



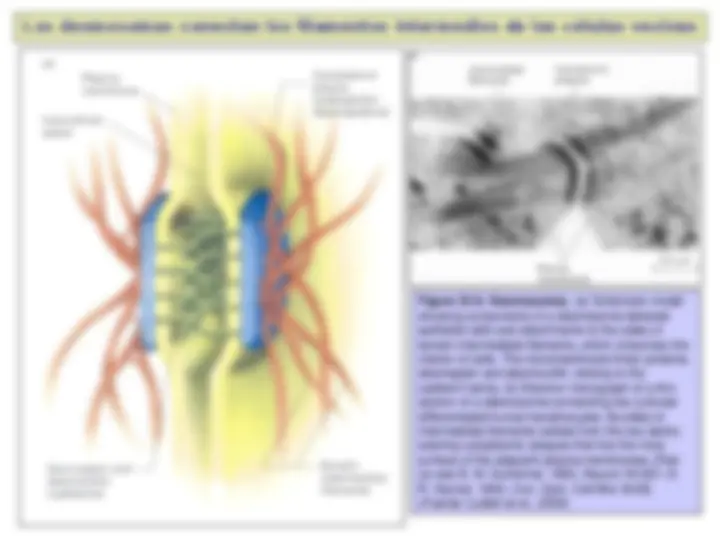
Esquema que muestra los diversos
tipos de uniones intercelulares. Basado
en el epitelio intestinal. (Fuente: Alberts et al., 2008)
LAS UNIONES ENTRE CÉLULAS SON DE TRES
TIPOS FUNDAMENTALES














Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Estructura de la celula, Profesor: , Carrera: Biologia, Universidad: UV
Tipo: Apuntes
1 / 32

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
en el epitelio intestinal. (Fuente: Alberts et al., 2008)
Figure 19-26. Three mechanisms by which cell- surface molecules can mediate cell-cell adhesion. Although all of these mechanisms can operate in animals, the one that depends on an extracellular linker molecule seems to be least common. (Fuente: Alberts et al., 1993)
Figure 22-2. Major families of cell-adhesion molecules (CAMs). Integral membrane proteins are built of multiple domains. Cadherin and the immunoglobulin (Ig) superfamily of CAMs mediate homophilic cell-cell adhesion. For cadherin, calcium binding to sites (orange) between the five domains in the extracellular segment is necessary for cell adhesion; the N-terminal domain (blue) causes cadherin to dimerize and to bind cadherin dimers from the opposite membrane. The Ig superfamily contains multiple domains (green) similar in structure to immunoglobulins and frequently contain type III fibronectin repeats (purple). In a heterophilic interaction, the lectin domain of selectins binds carbohydrate chains in mucin-like CAMs on adjacent cells in the presence of Ca2+. The lectin domain is separated from the membrane by a series of repeated domains. The major cell-matrix adhesion molecule, integrin, is a heterodimer of α and β subunits. They bind to the cell- binding domain of fibronectin, laminin, or other matrix molecules. (Fuente: Lodish et al., 2000)
Figure 22-6. Desmosomes. (a) Schematic model showing components of a desmosome between epithelial cells and attachments to the sides of keratin intermediate filaments, which crisscross the interior of cells. The transmembrane linker proteins, desmoglein and desmocollin, belong to the cadherin family. (b) Electron micrograph of a thin section of a desmosome connecting two cultured differentiated human keratinocytes. Bundles of intermediate filaments radiate from the two darkly staining cytoplasmic plaques that line the inner surface of the adjacent plasma membranes. [Part (a) see B. M. Gumbiner, 1993, Neuron 11: 551; D. R. Garrod, 1993, Curr. Opin. Cell Biol. 5: 30] (Fuente: Lodish et al., 2000)
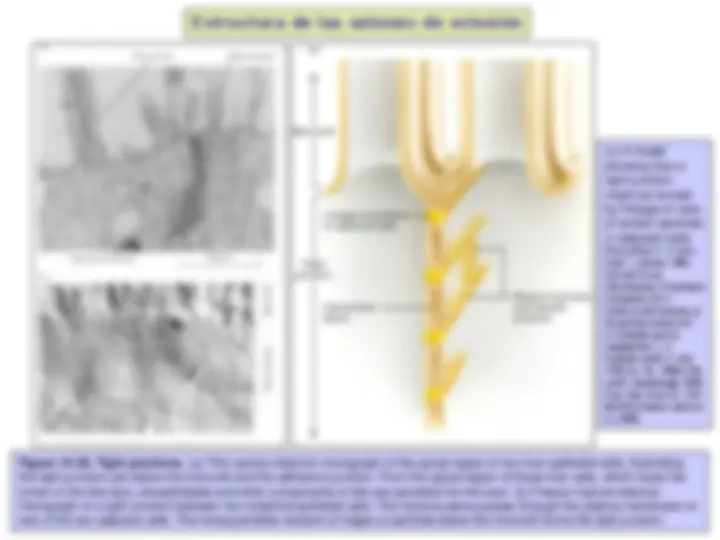
Figure 19-14. The linkages of classical cadherins to actin filaments. The cadherins are coupled indirectly to actin filaments via beta-catenin and other anchor proteins. Alfa-catenin, vinculin and plakoglobin (also called gamma- catenin) are probably also present in the linkage or involved in control of its assembly, but the details of the anchorage are not well understood. Another intracellular protein, called p-120 catenin, also binds to the cadherin cytoplasmic tail and regulates cadherin function. Beta-catenin has a second, and very important, function in intracellular signalling. (Fuente: Alberts et al., 2008).
