Baixe Bioquímuica e outras Manuais, Projetos, Pesquisas em PDF para Engenharia Química, somente na Docsity!
Lehninger Principles of Biochemistry Fourth Edition David L. Nelson (U. of Wisconsin–Madison) Michael M. Cox (U. of Wisconsin–Madison)
1. The Foundations of Biochemistry 1.1 Cellular Foundations 1.2 Chemical Foundations 1.3 Physical Foundations 1.4 Genetic Foundations 1.5 Evolutionary Foundations Distilled and reorganized from Chapters 1–3 of the previous edition, this overview provides a refresher on the cellular, chemical, physical, genetic, and evolutionary background to biochemistry, while orienting students toward what is unique about biochemistry.
PART I. STRUCTURE AND CATALYSIS
2. Water 2.1 Weak Interactions in Aqueous Systems 2.2 Ionization of Water, Weak Acids, and Weak Bases 2.3 Buffering against pH Changes in Biological Systems 2.4 Water as a Reactant 2.5 The Fitness of the Aqueous Environment for Living Organisms Includes new coverage of the concept of protein-bound water, illustrated with molecular graphics. 3. Amino Acids, Peptides, and Proteins 3.1 Amino Acids 3.2 Peptides and Proteins 3.3 Working with Proteins 3.4 The Covalent Structure of Proteins 3.5 Protein Sequences and Evolution Adds important new material on genomics and proteomics and their implications for the study of protein structure, function, and evolution. 4. The Three-Dimensional Structure of Proteins 4.1 Overview of Protein Structure 4.2 Protein Secondary Structure 4.3 Protein Tertiary and Quaternary Structures 4.4 Protein Denaturation and Folding Adds a new box on scurvy. 5. Protein Function 5.1 Reversible Binding of a Protein to a Ligand: Oxygen-Binding Proteins 5.2 Complementary Interactions between Proteins and Ligands: The Immune System and Immunoglobulins 5.3 Protein Interactions Modulated by Chemical Energy: Actin, Myosin, and Molecular Motors Adds a new box on carbon monoxide poisoning 6. Enzymes 6.1 An Introduction to Enzymes 6.2 How Enzymes Work 6.3 Enzyme Kinetics as An Approach to Understanding Mechanism 6.4 Examples of Enzymatic Reactions 6.5 Regulatory Enzymes Offers a revised presentation of the mechanism of chymotrypsin (the first reaction mechanism in the book), featuring a two-page figure that takes students through this particular mechanism, while serving as a step-by-step guide to interpreting any
reaction mechanism Features new coverage of the mechanism for lysozyme including the controversial aspects of the mechanism and currently favored resolution based on work published in 2001.
7. Carbohydrates and Glycobiology 7.1 Monosaccharides and Disaccharides 7.2 Polysaccharides 7.3 Glycoconjugates: Proteoglycans, Glycoproteins, and Glycolipids 7.4 Carbohydrates as Informational Molecules: The Sugar Code 7.5 Working with Carbohydrates Includes new section on polysaccharide conformations. A striking new discussion of the "sugar code" looks at polysaccharides as informational molecules, with detailed discussions of lectins, selectins, and oligosaccharide-bearing hormones. Features new material on structural heteropolysaccharides and proteoglycans Covers recent techniques for carbohydrate analysis. 8. Nucleotides and Nucleic Acids 8.1 Some Basics 8.2 Nucleic Acid Structure 8.3 Nucleic Acid Chemistry 8.4 Other Functions of Nucleotides 9. DNA-Based Information Technologies 9.1 DNA Cloning: The Basics 9.2 From Genes to Genomes 9.3 From Genomes to Proteomes 9.4 Genome Alterations and New Products of Biotechnology Introduces the human genome. Biochemical insights derived from the human genome are integrated throughout the text. Tracking the emergence of genomics and proteomics, this chapter establishes DNA technology as a core topic and a path to understanding metabolism, signaling, and other topics covered in the middle chapters of this edition. Includes up-to-date coverage of microarrays, protein chips, comparative genomics, and techniques in cloning and analysis. 10. Lipids 10.1 Storage Lipids 10.2 Structural Lipids in Membranes 10.3 Lipids as Signals, Cofactors, and Pigments 10.4 Working with Lipids Integrates new topics specific to chloroplasts and archaebacteria Adds material on lipids as signal molecules. 11. Biological Membranes and Transport 11.1 The Composition and Architecture of Membranes 11.2 Membrane Dynamics 11.3 Solute Transport across Membranes Includes a description of membrane rafts and microdomains within membranes, and a new box on the use of atomic force microscopy to visualize them. Looks at the role of caveolins in the formation of membrane caveolae Covers the investigation of hop diffusion of membrane lipids using FRAP (fluorescence recovery after photobleaching) Adds new details to the discussion of the mechanism of Ca^2 - ATPase (SERCA
15.1 The Metabolism of Glycogen in Animals 15.2 Regulation of Metabolic Pathways 15.3 Coordinated Regulation of Glycolysis and Gluconeogenesis 15.4 Coordinated Regulation of Glycogen Synthesis and Breakdown 15.5 Analysis of Metabolic Control Brings together the concepts and principles of metabolic regulation in one chapter Concludes with the latest conceptual approaches to the regulation of metabolism, including metabolic control analysis and contemporary methods for studying and predicting the flux through metabolic pathways
16. The Citric Acid Cycle 16.1 Production of Acetyl-CoA (Activated Acetate) 16.2 Reactions of the Citric Acid Cycle 16.3 Regulation of the Citric Acid Cycle 16.4 The Glyoxylate Cycle Expands and updates the presentation of the mechanism for pyruvate carboxylase. Adds coverage of the mechanisms of isocitrate dehydrogenase and citrate synthase. 