. Cuando la señal llega al receptor, este cambia de conformación, se autofosforila sus propias tirosinas. A continuación, esto genera que se abra un dominio catalítico que fosforila a Grb2, que le permite la unión a Sos. Sos es un GEF de Ras.
LÍPIDOS Y SEÑALIZACIÓN CELULAR
Los más importantes son los glicerofosfolípidos y esfingolípidos. Su señalización se basa en hidrólisis: fosfolipasas y PLK/PLPasa.
Los PL están formados por un alcohol, el glicerol con tres grupos hidroxilo. A estos están esterificados dos ácidos grasos, uno saturado y otro insaturado; el tercer hidroxilo esta esterificado a un fosfato (carga negativa) y este unido a un compuesto alcohólico como colina , etanolamina o inositol. Hay tres tipos de PLPasas:
PLA 1 : Forma un ácido graso saturado y un lisofosfolípido , LPL (un PL con un solo AG) PLA 2 : Forma un ácido grado insaturado y un lisofosfolípido. En general en señalización se forma ácido araquidónico , 20:4 , AA PLC : Forma diacilglicero l, DAG y un alcohol fosfato. PLD : Forma ácido fosfatídico , PA y el alcohol.
El fosfatidilinositol , PI es un PL cuyo alcohol es el inositol , que es un hexanohexanol, que es fácilmente fosforilable, sobre todo en las posiciones 4 y 5: PI4,5bisP o PI4,5P 2. Si sobre este
compuesto actúa PLC, se forma el DAG e IP 3. PIP 2 →DAG+IP 3
Tanto los PL como el colesterol están situados asimétricamente en la membrana (tanto
fuera como dentro), y además también difieren las membranas internas. Por ejemplo
la membrana de los eritrocitos se puede lisar emulsionando colesterol sin dañar la
membrana de los orgánulos. Las señales derivadas de fosfolípidos suelen actuar
mediante lípido oxigenanas , como COX de las prostaglandinas.
Fosfolipasa C
La PLC presenta varios dominios:
PH : Es un dominio de homología plecstrina que une a PI y es regulable por EF-Hands : Es un dominio de unión de Ca2+. Puede ser de inhibición o activación. XY : De actividad catalítica CRD ( Cys Rich Domain ) - C 2 : Dominio de unión a lípidos con calcio como PS o DAG
Un ejemplo de señal mediada por PLC es la ruptura del PIP 2 mediante PI-PLC. Esto va a liberar DAG, que al ser soluble en la membrana difundirá por ahí y activará la PKC (se transloque desde el citoplasma a la membrana) e IP 3 que difundirá al citoplasma y movilizará Ca2+ mediante activación de canales iónicos. También se activa esta vía mediante receptores
acoplados a proteínas Gq.
La PKC tiene dos dominios: C 1 de unión a DAG y C 2 de unión a PS/Ca2+^ y se regulan por fosforilación. Los ésteresde forbol son moléculas de síntesis de industrial análogos a DAG. En su estado inactivo están los dominios catalítico y de pseudosustrato (ya que el sustrato también es una parte de la proteína, que es la le bloquea) bloqueados, cuando se fosforilan sus aa y en presencia de calcio se une a la membrana y deja sus dominios catalíticos abiertos. Al igual que Ras, es un protooncogen.
Fosfolipasa D
La PLD forma PA y el alcohol. Sin embargo aún no se sabe muy bien la función del PA. La verdadera función es la de participar en las modificaciones de membrana como formación de vesículas y la internalización de receptores. Este hecho lo descubrieron Sudhof , Rothman y Schekman que ganaron el Nobel en 2013 (‘Trafico vesicular’)
LÍPIDO KINASAS Y FOSFATASAS. PLK Y PLPasas
En particular nos fijaremos en las que actúan sobre el inositol. Un ejemplo de PIPasa es la que produce PI 4 P.
Una PIK importante es la PI 3 K que forma PI3,4,5P 3 (que se ve afectada por señales). El efector de esta señal es la PKB que actúa sobre la proliferación y la supervivencia celular, sin embargo esta es activada previamente por una PDK1. Para regular esta cascada aparece PTEN (protooncogen) que es una fosfatasa. Hay tres clases de PIK pero nos interesan las de clase I. Se dice ‘ pool ’ de algo cuando nos referimos al lugar donde está mayoritariamente.