Molecule Predominant Cellular Distribution
E-cadherin Preimplantation embryos, non-neural epithelial tissue
P-cadherin Trophoblast (placenta)
N-cadherin Nervous system, lens, cardiac and skeletal muscle
SOURCE: M. Takeichi, 1988, Development 102: 639; M. Takeichi, 1991, Science 251: 1451; H. Inuzuka et al., 1991, Neuron 7: 69; and M. Donalies et al., 1991, Proc. Nat'l. Acad. Sci. USA 88: 8024.
Figure 10-42. The protein-carbohydrate interaction that initiates the transient adhesion of neutrophils to endothelial cells at sites of inflammation. (A) The lectin domain of P-selectin binds to the specific oligosaccharide shown in (B), which is present on both cell-surface glycoprotein and glycolipid molecules. The lectin domain of the selectins is homologous to lectin domains found on many other carbohydrate-binding proteins in animals; because the binding to their specific sugar ligand requires extracellular Ca2+, they are called C-type lectins. A three-dimensional structure of one of these lectin domains, determined by x-ray crystallography, is shown in (C); its bound sugar is colored blue. Gal = galactose; GlcNAc = N - acetylglucosamine; Fuc = fucose; NANA = sialic acid. (Fuente: Alberts et al., 1993)
Figure 22-4. Interactions between cell-adhesion molecules during the initial binding and tight binding of T cells, a kind of leukocyte, to activation endothelial cells. Once a T cell has firmly adhered to the endothelium, it can move (extravasate) into the underlying tissue. Activation of the endothelium requires signals, such as platelet-activating factor (PAF), that are released in areas of infection or inflammation; thus extravasation occurs only in such areas. [Adapted from R. O. Hynes and A. Lander, 1992, Cell 68: 303.] (Fuente: Lodish et al., 2000)
Figure 15-28. Tight junctions. (a) Thin-section electron micrograph of the apical region of two liver epithelial cells, illustrating the tight junction just below the microvilli and the adherens junction. From the apical region of these liver cells, which faces the lumen of the bile duct, phospholipids and other components of bile are secreted into the duct. (b) Freeze-fracture electron micrograph of a tight junction between two intestinal epithelial cells. The fracture plane passes through the plasma membrane of one of the two adjacent cells. The honeycomblike network of ridges of particles below the microvilli forms the tight junction.
(c) A model showing how a tight junction might be formed by linkage of rows of protein particles in adjacent cells. [Part (a) from P. A. Cross and K. L. Mercer, 1993, Cell and Tissue Ultrastructure, A Functional Perspective, W. H. Freeman and Company, p. 50; part (b) courtesy of L. A. Staehelin; part (c) adapted from L. A. Staehelin and B. E. Hull, 1978, Sci. Am. 238(5):140, and D. Goodenough, 1999, Proc. Natl. Acad. Sci. USA 96:319.] (Fuente: Lodish et al., 2000)
Figure 22-8. Structure of gap junctions. (a) In this model, a gap junction is a cluster of channels between two plasma membranes that are separated by a gap of about 2
Figure 21-35. An electric synapse. (a) The plasma membranes of the presynaptic and postsynaptic cells are linked by gap junctions. Flow of ions through these channels allows electric impulses to be transmitted directly from one cell to the other. (b) Negatively stained, electron microscopic image of the cytosolic face of a region of plasma membrane enriched in gap junctions; each “doughnut” forms a channel connecting two cells. [Part (b) courtesy of N. Gilula.]
