Baixe Genes capitulo 1 e outras Manuais, Projetos, Pesquisas em PDF para Cultura, somente na Docsity!
dupla hélice e pareiam umas com as outras por meio de
pontes de hidrogênio, formando somente os pares A-T ou
G-C.
- O diâmetro da dupla hélice é de 20 Å, havendo uma volta
completa a cada 34 Å, com cada volta composta por dez
pares de bases.
- A dupla hélice forma um sulco maior (largo) e outro me-
nor (estreito).
1.7 A Replicação do DNA É Semiconservativa
- O experimento de Meselson-Stahl empregou marcação por
densidade para provar que uma única cadeia polinucleotí-
dica é a unidade de DNA que é conservada durante a repli-
cação.
- Cada fita de um duplex de DNA atua como molde na sínte-
se de uma fita filha.
- As seqüências das fitas filhas são determinadas pelo pa-
reamento de bases complementares com as fitas paren-
tais separadas.
1.8 As Fitas de DNA São Separadas na Forquilha de Re- plicação
- A replicação do DNA é realizada por um complexo enzimá-
tico que separa as fitas parentais e sintetiza as fitas fi-
lhas.
- A forquilha de replicação é o ponto onde as fitas paren-
tais são separadas.
- As enzimas que sintetizam DNA são denominadas DNA po-
limerases; as enzimas que sintetizam RNA são denomina-
das RNA polimerases.
- Nucleases são enzimas que degradam ácidos nucléicos;
estas incluem DNAases e RNAases e podem ser divididas
em endonucleases e exonucleases.
1.9 A Informação Genética Pode Ser Fornecida pelo DNA ou RNA
- Os genes celulares são de DNA, embora vírus e viróides
possam apresentar genomas de RNA.
- O DNA é convertido em RNA pela transcrição, e o RNA
pode ser convertido em DNA pela transcrição reversa.
- A tradução do RNA em proteína é unidirecional.
1.10 Os Ácidos Nucléicos Hibridizam-se por Pareamento de Bases
- O aquecimento promove a separação das duas fitas de um
DNA duplex.
- A T m é o ponto médio da faixa de temperatura de desnatu-
ração.
- Fitas simples complementares podem renaturar quando a
temperatura é reduzida.
1.1 Introdução
1.2 O DNA É o Material Genético das Bactérias
- A transformação bacteriana forneceu a primeira prova de
que o DNA é o material genético das bactérias. As pro-
priedades genéticas podem ser transferidas de uma linha-
gem bacteriana para outra, extraindo-se o DNA da primei-
ra linhagem e adicionando-o à segunda.
1.3 O DNA É o Material Genético dos Vírus
- A infecção por fagos provou que o DNA é o material gené-
tico dos vírus. Quando o DNA e as proteínas de bacterió-
fagos são marcados com diferentes isótopos radioativos,
apenas o DNA é transmitido para a progênie de fagos pro-
duzida pelas bactérias infectadas.
1.4 O DNA É o Material Genético das Células Animais
- O DNA pode ser utilizado para introduzir novas carac-
terísticas genéticas em células animais ou em animais
inteiros.
- Em alguns vírus, o material genético é RNA.
1.5 As Cadeias Polinucleotídicas Possuem Bases Nitro- genadas Ligadas a um Esqueleto de Açúcar-Fosfato
- Um nucleosídeo consiste em uma base de purina ou piri-
midina ligada à posição 1 de um açúcar do tipo pentose.
- As posições no anel de ribose são designadas por um após-
trofo (’) para distingui-las.
- A diferença entre o DNA e o RNA está no grupamento da
posição 2’ do açúcar. O DNA possui um açúcar do tipo
desoxirribose (2’-H); o RNA tem um açúcar do tipo ribose
(2’-OH).
- Um nucleotídeo consiste em um nucleosídeo ligado ao
grupamento fosfato tanto na posição 5’ ou 3’ da
(desoxi)ribose.
- Resíduos sucessivos de (desoxi)ribose de uma cadeia po-
linucleotídica são ligados por um grupamento fosfato en-
tre a posição 3’ de um açúcar e a posição 5’ do açúcar
adjacente.
- Uma das extremidades da cadeia (convencionalmente, a
esquerda) possui uma extremidade 5’ livre, e a outra, uma
extremidade 3’ livre.
- O DNA contém as quatro bases: adenina, guanina, citosi-
na e timina; o RNA possui uracila, ao invés de timina.
1.6 O DNA É uma Dupla Hélice
- A forma B do DNA é uma dupla hélice consistindo em duas
cadeias polinucleotídicas de sentido antiparalelo.
- As bases nitrogenadas de cada cadeia são anéis planos de
purinas ou pirimidinas que se projetam para o interior da
Continua na próxima página
Os Genes São DNA
2 CAPÍTULO 1 OS GENES SÃO DNA
- A desnaturação e a renaturação/hibridização po-
dem ocorrer entre combinações de DNA-DNA,
DNA-RNA ou RNA-RNA e podem ser intermole-
culares ou intramoleculares.