17. Fatty Acid Catabolism 17.1 Digestion, Mobilization, and Transport of Fats 17.2 Oxidation of Fatty Acids 17.3 Ketone Bodies Updates coverage of trifunctional protein New section on the role of perilipin phosphorylation in the control of fat mobilization New discussion of the role of acetyl-CoA in the integration of fatty acid oxidation and synthesis Updates coverage of the medical consequences of genetic defects in fatty acyl–CoA dehydrogenases Takes a fresh look at medical issues related to peroxisomes 18. Amino Acid Oxidation and the Production of Urea 18.1 Metabolic Fates of Amino Groups 18.2 Nitrogen Excretion and the Urea Cycle 18.3 Pathways of Amino Acid Degradation Integrates the latest on regulation of reactions throughout the chapter, with new material on genetic defects in urea cycle enzymes, and updated information on the regulatory function of N-acetylglutamate synthase. Reorganizes coverage of amino acid degradation to focus on the big picture Adds new material on the relative importance of several degradative pathways Includes a new description of the interplay of the pyridoxal phosphate and tetrahydrofolate cofactors in serine and glycine metabolism 19. Oxidative Phosphorylation and Photophosphorylation Oxidative Phosporylation 19.1 Electron-Transfer Reactions in Mitochondria 19.2 ATP Synthesis 19.3 Regulation of Oxidative Phosphorylation 19.4 Mitochondrial Genes: Their Origin and the Effects of Mutations 19.5 The Role of Mitochondria in Apoptosis and Oxidative Stress Photosynthesis: Harvesting Light Energy 19.6 General Features of Photophosphorylation 19.7 Light Absorption 19.8 The Central Photochemical Event: Light-Driven Electron Flow 19.9 ATP Synthesis by Photophosphorylation Adds a prominent new section on the roles of mitochondria in apoptosis and oxidative stress Now covers the role of IF1 in the inhibition of ATP synthase during ischemia
Includes revelatory details on the light-dependent pathways of electron transfer in photosynthesis, based on newly available molecular structures
20. Carbohydrate Biosynthesis in Plants and Bacteria 20.1 Photosynthetic Carbohydrate Synthesis 20.2 Photorespiration and the C 4 and CAM Pathways 20.3 Biosynthesis of Starch and Sucrose 20.4 Synthesis of Cell Wall Polysaccharides: Plant Cellulose and Bacterial Peptidoglycan 20.5 Integration of Carbohydrate Metabolism in the Plant Cell Reorganizes the coverage of photosynthesis and the C 4 and CAM pathways Adds a major new section on the synthesis of cellulose and bacterial peptidoglycan 21. Lipid Biosynthesis 21.1 Biosynthesis of Fatty Acids and Eicosanoids 21.2 Biosynthesis of Triacylglycerols 21.3 Biosynthesis of Membrane Phospholipids 21.4 Biosynthesis of Cholesterol, Steroids, and Isoprenoids Features an important new section on glyceroneogenesis and the triacylglycerol cycle between adipose tissue and liver, including their roles in fatty acid metabolism (especially during starvation) and the emergence of thiazolidinediones as regulators of glyceroneogenesis in the treatment of type II diabetes Includes a timely new discussion on the regulation of cholesterol metabolism at the genetic level, with consideration of sterol regulatory element-binding proteins (SREBPs). 22. Biosynthesis of Amino Acids, Nucleotides, and Related Molecules 22.1 Overview of Nitrogen Metabolism 22.2 Biosynthesis of Amino Acids 22.3 Molecules Derived from Amino Acids 22.4 Biosynthesis and Degradation of Nucleotides Adds material on the regulation of nitrogen metabolism at the level of transcription Significantly expands coverage of synthesis and degradation of heme 23. Integration and Hormonal Regulation of Mammalian Metabolism 23.1 Tissue-Specific Metabolism: The Division of Labor 23.2 Hormonal Regulation of Fuel Metabolism 23.3 Long Term Regulation of Body Mass 23.4 Hormones: Diverse Structures for Diverse Functions Reorganized presentation leads students through the complex interactions of integrated metabolism step by step Features extensively revised coverage of insulin and glucagon metabolism that includes the integration of carbohydrate and fat metabolism New discussion of the role of AMP-dependent protein kinase in metabolic integration Updates coverage of the fast-moving field of obesity, regulation of body mass, and the leptin and adiponectin regulatory systems Adds a discussion of Ghrelin and PYY3-36 as regulators of short-term eating behavior Covers the effects of diet on the regulation of gene expression, considering the role of peroxisome proliferator-activated receptors (PPARs)
PART III. INFORMATION PATHWAYS
24. Genes and Chromosomes 24.1 Chromosomal Elements 24.2 DNA Supercoiling 24.3 The Structure of Chromosomes
chapter
F
ifteen to twenty billion years ago, the universe arose as a cataclysmic eruption of hot, energy-rich sub- atomic particles. Within seconds, the simplest elements (hydrogen and helium) were formed. As the universe expanded and cooled, material condensed under the in- fluence of gravity to form stars. Some stars became enormous and then exploded as supernovae, releasing the energy needed to fuse simpler atomic nuclei into the more complex elements. Thus were produced, over bil- lions of years, the Earth itself and the chemical elements found on the Earth today. About four billion years ago,
life arose—simple microorganisms with the ability to ex- tract energy from organic compounds or from sunlight, which they used to make a vast array of more complex biomolecules from the simple elements and compounds on the Earth’s surface. Biochemistry asks how the remarkable properties of living organisms arise from the thousands of differ- ent lifeless biomolecules. When these molecules are iso- lated and examined individually, they conform to all the physical and chemical laws that describe the behavior of inanimate matter—as do all the processes occurring in living organisms. The study of biochemistry shows how the collections of inanimate molecules that consti- tute living organisms interact to maintain and perpetu- ate life animated solely by the physical and chemical laws that govern the nonliving universe. Yet organisms possess extraordinary attributes, properties that distinguish them from other collections of matter. What are these distinguishing features of liv- ing organisms? A high degree of chemical complexity and microscopic organization. Thousands of differ- ent molecules make up a cell’s intricate internal structures (Fig. 1–1a). Each has its characteristic sequence of subunits, its unique three-dimensional structure, and its highly specific selection of binding partners in the cell. Systems for extracting, transforming, and using energy from the environment (Fig. 1–1b), enabling organisms to build and maintain their intricate structures and to do mechanical, chemical, osmotic, and electrical work. Inanimate matter tends, rather, to decay toward a more disordered state, to come to equilibrium with its surroundings.