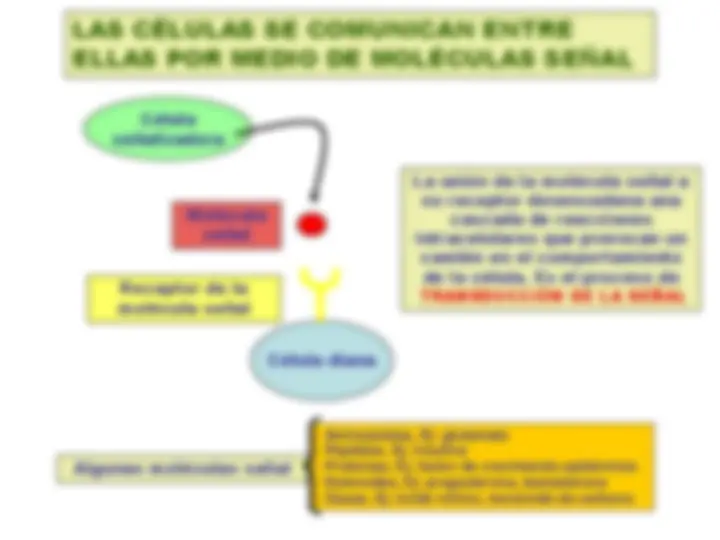
Figure 20-1. General schemes of intercellular signaling in animals. (a – c) Cell-to-cell signaling by extracellular chemicals occurs over distances from a few micrometers in autocrine and paracrine signaling to several meters in endocrine signaling. (d) Proteins attached to the plasma membrane of one cell can interact directly with receptors on an adjacent cell. (Fuente: Lodish et al., 2000)
Señalización endocrina : células
diana a larga distancia
paracrina : células
diana adyacentes
Señalización autocrina : la
misma célula señalizadora es célula diana
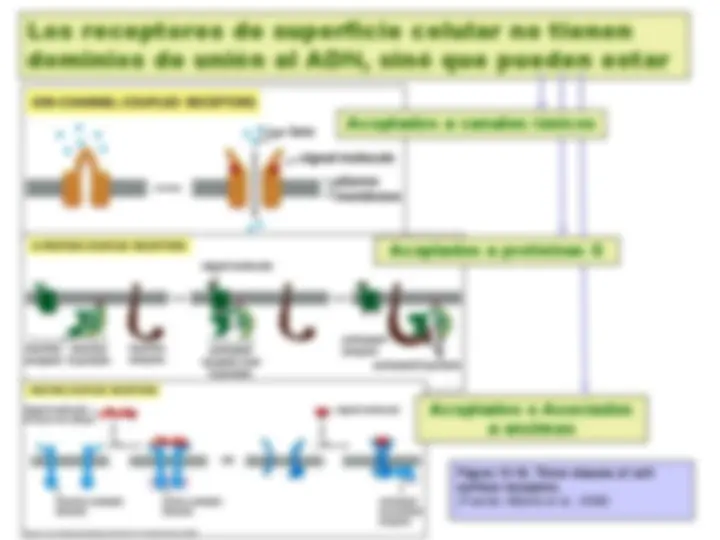
Figure 15-3. Extracellular signaling molecules bind to either cell-surface receptors or intracellular receptors. Most signaling molecules are hydrophilic and are therefore unable to cross the plasma membrane directly; instead, they bind to cell-surface receptors, which in turn generate one or more signals inside the target cell. Some small signaling molecules, by contrast, diffuse across the plasma membrane and bind to receptors inside the target celleither in the cytosol or in the nucleus (as shown). Many of these small signaling molecules are hydrophobic and nearly insoluble in aqueous solutions; they are therefore transported in the bloodstream and other extracellular fluids bound to carrier proteins, from which they dissociate before entering the target cell. (Fuente: Alberts et al., 2008)