- A capacidade de duas preparações de ácidos nu-
cléicos de fita simples hibridizarem é uma me-
dida de suas complementaridades.
1.11 As Mutações Modificam a Seqüência do DNA
- Todas as mutações consistem em modificações
na seqüência do DNA.
- Mutações podem ocorrer espontaneamente ou
serem induzidas por agentes mutagênicos.
1.12 As Mutações Podem Afetar Pares de Bases Únicos ou Seqüências mais Longas
- Uma mutação pontual altera um único par de
bases.
- Mutações pontuais podem ser causadas pela con-
versão química de uma base em outra, ou por
erros que ocorrem durante a replicação.
- Uma transição substitui um par de bases G-C
por um par de bases A-T, ou vice-versa.
- Uma transversão substitui uma purina por uma
pirimidina, tal como a modificação A-T em T-A.
- Inserções são o tipo mais comum de mutação e
resultam da movimentação de elementos de
transposição.
1.13 Os Efeitos das Mutações Podem Ser Revertidos
nucléicos que compreendem o genoma pode con- ter um grande número de genes. Genomas de seres vivos podem conter de menos de 500 genes (no caso de um micoplasma, um tipo de bactéria) até mais de 25.000, em um ser humano. Neste capítulo, analisaremos as propriedades dos genes no que se refere à sua construção molecular básica. A FIGURA 1.1 resume os estágios de transi- ção a partir do conceito histórico do gene até a de- finição moderna do genoma. Um genoma consiste no conjunto completo de cromossomos de qualquer organismo em particu- lar. Este compreende, portanto, uma série de mo- léculas de DNA (uma para cada cromossomo), cada uma contendo muitos genes. A definição final de um genoma está na determinação da seqüência de DNA de cada cromossomo. A primeira definição de gene como uma unidade funcional se seguiu à descoberta de que genes indivi- duais eram responsáveis pela produção de proteínas específicas. A diferença na natureza química entre o DNA do gene e o seu produto protéico levou ao con- ceito de que um gene codifica uma proteína. Isso, por sua vez, levou à descoberta de um aparato complexo que permite à seqüência de DNA de um gene origi- nar a seqüência de aminoácidos de uma proteína.
1.1 Introdução
A natureza hereditária de todo o organismo vivo é definida por seu genoma , que consiste em uma lon- ga seqüência de ácido nucléico, que fornece a in- formação necessária para construir o organismo. Uti- lizamos o termo “informação”, porque o genoma por si só não desempenha qualquer papel ativo na construção do organismo: na realidade, é a seqüên- cia das subunidades individuais (bases) do ácido nucléico que determina as características hereditá- rias. Por meio de uma complexa série de intera- ções, essa seqüência é utilizada para produzir todas as proteínas do organismo no momento e local apro- priados. As proteínas fazem parte da estrutura do organismo, ou apresentam a capacidade de cons- truir as estruturas, ou ainda realizam as reações me- tabólicas necessárias à vida. O genoma contém o conjunto completo da in- formação hereditária para qualquer organismo. Fi- sicamente, o genoma pode ser dividido em algu- mas moléculas diferentes de ácidos nucléicos. Fun- cionalmente este pode ser dividido em genes. Cada gene corresponde a uma seqüência no interior do ácido nucléico, a qual representa uma única proteí- na. Cada uma das diferentes moléculas de ácidos
- As mutações diretas inativam um gene, e as mu-
tações reversas (ou revertentes) revertem seus
efeitos.
- As inserções podem ser revertidas pela deleção
do material inserido, e as deleções não são re-
vertidas.
- A supressão ocorre quando uma mutação em um
segundo gene anula o efeito da mutação do pri-
meiro gene.
1.14 As Mutações São Concentradas em Hotspots
- A freqüência de uma mutação em qualquer par
de bases é determinada por uma flutuação esta-
tística, exceto nos hotspots (“sítios quentes”),
onde a freqüência é aumentada em pelo menos
uma ordem de grandeza.
1.15 Muitos Hotspots Resultam de Bases Modificadas
- Uma causa comum de ocorrência de hotspots é a
presença da base modificada 5-metilcitosina, que
é espontaneamente desaminada, originando ti-
mina.
1.16 Alguns Agentes Hereditários São extremamente Pequenos
- Alguns agentes hereditários muito pequenos não
codificam proteínas e consistem em RNA ou pro-
teína com propriedades hereditárias.