THE FOUNDATIONS
OF BIOCHEMISTRY
1.1 Cellular Foundations 3
1.2 Chemical Foundations 12
1.3 Physical Foundations 21
1.4 Genetic Foundations 28
1.5 Evolutionary Foundations 31
With the cell, biology discovered its atom... To
characterize life, it was henceforth essential to study the
cell and analyze its structure: to single out the common
denominators, necessary for the life of every cell;
alternatively, to identify differences associated with the
performance of special functions.
—François Jacob, La logique du vivant: une histoire de l’hérédité (The Logic of Life: A History of Heredity), 1970
We must, however, acknowledge, as it seems to me, that
man with all his noble qualities... still bears in his
bodily frame the indelible stamp of his lowly origin.
—Charles Darwin, The Descent of Man, 1871
A capacity for precise self-replication and self-assembly (Fig. 1–1c). A single bacterial cell placed in a sterile nutrient medium can give rise to a billion identical “daughter” cells in 24 hours. Each cell contains thousands of different molecules, some extremely complex; yet each bacterium is a faithful copy of the original, its construction directed entirely from information contained within the genetic material of the original cell. Mechanisms for sensing and responding to alterations in their surroundings, constantly adjusting to these changes by adapting their internal chemistry. Defined functions for each of their compo- nents and regulated interactions among them.
This is true not only of macroscopic structures, such as leaves and stems or hearts and lungs, but also of microscopic intracellular structures and indi- vidual chemical compounds. The interplay among the chemical components of a living organism is dy- namic; changes in one component cause coordinat- ing or compensating changes in another, with the whole ensemble displaying a character beyond that of its individual parts. The collection of molecules carries out a program, the end result of which is reproduction of the program and self-perpetuation of that collection of molecules—in short, life. A history of evolutionary change. Organisms change their inherited life strategies to survive in new circumstances. The result of eons of evolution is an enormous diversity of life forms, superficially very different (Fig. 1–2) but fundamentally related through their shared ancestry.
Despite these common properties, and the funda- mental unity of life they reveal, very few generalizations about living organisms are absolutely correct for every organism under every condition; there is enormous di- versity. The range of habitats in which organisms live, from hot springs to Arctic tundra, from animal intestines to college dormitories, is matched by a correspondingly wide range of specific biochemical adaptations, achieved
2 Chapter 1 The Foundations of Biochemistry
(a)
(c)
(b)
FIGURE 1–1 Some characteristics of living matter. (a) Microscopic complexity and organization are apparent in this colorized thin sec- tion of vertebrate muscle tissue, viewed with the electron microscope. (b) A prairie falcon acquires nutrients by consuming a smaller bird. (c) Biological reproduction occurs with near-perfect fidelity.
FIGURE 1–2 Diverse living organisms share common chemical fea- tures. Birds, beasts, plants, and soil microorganisms share with hu- mans the same basic structural units (cells) and the same kinds of macromolecules (DNA, RNA, proteins) made up of the same kinds of monomeric subunits (nucleotides, amino acids). They utilize the same pathways for synthesis of cellular components, share the same genetic code, and derive from the same evolutionary ancestors. Shown here is a detail from “The Garden of Eden,” by Jan van Kessel the Younger (1626–1679).
the complete set of genes, composed of DNA—is stored and replicated. The nucleoid, in bacteria, is not sepa- rated from the cytoplasm by a membrane; the nucleus, in higher organisms, consists of nuclear material en- closed within a double membrane, the nuclear envelope. Cells with nuclear envelopes are called eukaryotes (Greek eu, “true,” and karyon, “nucleus”); those with- out nuclear envelopes—bacterial cells—are prokary- otes (Greek pro, “before”).
Cellular Dimensions Are Limited by Oxygen Diffusion
Most cells are microscopic, invisible to the unaided eye. Animal and plant cells are typically 5 to 100 � m in di- ameter, and many bacteria are only 1 to 2 � m long (see the inside back cover for information on units and their abbreviations). What limits the dimensions of a cell? The lower limit is probably set by the minimum number of each type of biomolecule required by the cell. The smallest cells, certain bacteria known as mycoplasmas, are 300 nm in diameter and have a volume of about 10 �^14 mL. A single bacterial ribosome is about 20 nm in its longest dimension, so a few ribosomes take up a sub- stantial fraction of the volume in a mycoplasmal cell. The upper limit of cell size is probably set by the rate of diffusion of solute molecules in aqueous systems. For example, a bacterial cell that depends upon oxygen- consuming reactions for energy production must obtain
molecular oxygen by diffusion from the surrounding medium through its plasma membrane. The cell is so small, and the ratio of its surface area to its volume is so large, that every part of its cytoplasm is easily reached by O 2 diffusing into the cell. As cell size increases, how- ever, surface-to-volume ratio decreases, until metabo- lism consumes O 2 faster than diffusion can supply it. Metabolism that requires O 2 thus becomes impossible as cell size increases beyond a certain point, placing a theoretical upper limit on the size of the cell.