Figure 15-12. The intracellular receptor superfamily. (A) A model of an intracellular receptor protein. In its inactive state the receptor is bound to an inhibitory protein complex that contains a heat-shock protein called Hsp90. The binding of ligand to the receptor causes the inhibitory complex to dissociate, thereby activating the receptor by exposing its DNA- binding site. The model shown is based on the receptor for cortisol, but all of the receptors in this superfamily have a related structure, as shown in (B), where the short DNA-binding domain in each receptor is shown in green. Domain-swap experiments suggest that many of the hormone-binding, transcription-activating, and DNA-binding domains in these receptors can function as interchangeable modules. It is thought that all of the intracellular receptor proteins bind to DNA as either homodimers or heterodimers (Fuente: Alberts et al., 1993)
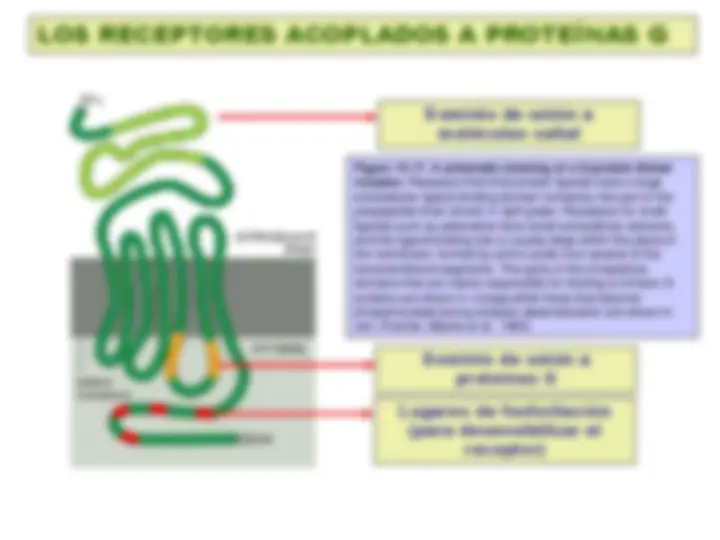
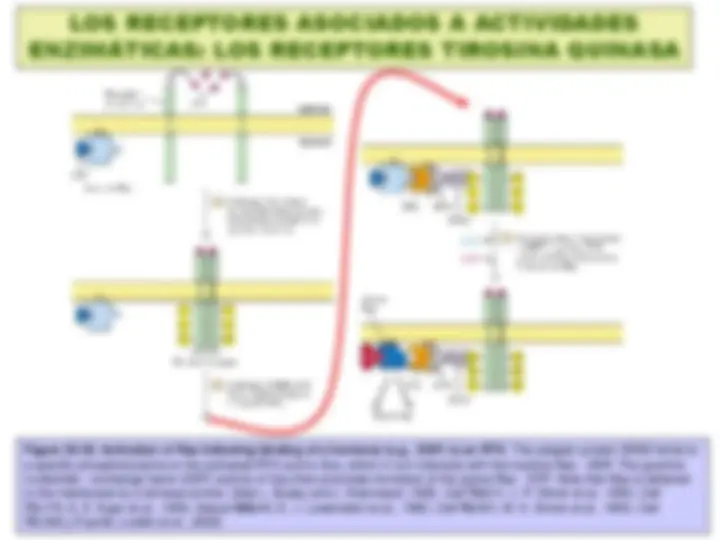
Figure 15-17. A schematic drawing of a G-protein-linked receptor. Receptors that bind protein ligands have a large extracellular ligand-binding domain formed by the part of the polypeptide chain shown in light green. Receptors for small ligands such as adrenaline have small extracellular domains, and the ligand-binding site is usually deep within the plane of the membrane, formed by amino acids from several of the transmembrane segments. The