1.17 Resumo
4 CAPÍTULO 1 OS GENES SÃO DNA
Neste experimento, as bactérias S mortas eram do tipo III. As bactérias R vivas eras derivadas do tipo II. As bactérias virulentas recuperadas da in- fecção mista apresentavam um envoltório liso do tipo III. Assim, alguma propriedade das bactérias S do tipo III mortas podia transformar as bactérias R vivas, de tal forma que eram capazes de sintetizar o polissacarídeo capsular do tipo III, tornando-se vi- rulentas. A FIGURA 1.4 ilustra a identificação do compo- nente das bactérias mortas que é responsável pela transformação. Este foi denominado princípio transformante. Ele foi purificado por meio do de- senvolvimento de um sistema livre de células, no qual extratos das bactérias S mortas podiam ser adi- cionados às bactérias R vivas antes da injeção no
FIGURA 1.3 Bactérias do tipo S mortas pelo calor, ou bactérias vivas do tipo R não matam os camundongos, mas a injeção simultânea de ambas pode matar os camundongos tão eficien- temente quanto células do tipo S vivas.
Com cápsula Aparência lisa (S)
Sem cápsula Aparência rugosa (R)
Transformação de bactérias
Tipos de pneumococos
S vivas
Injeção de células Resultado
S mortas pelo calor
R vivas
S mortas pelo calor R vivas
Vive
Vive
Morre
Morre
animal. A purificação do princípio transformante, em 1944, revelou que se tratava do ácido desoxir- ribonucléico (DNA).
1.3 O DNA É o Material Genético dos Vírus
Conceitos Essenciais
- A infecção por fagos provou que o DNA é o material genético dos vírus. Quando o DNA e as proteínas de bacteriófagos são marcados com diferentes isótopos ra- dioativos, apenas o DNA é transmitido para a progênie de fagos produzida pelas bactérias infectadas.
A partir da demonstração de que o DNA era o ma- terial genético das bactérias, o próximo passo foi demonstrar que o DNA era o material genético de um sistema bastante diferente. O fago T2 é um ví- rus que infecta a bactéria Escherichia coli. Quando partículas do fago (vírus) são adicionadas às bacté- rias, estas as adsorvem em sua superfície externa, parte do material penetra na bactéria e, após cer- ca de 20 minutos, cada bactéria se rompe (sofre lise), liberando um grande número da progênie de fagos. A FIGURA 1.5 ilustra os resultados de um expe- rimento realizado em 1952, no qual bactérias fo- ram infectadas com fagos T2, os quais haviam sido marcados radioativamente ou em seu DNA (com (^32) P), ou em seu componente protéico (com 35 S).
As bactérias infectadas foram agitadas em um ho- mogeneizador, e duas frações foram separadas por centrifugação. Uma fração continha os capsídeos vazios dos fagos que foram liberados da superfície das bactérias; a outra consistia nas próprias bacté- rias infectadas. A maior parte da marcação de 32 P estava pre- sente nas bactérias infectadas. A progênie de partí- culas virais produzidas pela infecção continha ~30% da marcação original de 32 P. A progênie recebeu muito pouco – menos de 1 % – da proteína conti- da na população original de fagos. Os capsídeos virais consistem em proteínas e, portanto, apresen- tavam a marcação com 35 S radioativo. Portanto, este experimento mostrou diretamente que apenas o DNA dos fagos parentais entra nas bactérias e en- tão se torna parte da progênie viral, o que corres- ponde exatamente ao padrão de hereditariedade es- perado para o material genético. Um fago se reproduz comandando a maquina- ria de uma célula hospedeira infectada, no sentido de que esta produza mais cópias suas. O fago pos-
FIGURA 1.4 O DNA das bactérias do tipo S pode transformar as bactérias do tipo R em bactérias do mesmo tipo S.
Bactérias S vivas recuperadas do camundongo morto DNA extraído
Bactérias R Transformação Bactérias S
O princípio transformante é o DNA
Camundongo injetado com bactérias S mortas pelo calor e bactérias R vivas
sui um material genético cujo comportamento é
análogo ao apresentado por genomas celulares: seus
traços são reproduzidos de forma fiel e são subme-
tidos às mesmas regras que governam a hereditarie-
dade. O caso do T2 reforça a conclusão geral de
que o DNA é o material genético, sendo este parte
do genoma de uma célula ou de um vírus.
1.4 O DNA É o Material Genético
das Células Animais
Conceitos Essenciais
- O DNA pode ser utilizado para introduzir novas carac- terísticas genéticas em células animais ou em animais inteiros.
- Em alguns vírus, o material genético é RNA.
Quando DNA é adicionado a populações de célu-
las eucarióticas em cultura, o ácido nucléico entra
na célula e, em algumas delas, resulta na produção
de novas proteínas. Quando um DNA purificado é
usado, sua incorporação leva à produção de uma
proteína em particular. A FIGURA 1.6 representa um
dos sistemas-padrão.
Apesar de, por razões históricas, esses experi-
mentos serem denominados transfecção, quando
FIGURA 1.5 O material genético do fago T2 é DNA.