There Are Three Distinct Domains of Life All living organisms fall into one of three large groups (kingdoms, or domains) that define three branches of evolution from a common progenitor (Fig. 1–4). Two large groups of prokaryotes can be distinguished on bio- chemical grounds: archaebacteria (Greek arche-, “ori- gin”) and eubacteria (again, from Greek eu, “true”). Eubacteria inhabit soils, surface waters, and the tissues of other living or decaying organisms. Most of the well- studied bacteria, including Escherichia coli, are eu- bacteria. The archaebacteria, more recently discovered, are less well characterized biochemically; most inhabit extreme environments—salt lakes, hot springs, highly acidic bogs, and the ocean depths. The available evi- dence suggests that the archaebacteria and eubacteria diverged early in evolution and constitute two separate
4 Chapter 1 The Foundations of Biochemistry
Purple bacteria
Cyanobacteria Flavobacteria
Thermotoga
Extreme halophiles Methanogens (^) Extreme thermophiles
Microsporidia
Flagellates
Plants
Fungi Animals Ciliates
Archaebacteria
Gram- positive bacteria
Eubacteria Eukaryotes
Green nonsulfur bacteria
FIGURE 1–4 Phylogeny of the three domains of life. Phylogenetic relationships are often illustrated by a “family tree” of this type. The fewer the branch points between any two organisms, the closer is their evolutionary relationship.
domains, sometimes called Archaea and Bacteria. All eu- karyotic organisms, which make up the third domain, Eukarya, evolved from the same branch that gave rise to the Archaea; archaebacteria are therefore more closely related to eukaryotes than to eubacteria. Within the domains of Archaea and Bacteria are sub- groups distinguished by the habitats in which they live. In aerobic habitats with a plentiful supply of oxygen, some resident organisms derive energy from the trans- fer of electrons from fuel molecules to oxygen. Other environments are anaerobic, virtually devoid of oxy- gen, and microorganisms adapted to these environments obtain energy by transferring electrons to nitrate (form- ing N 2 ), sulfate (forming H 2 S), or CO 2 (forming CH 4 ). Many organisms that have evolved in anaerobic envi- ronments are obligate anaerobes: they die when ex- posed to oxygen. We can classify organisms according to how they obtain the energy and carbon they need for synthesiz- ing cellular material (as summarized in Fig. 1–5). There are two broad categories based on energy sources: pho- totrophs (Greek trophe-, “nourishment”) trap and use sunlight, and chemotrophs derive their energy from oxidation of a fuel. All chemotrophs require a source of organic nutrients; they cannot fix CO 2 into organic com- pounds. The phototrophs can be further divided into those that can obtain all needed carbon from CO 2 (au- totrophs) and those that require organic nutrients (heterotrophs). No chemotroph can get its carbon
atoms exclusively from CO 2 (that is, no chemotrophs are autotrophs), but the chemotrophs may be further classified according to a different criterion: whether the fuels they oxidize are inorganic (lithotrophs) or or- ganic (organotrophs). Most known organisms fall within one of these four broad categories—autotrophs or heterotrophs among the photosynthesizers, lithotrophs or organotrophs among the chemical oxidizers. The prokaryotes have several gen- eral modes of obtaining carbon and energy. Escherichia coli, for example, is a chemoorganoheterotroph; it re- quires organic compounds from its environment as fuel and as a source of carbon. Cyanobacteria are photo- lithoautotrophs; they use sunlight as an energy source and convert CO 2 into biomolecules. We humans, like E. coli , are chemoorganoheterotrophs.
Escherichia coli Is the Most-Studied Prokaryotic Cell Bacterial cells share certain common structural fea- tures, but also show group-specific specializations (Fig. 1–6). E. coli is a usually harmless inhabitant of the hu- man intestinal tract. The E. coli cell is about 2 � m long and a little less than 1 � m in diameter. It has a protec- tive outer membrane and an inner plasma membrane that encloses the cytoplasm and the nucleoid. Between the inner and outer membranes is a thin but strong layer of polymers called peptidoglycans, which gives the cell its shape and rigidity. The plasma membrane and the
1.1 Cellular Foundations 5
Heterotrophs (carbon from organic compounds) Examples: •Purple bacteria •Green bacteria
Autotrophs (carbon from CO 2 ) Examples: •Cyanobacteria •Plants
Heterotrophs (carbon from organic compounds)
Phototrophs (energy from light)
Chemotrophs (energy from chemical compounds)
All organisms
Lithotrophs (energy from inorganic compounds) Examples: •Sulfur bacteria •Hydrogen bacteria
Organotrophs (energy from organic compounds) Examples: •Most prokaryotes •All nonphototrophic eukaryotes
FIGURE 1–5 Organisms can be classified according to their source of energy (sunlight or oxidizable chemical compounds) and their source of carbon for the synthesis of cellular material.