Dos subunidades: Una catalítica y otra reguladora
Catalítica : Dominio catalítico, dominio C 2 , dominio helicoidal (no se sabe para qué), dominio de unión a Ras (Ras-GTP) y de unión a la p85 (subunidad reguladora). Puede fosforilar a la unidad catalítica. Reguladora : Dominios SH2 y SH3 (de tirosinas fosforiladas) y de unión a p (subunidad catalítica).
También presentan dominios PH.
CANALES IÓNICOS
Los iones son moléculas muy pequeñas con carga. La membrana genera un gradiente electroquímico gracias a una permeabilidad selectiva. Los sistemas de transporte son:
Difusión simple Facilitada (proteínas facilitadoras a favor de gradiente) Transporte activo (acoplado a hidrólisis de ATP en contra de gradiente. Son bombas) Transporte secundario (dos solutos)
Existe una diferencia de concentración de iones a ambos lados de la membrana. El sodio es unas doce veces mayor concentración fuera que dentro, el potasio unas diez veces menor fuera que dentro, el calcio unas 1500 veces mayor fuera. Esta diferencia genera un potencial.
En relación con la señalización tenemos: una concentración en reposo (sin actividad) que gracias a un ‘mecanismo de encendido’ se cambie la concentración a más o menos. Una vez tengamos esta situación también hay un ‘mecanismo de apagado’.
DIFUSIÓN FACILITADA
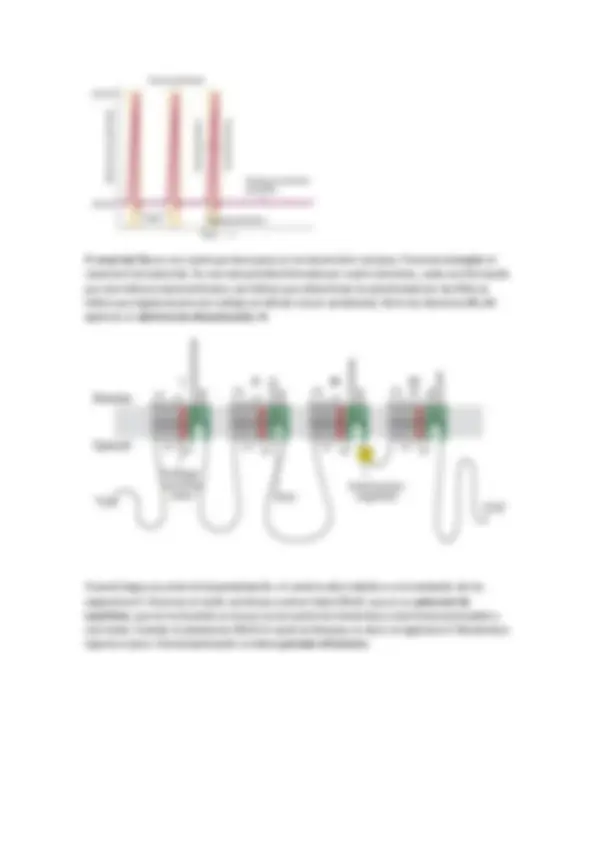
Se explicara mediante el ejemplo del canal de K. Si se abre este canal el potasio sale de la célula. Estos canales están formados por hélices anfipáticas (aa polares hacia dentro de esta y aa hidrofóbicos frente a la membrana), el interior de la hélice es lógicamente hidrofílico ya que por dentro pasa el ion. Estos canales son selectivos. El transporte suele ser no saturable, ya que el canal cuando se abre deja pasar a los iones indefinidamente, la apertura y cierre es lo regulado. Hay una seria de aa que interaccionan con el ion, lo que delimita su entrada es el estado de hidratación de este.
Cando se introduce en el poro debe de deshidratarse, los aa mimetizan la longitud del puente de hidrógeno para que la energía de solvatación sea mínima. Si introdujésemos un ion distinto, aunque tenga la misma carga, el distinto tamaño no permitiría su entrada (aunque sea más pequeño). El balance de energía debe ser positivo y además, en definitiva, la interacción depende del radio iónico.
La velocidad de transporte depende del tiempo en que esté abierto el poro. Cuando entra un primer potasio la velocidad de entrada es ligeramente pequeña, pero al introducirse el segundo aparece una repulsión electrostática y así aumente la velocidad.