Apenas DNA é herdado pela progênie de fagos
As bactérias infectadas contêm 70% da marca de 32 P
A progênie de fagos têm 3 0% da marca de 32 P e menos de 1% da marca de 35 S
Bactérias infectadas com fagos marcados
Separação dos capsídeos e das bactérias
Os capsídeos contêm 8 0% da marca de 35 S Partículas^ virais^ da progênie são separadas
(^35) S na Proteína
(^32) P no DNA
Adição do DNA TK +
Células que não possuem o gene TK não conseguem produzir timidina-quinase e morrem na ausência de timidina
Algumas células incorporam o gene TK; descendentes de uma célula transfectada se acumulam, originando uma colônia
Células mortas Células vivas
A transfecção introduz DNA novo em células
Colônia de células TK +
realizados em células eucarióticas, são um equiva- lente direto da transformação bacteriana. O DNA que é introduzido na célula receptora se torna par- te do seu material genético e é herdado da mesma forma que qualquer outra parte. Sua expressão con- fere um novo traço às células (síntese de timidina- quinase, no exemplo da Figura 1.6). Inicialmente, esses experimentos foram bem-sucedidos apenas com células individuais, adaptadas ao crescimento em meio de cultura. Desde então, no entanto, DNA foi introduzido em óvulos de camundongos por microinjeção, tornando-se uma parte estável do ma- terial genético do camundongo. Tais experimentos mostram diretamente que além do DNA ser o material genético de eucario- tos, ele pode ser transferido entre diferentes espécies e ainda permanecer funcional. O material genético de todos os organismos co- nhecidos e de muitos vírus é o DNA. Entretanto, alguns vírus usam um tipo alternativo de ácido nu- cléico, o ácido ribonucléico (RNA) , como material genético. Assim, o princípio geral em relação à na- tureza do material genético é tal que este é sempre um ácido nucléico, o DNA, exceto nos casos dos vírus de RNA.
FIGURA 1.6 Células eucarióticas podem adquirir um novo fe- nótipo como resultado de transfecção pela adição de DNA.
1.4 O DNA É O MATERIAL GENÉTICO DAS CÉLULAS ANIMAIS 5
FIGURA 1.7 Uma cadeia polinucleotídica consiste em uma série de liga- ções de açúcar-fosfato 5’-3’ que forma um esqueleto a partir do qual as bases se projetam.
O diâmetro constante da hélice pode ser explicado se as bases em cada cadeia estive- rem voltadas para o interior e restrinjam-se de tal forma que uma purina está sempre em oposição a uma pirimidina, evitando o pareamento de purina-purina (muito largo) e pirimidina-pirimidina (muito estreito).
- Independente das quantidades absolutas de cada base, a proporção de G é sempre igual à proporção de C no DNA, e a proporção de A é sempre a mesma de T. Assim, a com- posição de qualquer DNA pode ser descrita pela proporção de suas bases, isto é G + C. Essa proporção varia de 26 a 74% nas dife- rentes espécies.
Watson e Crick propuseram que as duas ca-
deias, polinucleotídicas na dupla hélice se asso-
ciavam por pontes de hidrogênio entre as bases ni-
trogenadas. G pode formar especificamente pon-
tes de hidrogênio apenas com C, e A pode for-
mar especificamente com T. Essas reações são des-
critas como pareamento de bases, e as bases pa-
readas (G com C, ou A com T) são ditas com-
plementares.
O modelo propôs ainda que as duas cadeias po-
linucleotídicas corriam em direções opostas ( anti-
paralelas ), como ilustrado na FIGURA 1.8. Obser-
vando-se ao longo da hélice, uma fita corre na dire-
ção 5´ para 3´, e a outra, de 3´ para 5´.
O esqueleto de açúcar-fosfato situa-se na face
externa e apresenta cargas negativas nos grupamen-
tos fosfato. Quando o DNA está em solução in vi-
tro , as cargas são neutralizadas pela ligação de íons
metálicos, tipicamente Na +^. Na célula, proteínas
carregadas positivamente contribuem com parte da
força de neutralização. Essas proteínas desempe-
nham um importante papel na determinação da or-
ganização do DNA na célula.
As bases encontram-se voltadas para o interior.
Estas são estruturas planas, que se dispõem aos pa-
res, perpendiculares ao eixo da hélice. Consideran-
do a dupla hélice como uma escada em espiral, os
pares de bases seriam os degraus, conforme ilustra-
do esquematicamente na FIGURA 1.9. Prosseguin-
do ao longo da hélice, as bases estão empilhadas
umas sobre as outras, semelhante a uma pilha de
pratos.