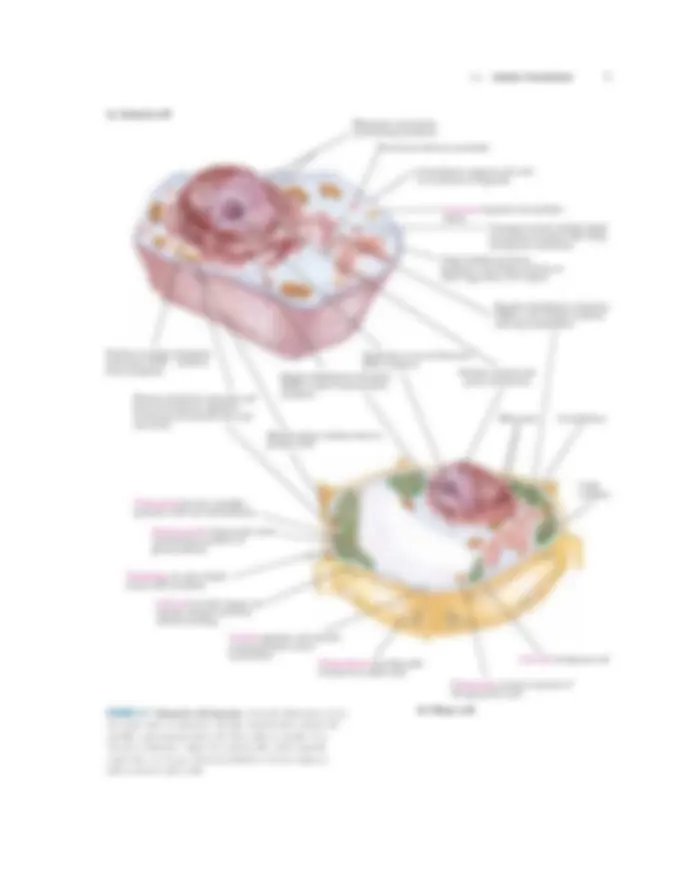
1.1 Cellular Foundations 7
Ribosomes are protein- synthesizing machines Peroxisome destroys peroxides
Lysosome degrades intracellular debris Transport vesicle shuttles lipids and proteins between ER, Golgi, and plasma membrane Golgi complex processes, packages, and targets proteins to other organelles or for export
Smooth endoplasmic reticulum (SER) is site of lipid synthesis and drug metabolism
Nucleus contains the genes (chromatin)
Ribosomes Cytoskeleton
Cytoskeleton supports cell, aids in movement of organells
Golgi complex
Nucleolus is site of ribosomal RNA synthesis Rough endoplasmic reticulum (RER) is site of much protein synthesis
Mitochondrion oxidizes fuels to produce ATP
Plasma membrane separates cell from environment, regulates movement of materials into and out of cell
Chloroplast harvests sunlight, produces ATP and carbohydrates
Starch granule temporarily stores carbohydrate products of photosynthesis
Thylakoids are site of light- driven ATP synthesis Cell wall provides shape and rigidity; protects cell from osmotic swelling
Plasmodesma provides path Cell wall of adjacent cell between two plant cells
Nuclear envelope segregates chromatin (DNA � protein) from cytoplasm
Vacuole degrades and recycles macromolecules, stores metabolites
(a) Animal cell
(b) Plant cell
Glyoxysome contains enzymes of the glyoxylate cycle
FIGURE 1–7 Eukaryotic cell structure. Schematic illustrations of the two major types of eukaryotic cell: (a) a representative animal cell and (b) a representative plant cell. Plant cells are usually 10 to 100 � m in diameter—larger than animal cells, which typically range from 5 to 30 � m. Structures labeled in red are unique to either animal or plant cells.
❚
❚ ❚
❚
❚
❚
❚
❚^
❚
❚
❚
❚❚
❚
❚ ❚ ❚
❚ ❚
❚
❚ ❚
❚
❚
❚
❚
❚ ❚ ❚
❚ ❚
❚
❚ ❚ ❚
❚
❚
❚
❚
❚
❚
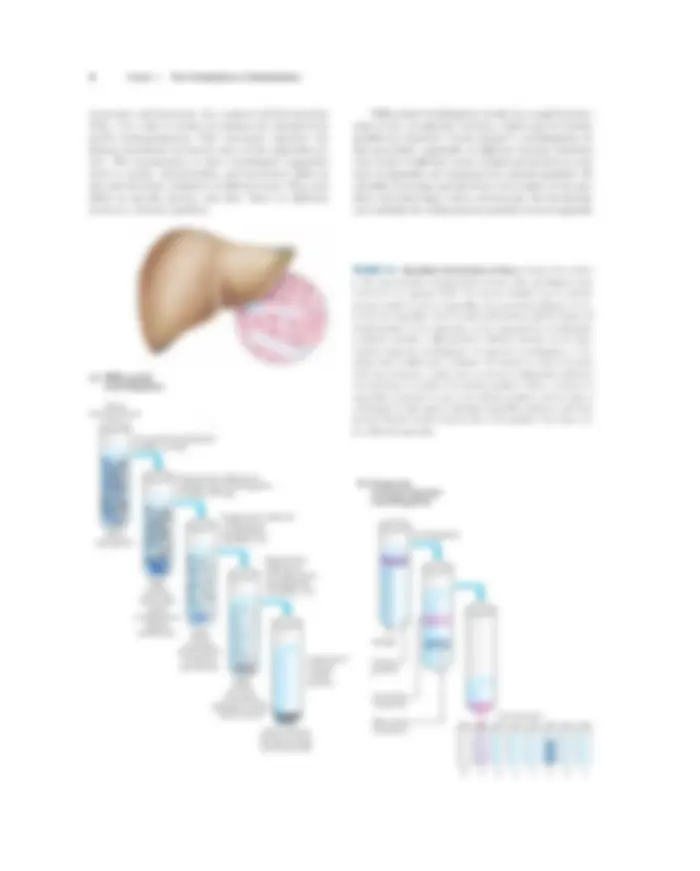
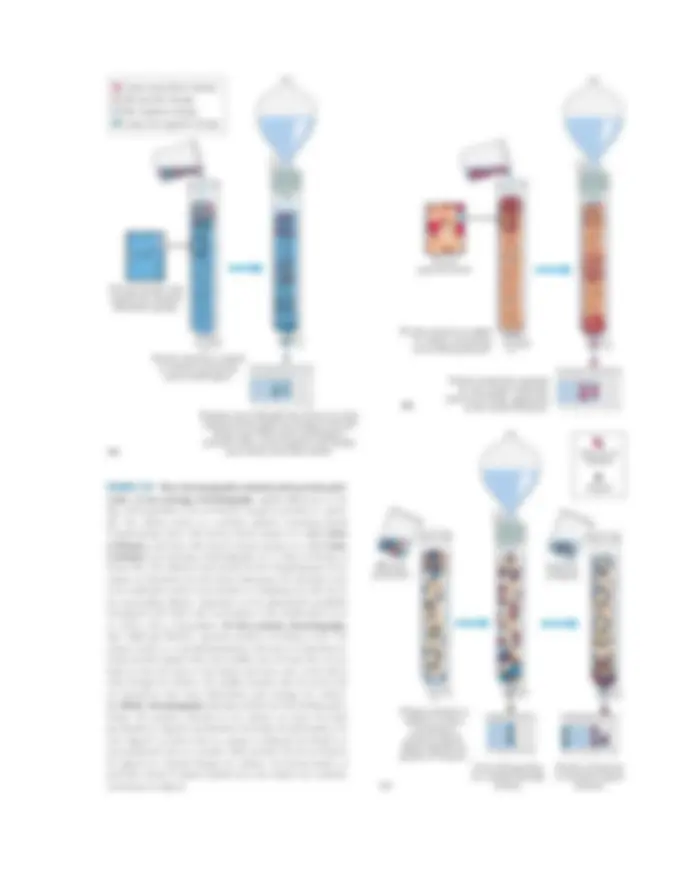
structures and functions. In a typical cell fractionation (Fig. 1–8), cells or tissues in solution are disrupted by gentle homogenization. This treatment ruptures the plasma membrane but leaves most of the organelles in- tact. The homogenate is then centrifuged; organelles such as nuclei, mitochondria, and lysosomes differ in size and therefore sediment at different rates. They also differ in specific gravity, and they “float” at different levels in a density gradient.