Los mecanismos de disipación del poro están regulados, existen tres tipos:
ROC: Canales iónicos operados por ligando extracelular (receptor) como neurotransmisores VOC: Canales iónicos operados por voltaje. Se activan mediante diferencias de voltaje, es decir, canales iónicos activan otros canales iónicos. SMOC : Canales iónicos operados por ligando intracelular (segundos mensajeros) como Ca, AA, cAMP …
Un ejemplo es el impulso nervioso. La neurona pre sináptica genera una onda de voltaje que en la membrana pre sináptica (axón) genera una salida de un neurotransmisor hacia la hendidura sináptica ; este se une a receptores de la membrana post sináptica (dendrita) que genera otra onda, y así continúa el potencial de acción.
CANAL DE POTASIO. El canal de potasio también tiene 6 hélices transmembrana cada domino (cuatro): dos que forman el canal y uno regulador. Todo el mecanismo se basa en las interacciones entren aa con aa y aa con la membrana. Estos cambios generan distintas conformaciones ya que hay hélices que físicamente se mueven en el espacio.
ATPasa. Cuando ha pasado un impulso la célula debe reconcentrar los iones nuevamente. En esta situación actúan las ATPasas de tipo P. Poseen cuatro dominios:
Dominio fosforilado P , que curiosamente en esta familia es el Asp Domino M (transmembrana) Dominio A (actuador) Dominio N (unión a ATP)
Se gasta ATP para translocar iones. En resumen su movimiento se basa en que se unen los iones de la zona poco concentrada y el cambio de conformación gire la proteína, cuando gira y los suelta cerrarse para que no entren. La ATPasa de Ca2+^ del RE también funciona así. Tiene los mismos dominios que la Na+-K+^ ATPasa.
CALCIO
El calcio es un modulador muy importante ya que la concentración en el interior de la célula es muy, muy pequeña: 100nM. Tenemos, al igual que los demás iones, mecanismos de apagado y de encendido.
La concentración de Ca se regula mediante ATPasas. Hay dos familias:
PMCA ( plasma membrane calcium ATPase ) De orgánulos: SERCA ( sarcoplasmic endoplasmic reticulum calcium ATPase ) y NCX (mitocondrial y de membrana, pero su función es reguladora de su propio metabolismo: buffer)
Es curioso que la ATPasa uniportador de Ca tiene mayor afinidad y menor capacidad de transporte que el intercambiador Ca-Na. Esto es así porque la función del uniportador es simplemente de ajuste 'fino' de concentraciones. El RE es el orgánulo de acumulación, con una concentración de 100M
Los mecanismos de encendido se basan en la apertura de canales o de membrana o del RE:
Receptores metabotrópicos. Aquellos que actúan mediante proteínas G. Son los que generan segundos mensajeros como son IP 3 o cADPR Receptores ionotrópicos. Aquellos que son canales iónicos.
IP 3. PLC rompe PIP 2 para dar DAG e IP 3. Este segundo mensajero se une a un receptor de la membrana del RE que provocará la salida desde el lumen de Ca. SERCA es una proteína formada por cuatro dominos, cada dominios de seis hélices transmembrana. También une calcio tanto por la cara externa como por la interna. Tiene centros de inhibición y centros de activación en la cara citoplasmática. Cuando [Ca] es baja, aparece un sitio de alta afinidad de efecto activatorio; cuando [Ca] aumenta, este se une a un sitio de baja afinidad de efecto inhibitorio. En la cara lumínica del retículo también hay dominios de unión de Ca, cuya función es la de regular y controlar [Ca].
cADPR/NAADP. Se conocen menos. Se vió que en celulas modificadas para que no se formara IP3, las células podían regular el Ca. El primero que se observó fue el cADPR (ADP ribosil cíclico) formado por la ADP ribosil ciclasa liberando nicotinamida. El mecanismo de activación de la enzima no se conoce, pero se sabe que este compuesto una bomba de Ca del RE distinta a SERCA, RYR. Se llamó así porque sin saber porque la toxina rianodina (alcaloide de origen vegetal) actuaba ahí. Lo mismo pasa con el NAADP (nicotin ADP, la niacina no está acetilada).
CANALES.
ROC. Estos canales se basan en que cuando entran en contacto con alguna señal, el canal físicamente o se abre o se cierra. VOC. La diferencia de potencial de membrana abre o cierra estos canales. Cuando se empezaron a estudiar los canales se observó que dependiendo de tejido aparecían distintas corrientes de calcio. Estos son CaV 1 o tipo L (músculo, corazón, riñón ...), CaV 2 o tipo P/Q (riñón, SNC, pituitaria ...) y Cav 3 o tipo T (hígado, cerebro, riñón ...).