Cada par de bases é rotado em aproximadamente
36 o^ ao redor do eixo da hélice, em relação ao próxi-
mo par. Assim, ~10 pares de bases formam uma
volta completa de 360 o^. A torção das duas fitas for-
ma uma dupla hélice contendo um sulco menor
(~12 Å de largura) e um sulco maior (~22 Å de
largura), como pode ser visto no modelo em escala
Base de pirimidina
Subunidade de nucleotídeo
Base de purina
Esqueleto de açúcar-fosfato
Um polinucleotídeo apresenta uma estrutura repetitiva
Ligações fosfodiéster 5 ’ - 3 ’
H 2 C
H 2 C
O
O
H 2 C O
O
OP^ O
O
O
OP^ O
O
O
O P^ O
O
O
Os fosfatos têm carga negativa
O interior é hidrofóbico
DNA
Ponte de hidrogênio
ESQUELETO
DE AÇÚCAR-
FOSFATO
o
o
o
o
o
o
o
o
o
o
O
O
T
C G
A
O
O
T
G C
A
H 3 C
H H
H H H H H H H N
N N^ N
N
H N
H
HN H
N H O
N
N
N
N N
N
H H
H
H
H H
N
H H
CH 3
H
H
N
N N
N
N
N H
N
N
H
H
H
N
N N
N
N
N N
O
O
O
A dupla hélice apresenta largura constante
FIGURA 1.8 A dupla hélice mantém uma largura constante, porque as purinas sempre se posicionam frente a pirimidinas nos pares de bases complementares A-T e G-C. A seqüência na figura é T-A, C-G, A-T, G-C.
1.6 O DNA É UMA DUPLA HÉLICE 7
8 CAPÍTULO 1 OS GENES SÃO DNA
da FIGURA 1.10. A dupla hélice é dextrógira ; as vol- tas se dão no sentido horário ao longo do eixo da hélice. Essas características representam o modelo aceito para o que é conhecido como forma B do DNA. É importante notar que a forma B representa uma média , e não uma estrutura precisamente es- pecificada. A estrutura do DNA pode sofrer altera- ções locais. Se esta apresenta mais bases por voltas é denominada superenovelada ; se apresenta menos bases por volta é denominada relaxada. O enovela- mento local pode ser afetado pela conformação glo- bal da dupla hélice de DNA no espaço ou pela liga- ção de proteínas em sítios específicos.
1.7 A Replicação do DNA É
Semiconservativa
Conceitos Essenciais
- O experimento de Meselson-Stahl empregou marcação por densidade para provar que uma única cadeia poli- nucleotídica é a unidade de DNA que é conservada du- rante a replicação.
- Cada fita de um duplex de DNA atua como molde na síntese de uma fita filha.
- As seqüências das fitas filhas são determinadas pelo pareamento de bases complementares com as fitas pa- rentais separadas.
É crucial que o material genético seja reproduzido de forma precisa. As duas fitas polinucleotídicas são ligadas apenas por pontes de hidrogênio; assim, são capazes de se separar sem requerer quebras de liga- ções covalentes. A especificidade do pareamento de bases sugere que cada uma das fitas parentais se- paradas possa agir como fita molde na síntese de uma fita filha complementar. A FIGURA 1.11 mos- tra o princípio pelo qual uma fita filha é monta- da sobre cada uma das fitas parentais. A seqüên- cia da fita filha é determinada pela fita parental, um A na fita parental promove a colocação de um T na fita filha, um G parental direciona a in- corporação de um C na fita filha, e assim suces- sivamente. A parte superior da Figura 1.11 ilustra um du- plex parental (não-replicado) que consiste nas duas fitas parentais originais. A parte inferior mostra os dois dúplices filhos que estão sendo produzidos por meio do pareamento de bases complementares. Cada um dos dúplices filhos possui uma seqüência idêntica à parental original, contendo uma fita pa-
GG
CC
T A
GG
CC
G
C GGG
T A
G
C
Açúcar Base Fosfato
Pares de bases planos conectam as fitas de DNA
FIGURA 1.9 Os pares de bases planos estão perpendiculares ao esqueleto de açúcar-fosfato.
Sulco maior
A dupla hélice de DNA possui dois sulcos
Sulco menor
Diâmetro de 20 ÅDiâmetro de 20 Å
Largura de 12 ÅLargura de 12 Å
Largura de 22 ÅLargura de 22 Å
4 Å por volta da hélice
FIGURA 1.10 As duas fitas de DNA formam uma dupla hélice. Photo© Photodisc.
10 CAPÍTULO 1 OS GENES SÃO DNA
no segmento do duplex de DNA é separado em fitas simples, em qualquer momento. A estrutura helicoidal de uma molécula de DNA envolvida na replicação é ilustrada na FIGURA 1.. A região não replicada consiste no duplex parental, que se abre na região replicada, onde os dois dúpli- ces filhos se formaram. A estrutura em dupla hélice é rompida na junção entre as duas regiões, a qual é denominada forquilha de replicação. A replicação envolve a movimentação da forquilha de replica- ção ao longo do DNA parental; assim, ocorre um desenovelamento contínuo das fitas parentais e um enovelamento dos dúplices filhos. A síntese de ácidos nucléicos é catalisada por enzimas específicas, que reconhecem o molde e rea- lizam a tarefa de catalisar a adição de subunidades à cadeia polinucleotídica que está sendo sintetiza- da. As enzimas são denominadas de acordo com o tipo de cadeia que está sendo sintetizada: as DNA
polimerases sintetizam DNA e as RNA polimera- ses sintetizam RNA. A degradação de ácidos nucléicos também re- quer enzimas específicas: desoxirribonucleases ( DNAases ) degradam DNA e ribonucleases ( RNAases ) degradam RNA. As nucleases são agrupadas nas classes gerais de exonucleases e en- donucleases :
- As endonucleases clivam ligações no interior de moléculas de RNA ou DNA, gerando fragmentos distintos. Algumas DNAases cli- vam as duas fitas de um duplex de DNA no sítio-alvo, e outras clivam apenas uma das duas fitas. As endonucleases estão envolvi- das em reações de clivagem conforme apre- sentado na FIGURA 1..