Differential centrifugation results in a rough fraction- ation of the cytoplasmic contents, which may be further purified by isopycnic (“same density”) centrifugation. In this procedure, organelles of different buoyant densities (the result of different ratios of lipid and protein in each type of organelle) are separated on a density gradient. By carefully removing material from each region of the gra- dient and observing it with a microscope, the biochemist can establish the sedimentation position of each organelle
8 Chapter 1 The Foundations of Biochemistry
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚^
❚
❚
❚
❚
Centrifugation
Fractionation
Sample
Less dense component More dense component
Sucrose gradient
8 7 6 5 4 3 2 1
❚ ❚ ❚
❚ ❚ ❚
❚ ❚
❚
❚ ❚
❚
❚
❚
❚
❚ ❚ ❚
❚ ❚
❚
❚ ❚ ❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚
❚^
❚
❚
❚
❚
Isopycnic (sucrose-density) centrifugation
❚ ❚ ❚
❚ ❚ ❚
❚ ❚
❚
❚ ❚
❚
❚
❚
❚
❚ ❚ ❚
❚
(b)
❚
❚
❚ ❚ ❚
❚
❚
❚
❚
❚
❚
❚❚
▲
▲
▲
▲ ▲ ▲ ▲
▲ ▲
▲▲
▲
▲
▲▲
▲
▲ ▲
▲ ▲ ❚❚
❚❚
❚❚
❚❚
❚❚
❚❚
❚❚
❚❚
❚❚^
❚❚
❚❚
❚
❚❚
▲
Low-speed centrifugation (1,000 g , 10 min)
Supernatant subjected to medium-speed centrifugation (20,000 g , 20 min)
Supernatant subjected to high-speed centrifugation (80,000 g , 1 h)
Supernatant subjected to very high-speed centrifugation (150,000 g , 3 h)
Differential centrifugation
Tissue homogenization
Tissue homogenate
Pellet contains mitochondria, lysosomes, peroxisomes Pellet contains microsomes (fragments of ER), small vesicles Pellet contains ribosomes, large macromolecules
Pellet contains whole cells, nuclei, cytoskeletons, plasma membranes
Supernatant contains soluble proteins
❚❚❚❚
❚❚
❚❚ ❚❚ ❚❚
❚❚ ❚❚
❚❚
❚❚ ❚❚
❚❚
❚❚
❚❚
❚❚
❚❚
▲
❚❚ ❚❚
❚❚
(a)
▲ ▲▲▲ ▲
▲▲▲
▲
▲ ▲
▲▲
▲ (^) ▲
▲
▲
❚❚
❚❚
❚❚ ❚❚ ❚❚
❚❚
❚❚
❚❚
❚❚
❚❚
❚❚
▲▲ ▲
▲▲▲▲▲▲
❚❚❚❚❚❚❚❚❚❚ ❚❚❚❚❚❚❚❚❚❚❚❚
FIGURE 1–8 Subcellular fractionation of tissue. A tissue such as liver is first mechanically homogenized to break cells and disperse their contents in an aqueous buffer. The sucrose medium has an osmotic pressure similar to that in organelles, thus preventing diffusion of wa- ter into the organelles, which would swell and burst. (a) The large and small particles in the suspension can be separated by centrifugation at different speeds, or (b) particles of different density can be sepa- rated by isopycnic centrifugation. In isopycnic centrifugation, a cen- trifuge tube is filled with a solution, the density of which increases from top to bottom; a solute such as sucrose is dissolved at different concentrations to produce the density gradient. When a mixture of organelles is layered on top of the density gradient and the tube is centrifuged at high speed, individual organelles sediment until their buoyant density exactly matches that in the gradient. Each layer can be collected separately.