parts of the intracellular domains that are mainly responsible for binding to trimeric G proteins are shown in orange, while those that become phosphorylated during receptor desensitization are shown in red. (Fuente: Alberts et al., 1993)
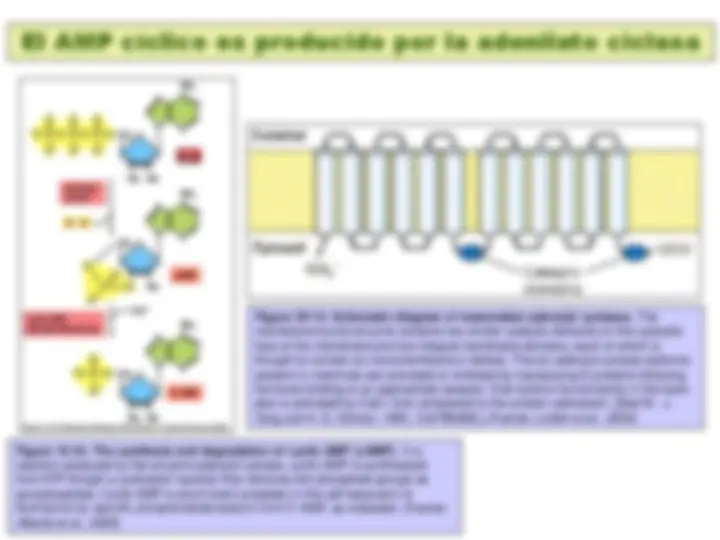
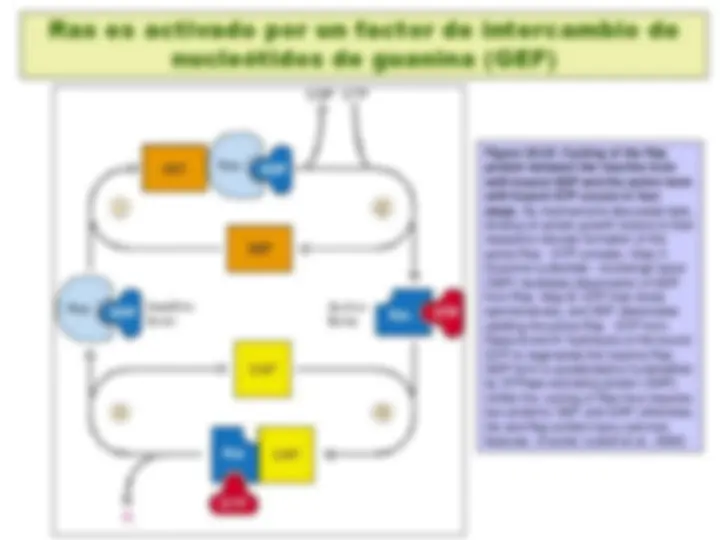
Figure 20-16. Activation of adenylyl cyclase following binding of an appropriate hormone (e.g., epinephrine, glucagon) to a Gs protein – coupled receptor. Following ligand binding to the receptor, the Gs protein relays the hormone signal to the effector protein, in this case adenylyl cyclase. Gs cycles between an inactive form with bound GDP and an active form with bound GTP. Dissociation of the active form yields the Gsα · GTP complex, which directly activates adenylyl cyclase. Activation is short-lived because GTP is rapidly hydrolyzed (step 5 ). This terminates the hormone signal and leads to reassembly of the inactive Gs · GDP form, returning the system to the resting state. Binding of another hormone molecule causes repetition of the cycle. Both the Gγ and Gsα subunits are linked to the membrane by covalent attachment to lipids. Binding of the activated receptor to Gsα promotes dissociation of GDP and its replacement with GTP. (Fuente: Lodish et al., 2000)
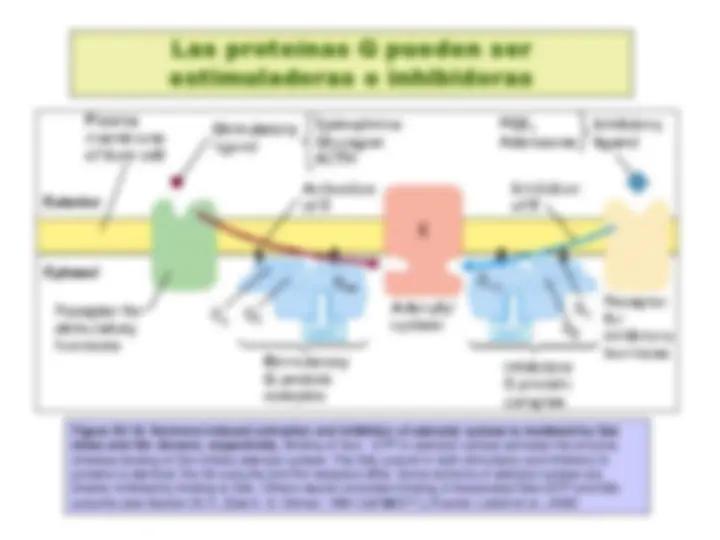
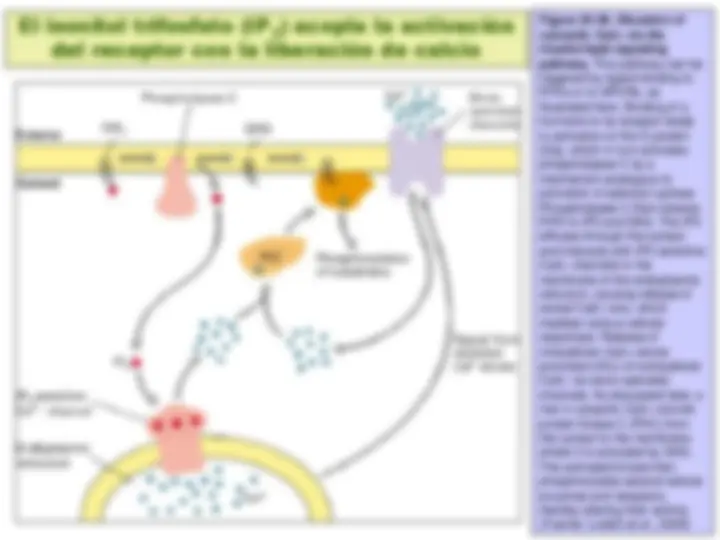
Transducción de la señal por medio de proteínas G triméricas