- As exonucleases removem sucessivos resíduos individuais da extremidade da molécula, ge- rando mononucleotídeos. Elas sempre atuam em uma única fita de ácido nucléico, sendo que cada exonuclease prossegue em uma di- reção específica, isto é, a partir da extremi- dade 5’ ou 3’, prosseguindo em direção à outra extremidade. Tais enzimas estão en- volvidas em reações que aparam as extremi- dades, como mostrado na FIGURA 1..
1.9 A Informação Genética Pode Ser
Fornecida pelo DNA ou RNA
Conceitos Essenciais
- Os genes celulares são de DNA, embora vírus e viróides possam apresentar genomas de RNA.
- O DNA é convertido em RNA pela transcrição, e o RNA pode ser convertido em DNA pela transcrição reversa.
- A tradução do RNA em proteína é unidirecional.
O dogma central define o paradigma da biologia molecular. Os genes são perpetuados como seqüên- cias de ácidos nucléicos, mas atuam sendo expres- sos sob a forma de proteínas. A replicação é res- ponsável pela herança da informação genética. A transcrição e a tradução são responsáveis pela con- versão dessa informação de uma forma para outra. A FIGURA 1.16 ilustra os papéis da replicação, da transcrição e da tradução, vistas pela perspectiva do dogma central:
- A perpetuação do ácido nucléico pode envolver tanto DNA ou RNA como material genético. As células utilizam apenas DNA. Alguns ví- rus usam RNA, e a replicação do RNA viral ocorre na célula infectada.
FIGURA 1.15 Uma exonuclease remove uma base de cada vez, clivando a última ligação em uma cadeia polinucleotídica.
FIGURA 1.14 Uma endonuclease cliva uma ligação no interior de um ácido nucléico. Esse exemplo mostra uma enzima que ataca uma fita de um duplex de DNA.
Endonucleases atacam ligações internas
Ligação quebrada
DNAs replicados
DNA parental
Forquilha de replicação
Uma forquilha de replicação move-se ao longo do DNA
FIGURA 1.13 A forquilha de replicação é a região do DNA na qual há uma transição entre o duplex parental desenovelado e os dúplices filhos recém-sintetizados.
Exonucleases degradam a partir das extremidades
FIGURA 1.16 O dogma central afirma que a informação no ácido nucléico pode ser perpetuada ou transferida, mas a trans- ferência de informação em proteína é irreversível.
- A expressão da informação genética celular é geralmente unidirecional. A transcrição do DNA gera moléculas de RNA que podem ser usadas em seguida apenas para gerar se- qüências de proteínas; via de regra, os RNAs não podem ser recuperados e usados como informação genética. A tradução do RNA em proteínas é sempre irreversível.
Esses mecanismos são igualmente efetivos em
relação à informação genética de procariotos e de
eucariotos e para a informação carreada por vírus.
Os genomas de todos os seres vivos consistem em
um duplex de DNA. Vírus têm genomas que con-
sistem em DNA ou RNA, havendo exemplos de
cada um dos tipos, que são fitas duplas (fd) ou fitas
simples (fs). Detalhes do mecanismo usado para re-
plicar o ácido nucléico variam dentre os sistemas
virais, mas o princípio da replicação via síntese de
fitas complementares é o mesmo, como ilustrado
na FIGURA 1..
Os genomas celulares reproduzem o DNA pelo
mecanismo de replicação semiconservativa. Os ge-
nomas virais de fita dupla, DNA ou RNA, tam-
bém se replicam usando as fitas individuais do du-
plex como moldes para a síntese das fitas comple-
mentares.
Os vírus com genomas de fita simples usam a
fita simples como molde para sintetizar uma fita
complementar; essa fita complementar é, por sua
vez, utilizada para sintetizar o seu complemento,
que é, obviamente, idêntico à fita original. A repli-
cação pode envolver a formação de intermediários
DNA
RNA
Proteína
Replicação
Replicação
Transcrição reversa
Transcrição
Tradução
O dogma central descreve o fluxo da informação
A fita simples parental é usada para sintetizar a fita complementar
A fita complementar é usada para sintetizar uma cópia da fita parental
Os ácidos nucléicos replicam-se por meio de fitas complementares
Molde de fita dupla Fitas novas
Fita antiga
Fita antiga
A replicação gera dois dúplices filhos, cada um contendo uma fita parental e uma fita recém-sintetizada
Molde de fita simples
FIGURA 1.17 Ácidos nucléicos de fita dupla e de fita simples replicam-se pela síntese de fitas complementares, dirigida pe- las regras do pareamento de bases.