Although complex, this organization of the cyto- plasm is far from random. The motion and the position- ing of organelles and cytoskeletal elements are under tight regulation, and at certain stages in a eukaryotic cell’s life, dramatic, finely orchestrated reorganizations, such as the events of mitosis, occur. The interactions be- tween the cytoskeleton and organelles are noncovalent,
reversible, and subject to regulation in response to var- ious intracellular and extracellular signals.
Cells Build Supramolecular Structures Macromolecules and their monomeric subunits differ greatly in size (Fig. 1–10). A molecule of alanine is less than 0.5 nm long. Hemoglobin, the oxygen-carrying pro- tein of erythrocytes (red blood cells), consists of nearly 600 amino acid subunits in four long chains, folded into globular shapes and associated in a structure 5.5 nm in diameter. In turn, proteins are much smaller than ribo- somes (about 20 nm in diameter), which are in turn much smaller than organelles such as mitochondria, typ- ically 1,000 nm in diameter. It is a long jump from sim- ple biomolecules to cellular structures that can be seen
10 Chapter 1 The Foundations of Biochemistry
Uracil Thymine
- D -Ribose (^) 2-Deoxy- -D-ribose
O
H OH
NH (^2)
HOCH (^2)
Cytosine
H H H OH H
O
H OH
HOCH (^2) H H H OH OH
Adenine Guanine
COO�
Oleate
Palmitate H
CH 2 OH O
HO
OH
H H H OH OH
H
(b) The components of nucleic acids (c) Some components of lipids
(d) The parent sugar
HO P
O�
O
OH
Phosphoric acid
N
Choline
� CH 2 CH 2 OH CH (^3)
CH 3 CH 3
Glycerol
CH 2 OH CHOH CH 2 OH
CH (^2)
CH 3
CH 2
CH 2 CH 2 CH 2 CH 2
CH 2 CH 2 CH 2 CH 2 CH 2 CH 3
CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
COO� CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
CH 2
CH CH
� �
�
C
NH 2
C C
CH HC
N
N N H
N C
O
C C
CH C
HN
N N H
N
C
O
O
CH
CH C
HN
N H
O
CH
CH C
N
N H
C
O
O
CH
C C
HN
N H
H 2 N
CH (^3)
Nitrogenous bases
Five-carbon sugars
H (^3)
� N
H (^3)
� N H^3
�
�
N H (^3)
� N
OC
A COO�
COO�^ COO� COO�
H (^3)
� N
COO� H (^3)
� N
COO�
COO�
A CH (^3)
OH OC
A A CH 2 OH
OH (^) OC
A A C A H (^2)
OH
Alanine Serine Aspartate
OC
A A C A SH
H (^2)
OH
Cysteine Histidine
C
A OC
A OH H 2
OH Tyrosine
OC
A A C A H 2
OH
C H CH HC
N
NH
(a) Some of the amino acids of proteins
FIGURE 1–10 The organic compounds from which most cellular materials are constructed: the ABCs of biochemistry. Shown here are (a) six of the 20 amino acids from which all proteins are built (the side chains are shaded pink); (b) the five nitrogenous bases, two five- carbon sugars, and phosphoric acid from which all nucleic acids are built; (c) five components of membrane lipids; and (d) D-glucose, the parent sugar from which most carbohydrates are derived. Note that phosphoric acid is a component of both nucleic acids and membrane lipids.
with the light microscope. Figure 1–11 illustrates the structural hierarchy in cellular organization. The monomeric subunits in proteins, nucleic acids, and polysaccharides are joined by covalent bonds. In supramolecular complexes, however, macromolecules are held together by noncovalent interactions—much weaker, individually, than covalent bonds. Among these noncovalent interactions are hydrogen bonds (between polar groups), ionic interactions (between charged groups), hydrophobic interactions (among nonpolar groups in aqueous solution), and van der Waals inter- actions—all of which have energies substantially smaller than those of covalent bonds (Table 1–1). The nature of these noncovalent interactions is described in Chap- ter 2. The large numbers of weak interactions between macromolecules in supramolecular complexes stabilize these assemblies, producing their unique structures.
In Vitro Studies May Overlook Important Interactions
among Molecules
One approach to understanding a biological process is to study purified molecules in vitro (“in glass”—in the test tube), without interference from other molecules present in the intact cell—that is, in vivo (“in the liv- ing”). Although this approach has been remarkably re- vealing, we must keep in mind that the inside of a cell is quite different from the inside of a test tube. The “in- terfering” components eliminated by purification may be critical to the biological function or regulation of the molecule purified. For example, in vitro studies of pure
1.1 Cellular Foundations 11
Level 4: The cell and its organelles
Level 3: Supramolecular complexes
Level 2: Macromolecules
Level 1: Monomeric units
Nucleotides
Amino acids
Protein
Cellulose
Plasma membrane
Chromosome
Cell wall
Sugars
DNA (^) O� �O PO CH (^2) OO
NH 2
H H
N
N
H OHH
O H
H H 3 N CCOO� CH 3
H
O^ H OH
CH^2 OH H HO OH
OH H
O
CH 2 OH H
�
FIGURE 1–11 Structural hierarchy in the molecular organization of cells. In this plant cell, the nucleus is an organelle containing several types of supramolecular complexes, including chromosomes. Chro-
mosomes consist of macromolecules of DNA and many different pro- teins. Each type of macromolecule is made up of simple subunits— DNA of nucleotides (deoxyribonucleotides), for example.
*The greater the energy required for bond dissociation (breakage), the stronger the bond.