En el caso del músculo esquelético , más coordinado que en el cardíaco, también se activan por calcio. Las fibras colinérgicas procedentes de la médula activan mediante receptores que en realidad no genera entrada de Ca sino que su cambio conformacional abre los canales de rianodina del RE (están unidas físicamente estas membranas).
SMOC.
SOCS ( Store-Opertated calcium channels ). Se observó inhibiendo mediante una toxina la ATPasa de Ca del RE que entraba calcio desde fuera (ya que se había vaciado el RE). No se conoce realmente el mecanismo de señalización aún. Los canales son de la familia TRP (receptores valinoide de los picantes)
En la señalización mediada por calcio aparecen dos tipos de proteínas: proteínas tamponadoras (unen calcio pero median poco la concentración) y proteínas efectoras (unen calcio pero no median nada la concentración, la mitocondria es rica en ellas).
CALMODULINA
Presenta cuatro dominios EF-Hands que son de unión a calcio. Se llama mano EF ya que la dirección de las cadenas simbolizan la posición relativa de los dedos de la mano. Cuando une calcio la proteína se pliega y puede así activar otras proteínas diana mediante hélices, normalmente. Así el mensajero no es el calcio sino la calcio-calmodulina.
TEMA 5. PAPEL DE LOS GASES EN SEÑALIZACIÓN
El óxido nítrico contribuye a la vasodilatación, Premio Nobel en 1998 a Robert F. Furchgott , Louis J. Ignarro y Ferid Murad. Este equipo estaba estudiando el papel de la acetilcolina en la contracción del músculo esquelético. Cogieron un fragmento de vaso sanguíneo y midieron la tensión del músculo liso: al añadir la acetilcolina bajaba la tensión y la noradrenalina subía la tensión. En realidad no sabían en qué células actuaban estas moléculas. Cuando rascaron el endotelio los resultados cambiaban, entonces supusieron que estas producían algo para relajar el músculo: factor de relajación derivado del endotelio EDRF.
A Louis J. Ignarro se le encendió la bombilla y pensó que debía ser un gas que difundiera bien entre las membranas. En el experimento utilizó una proteína que uniera gases: la hemoglobina. Al analizarla espectroscópicamente vieron que el gas que cuadraba era el óxido nítrico. Así la acetilcolina se une al endotelio, este produce el EDRF que genera la relajación del tejido adyacente, el músculo liso.
La producción de este gas deriva del ciclo de la urea. La arginina se convierte en N-hidroxil- arginina. Este mediante la NO sintasa se convierte el L-citrulina liberando NO. Es una reacción complicada.
Esta enzima utiliza varios cofactores: FAD , FMN , tetrabiopterina , Ca/CM ... Tiene un grupo hemo y utiliza O 2. Algunas isoformas se regulan por Ca-calmodulina. La enzima presenta dos dominios: dominio de reducción y dominio oxigenasa.
Hay tres isoformas NOS-1 o neuronal , NOS-2 o inducible y NOS-3 o endotelial que se diferencian en su regulación, dominios de unión, genes ... Todas comparten el grupo hemos.
En bajas concentraciones es un segundo mensajero, pero en altas concentraciones es tóxico, de hecho algunas células del sistema inmune lo producen como biotóxico.
GUANILATO CICLASA
La acción de la NOS III se basa en la activación de proteínas que generan segundos mensajeros. Su producción se mide mediante calcio. La acetilcolina activa la PLC que genera una salida de Ca del RE, este se une a la calmodulina por su dominio EF, este activada se activa la NOS. El NO se une a la guanilato ciclasa. Este enzima es análogo a la AC y produce cGMP. El lugar de reconocimiento del NO es otro grupo hemo, al unirse al ligando esta estructura 'tira' de una hélice activando así a la proteína.
Existen dos tipos: guanilato ciclasa de membrana y guanilato ciclasa soluble. Esta última es la que responde a la regulación por NO y se sitúa en el citoplasma. La proteína tiene un dominio catalítico que cicla GTP y un dominio de regulación por NO. A continuación el cGMP se une a una kinasa, PKG ( proteína kinasa dependiente de cGMP ). La PKG es la que genera la relajación del músculo. El músculo liso también se contrae por calcio y por una proteína que se denomina MLCK ( kinasa de la cadena ligera de la miosina ). Esta proteína, regulada por Ca, fosforila la miosina y esta se contrae, contrayendo así el músculo. La PKG inhiben o [Ca] o MLCK.