estáveis de fita dupla ou o uso de ácidos nucléicos de fita dupla somente como um estágio transitório. A restrição em relação à transferência unidire- cional de DNA para RNA não é absoluta. Esta é superada pelos retrovírus , cujos genomas consis- tem em moléculas de RNA de fita simples. Duran- te o ciclo infectivo, o RNA é convertido pelo pro- cesso de transcrição reversa em um DNA de fita simples, o qual é, por sua vez, convertido em um DNA de fita dupla. Esse duplex de DNA torna-se parte do genoma da célula, sendo herdado como qualquer outro gene. Assim, a transcrição reversa per- mite que uma seqüência de RNA seja recuperada e utilizada como informação genética. A existência da replicação de RNA e da trans- crição reversa estabelece o princípio geral de que a informação sob a forma de seqüências de qualquer um dos tipos de ácido nucléico pode ser convertida no outro tipo. No entanto, durante o curso habitual dos eventos, a célula baseia-se nos processos de re- plicação do DNA, de transcrição e de tradução. En- tretanto, em ocasiões raras (possivelmente media- das por um vírus de RNA), a informação de um RNA celular é convertida em DNA e inserida no genoma. Embora a transcrição reversa não desem- penhe um papel nas operações regulares da célula, esta torna-se um mecanismo de potencial importância quando consideramos a evolução do genoma. Os mesmos princípios são seguidos na perpe- tuação da informação genética de genomas enor-
1.9 A INFORMAÇÃO GENÉTICA PODE SER FORNECIDA PELO DNA OU RNA 11
vitro. As forças não-covalentes que estabilizam a
dupla hélice são rompidas pelo aquecimento ou pela
exposição a baixas concentrações de sal. As duas
fitas de uma dupla hélice separam-se completamente
quando as pontes de hidrogênio entre elas são des-
feitas.
O processo de separação das fitas é denomina-
do desnaturação ou (mais coloquialmente) fusão.
(O termo “desnaturação” é também utilizado para
descrever a perda da estrutura verdadeira de uma
proteína; este é um termo geral e implica que a con-
formação natural de uma macromolécula foi con-
vertida em alguma outra forma.)
A desnaturação do DNA ocorre em uma faixa
estreita de temperatura e promove modificações
profundas em muitas de suas propriedades físicas.
O ponto médio da faixa de temperatura na qual as
fitas de DNA se separam é denominado temperatu-
ra de fusão ( T m ) (do inglês: melting temperature ).
Essa temperatura é dependente da proporção de
pares de bases G-C. Uma vez que o par G-C apre-
senta três pontes de hidrogênio, é mais estável que
um par A-T, que apresenta apenas duas pontes de
hidrogênio. Quanto maior o conteúdo de pares de
bases G-C, maior será a energia necessária para se-
parar as duas fitas. Em solução, sob condições fi-
siológicas, um DNA que contém 40% de G-C – um
valor típico de genomas de mamíferos – se desnatura
a uma T m de cerca de 87oC. Assim, o duplex de DNA
é estável em temperaturas existentes na célula.
A desnaturação do DNA é reversível sob condi-
ções apropriadas. A capacidade das duas fitas com-
plementares separadas formarem novamente uma
dupla hélice é denominada renaturação. A renatu-
ração depende do pareamento de bases específico
entre as fitas complementares. A FIGURA 1.20 mos-
tra que a reação ocorre em dois estágios. Inicial-
Pareamento intermolecular entre RNAs longos e curtos
RNA curto
RNA longo
Pareamento intramolecular no interior do RNA
DNA
RNA
Tanto o DNA como o RNA podem formar dúplices
DNA
FIGURA 1.19 O pareamento de bases ocorre no duplex de DNA e também em interações intra- e intermoleculares no RNA (ou DNA) de fita simples.
mente, as fitas simples do DNA em solução encon- tram-se aleatoriamente; se suas seqüências forem complementares, as duas fitas pareiam suas bases e geram uma região curta de dupla hélice. Em segui- da, a região de pareamento de bases estende-se ao longo da molécula por um efeito semelhante a um zíper, formando uma longa molécula de duplex. A renaturação da dupla hélice restaura as propriedades originais perdidas quando o DNA foi desnaturado. A renaturação descreve a reação entre duas se- qüências complementares que foram separadas por desnaturação. Entretanto, a técnica pode ser esten- dida para permitir que duas seqüências complemen- tares de quaisquer ácidos nucléicos reajam entre si e formem estruturas em duplex. Isso algumas é ve- zes chamado de anelamento , embora a reação seja geralmente descrita como hibridização , sempre que ácidos nucléicos de diferentes origens estão envol- vidos, como no caso quando uma das preparações consiste em DNA, e a outra, em RNA. A capacida- de de duas preparações de ácidos nucléicos hibridiza- rem constitui um teste preciso de sua complementari- dade, visto que apenas seqüências complementares po- dem forma uma estrutura em duplex. O princípio da reação de hibridização baseia-se na exposição de duas preparações de ácidos nucléi- cos de fita simples uma à outra, medindo-se em seguida a quantidade de material em fita dupla for- mado. A FIGURA 1.21 ilustra um procedimento no qual uma preparação de DNA é desnaturada e as fitas simples são adsorvidas a um filtro. Então, uma segunda preparação de DNA (ou RNA) desnatura- do é adicionada. O filtro é tratado de forma que a segunda preparação pode adsorver-se a ele somen-
FIGURA 1.20 Fitas simples de DNA desnaturado podem ser renaturadas, originando a forma de duplex.