TABLE 1–1 Strengths of Bonds Common
in Biomolecules
Bond Bond dissociation dissociation Type energy* Type energy of bond (kJ/mol) of bond (kJ/mol)
Single bonds Double bonds OOH 470 CPO 712 HOH 435 CPN 615 POO 419 CPC 611 COH 414 PPO 502 NOH 389 COO 352 Triple bonds COC 348 CmC 816 SOH 339 NmN 930 CON 293 COS 260 NOO 222 SOS 214
enzymes are commonly done at very low enzyme con- centrations in thoroughly stirred aqueous solutions. In the cell, an enzyme is dissolved or suspended in a gel- like cytosol with thousands of other proteins, some of which bind to that enzyme and influence its activity.
form the strongest bonds. The trace elements (Fig. 1–12) represent a miniscule fraction of the weight of the hu- man body, but all are essential to life, usually because they are essential to the function of specific proteins, including enzymes. The oxygen-transporting capacity of the hemoglobin molecule, for example, is absolutely dependent on four iron ions that make up only 0.3% of its mass.
Biomolecules Are Compounds of Carbon with
a Variety of Functional Groups
The chemistry of living organisms is organized around carbon, which accounts for more than half the dry weight of cells. Carbon can form single bonds with hy- drogen atoms, and both single and double bonds with oxygen and nitrogen atoms (Fig. 1–13). Of greatest sig- nificance in biology is the ability of carbon atoms to form very stable carbon–carbon single bonds. Each carbon atom can form single bonds with up to four other car- bon atoms. Two carbon atoms also can share two (or three) electron pairs, thus forming double (or triple) bonds. The four single bonds that can be formed by a car- bon atom are arranged tetrahedrally, with an angle of about 109.5� between any two bonds (Fig. 1–14) and an average length of 0.154 nm. There is free rotation around each single bond, unless very large or highly charged groups are attached to both carbon atoms, in which case rotation may be restricted. A double bond is shorter (about 0.134 nm) and rigid and allows little rotation about its axis. Covalently linked carbon atoms in biomolecules can form linear chains, branched chains, and cyclic struc- tures. To these carbon skeletons are added groups of other atoms, called functional groups, which confer specific chemical properties on the molecule. It seems likely that the bonding versatility of carbon was a ma- jor factor in the selection of carbon compounds for the molecular machinery of cells during the origin and evo- lution of living organisms. No other chemical element can form molecules of such widely different sizes and shapes or with such a variety of functional groups. Most biomolecules can be regarded as derivatives of hydrocarbons, with hydrogen atoms replaced by a va- riety of functional groups to yield different families of organic compounds. Typical of these are alcohols, which have one or more hydroxyl groups; amines, with amino groups; aldehydes and ketones, with carbonyl groups; and carboxylic acids, with carboxyl groups (Fig. 1–15). Many biomolecules are polyfunctional, containing two or more different kinds of functional groups (Fig. 1–16), each with its own chemical characteristics and reac- tions. The chemical “personality” of a compound is de- termined by the chemistry of its functional groups and their disposition in three-dimensional space.
1.2 Chemical Foundations 13
H C HH H
� O
� C O
� C N
C
C
O
� N
� C
� C C C
� C C C C
C
C
C
C
C
C
C
C �
OO
C
C
C
N
C
N
OO
C
C C
C N
N
C O
C
C
C
FIGURE 1–13 Versatility of carbon bonding. Carbon can form cova- lent single, double, and triple bonds (in red), particularly with other carbon atoms. Triple bonds are rare in biomolecules.
FIGURE 1–14 Geometry of carbon bonding. (a) Carbon atoms have a characteristic tetrahedral arrangement of their four single bonds. (b) Carbon–carbon single bonds have freedom of rotation, as shown for the compound ethane (CH 3 OCH 3 ). (c) Double bonds are shorter and do not allow free rotation. The two doubly bonded carbons and the atoms designated A, B, X, and Y all lie in the same rigid plane.
(a) (b)
(c)
109.5°
109.5°
C C
C
120 °
X
C C
A
B
Y
Cells Contain a Universal Set of Small Molecules
Dissolved in the aqueous phase (cytosol) of all cells is a collection of 100 to 200 different small organic mole- cules ( M r ~100 to ~500), the central metabolites in the major pathways occurring in nearly every cell—the metabolites and pathways that have been conserved throughout the course of evolution. (See Box 1–1 for an explanation of the various ways of referring to molecu-
lar weight.) This collection of molecules includes the common amino acids, nucleotides, sugars and their phosphorylated derivatives, and a number of mono-, di-, and tricarboxylic acids. The molecules are polar or charged, water soluble, and present in micromolar to millimolar concentrations. They are trapped within the cell because the plasma membrane is impermeable to them—although specific membrane transporters can catalyze the movement of some molecules into and out
14 Chapter 1 The Foundations of Biochemistry
Hydroxyl R O H (alcohol)
Carbonyl (aldehyde)
R C O
H
Carbonyl (ketone)
R C O
(^1) R 2
Carboxyl R C O
O�
O�^ O�
O�
Methyl R C
H
H
H
Ethyl R C
H
H
C
H
H
H
Ester R 1 C O
O R^2
Ether R 1 O R 2
Sulfhydryl R S H
Disulfide R 1 S S R^2
Phosphoryl R O P O
OH
Thioester R 1 C O
S R^2
Anhydride R 1 C O O
C R^2 (two car- boxylic acids)
O
Imidazole R N
C CH HN
H
C
Guanidino R N
H C N H
N
H
H
Amino R N
H
H
Amido R C O
N
H
H
Phenyl R C CH C H
H C
C
C
H
H
(carboxylic acid and phosphoric acid; also called acyl phosphate)
Mixed anhydride R C O O
OH
Phosphoanhydride R 1 O
O P R 2 O
P
O�
O
P
FIGURE 1–15 Some common functional O O R groups of biomolecules. In this figure and throughout the book, we use R to represent “any substituent.” It may be as simple as a hydrogen atom, but typically it is a carbon-containing moiety. When two or more substituents are shown in a molecule, we designate them R^1 , R^2 , and so forth.