O DNA pode ser desnaturado e renaturado
DNA de fita dupla
DNA de fita Simples
DNA
renaturado
Desnaturação
Renaturação
1.10 OS ÁCIDOS NUCLÉICOS HIBRIDIZAM-SE POR PAREAMENTO DE BASES 13
14 CAPÍTULO 1 OS GENES SÃO DNA
te se for capaz de parear suas bases com o DNA que foi inicialmente adsorvido. Geralmente, a se- gunda preparação é marcada radioativamente, de maneira que a reação pode ser medida pela quanti- dade de marcação radioativa retida pelo filtro. A extensão da hibridização entre dois ácidos nu- cléicos de fita simples é determinada por sua com- plementaridade. Duas seqüências não precisam ser perfeitamente complementares para hibridizarem. Se estas forem bastante relacionadas, mas não idênti- cas, um duplex imperfeito é formado, no qual o pareamento de bases é interrompido nas posições onde as duas fitas não são correspondentes.
1.11 As Mutações Modificam a
Seqüência do DNA
Conceitos Essenciais
- Todas as mutações consistem em modificações na se- qüência do DNA.
- Mutações podem ocorrer espontaneamente ou serem induzidas por agentes mutagênicos.
As mutações fornecem evidências decisivas de que o DNA é o material genético. Quando uma modi- ficação na seqüência de DNA provoca uma altera-
O DNA é desnaturado em solução
O DNA é desnaturado e adsorvido ao filtro
O filtro é mergulhado na solução
O DNA ligado ao filtro é medido
O ensaio em filtro mede a complementaridade
FIGURA 1.21 A hibridização no filtro estabelece se uma solu- ção de DNA (ou RNA) desnaturado contém seqüências comple- mentares às fitas imobilizadas no filtro.
ção na seqüência de uma proteína, podemos con- cluir que o DNA codifica aquela proteína. Além disso, uma modificação no fenótipo do organismo pode permitir-nos identificar a função da proteína. A existência de muitas mutações em um gene pode possibilitar a comparação entre as muitas formas variantes de uma dada proteína, e uma análise de- talhada pode ser usada para identificar as regiões da proteína responsáveis por funções enzimáticas individuais, ou outras funções. Todos os organismos estão sujeitos a um certo número de mutações como resultado de operações celulares normais ou interações aleatórias com o am- biente. Essas são denominadas mutações espontâ- neas ; a sua taxa de ocorrência é característica de um determinado organismo, sendo algumas vezes denominada nível basal. As mutações são eventos raros e, obviamente, aquelas que causam danos em um gene são selecionadas negativamente durante a evolução. Assim, é difícil obter-se grandes núme- ros de mutantes espontâneos para um estudo, a partir de populações naturais. A ocorrência de mutações pode ser aumentada pelo tratamento com determinados compostos. Estes são chamados de compostos mutagênicos , e as modificações que causam são referidas como mutações induzidas. A maioria dos compostos mu- tagênicos atua diretamente devido a sua capacidade de modificar uma determinada base do DNA, ou de incorporar-se ao ácido nucléico. A eficiência de um composto mutagênico é avaliada pelo aumento da taxa de mutações em relação aos níveis basais. A partir do uso de compostos mutagênicos, tornou-se possível a indução de muitas modificações em qualquer gene. Mutações espontâneas que inativam a função gênica ocorrem em bacteriófagos e bactérias em uma taxa relativamente constante de 3-4 x 10 -3^ por ge- noma, por geração. Dada a grande variação no ta- manho dos genomas entre bacteriófagos e bacté- rias, essa taxa corresponde a grandes diferenças na taxa de mutação por par de bases. Isso sugere que a taxa global de mutações esteve sujeita a forças sele- tivas que equilibraram os efeitos deletérios da maio- ria das mutações e os efeitos vantajosos de algumas mutações. Essa conclusão é reforçada pela observa- ção de que um microrganismo do domínio Archaea, que habita ambientes em condições extremas de temperatura e acidez (condições que normalmente danificariam o DNA), não apresenta uma taxa ele- vada de mutação e, de fato, tem uma taxa um pou- co abaixo da taxa média. A FIGURA 1.22 mostra que a taxa de mutações em bactérias corresponde a cerca de 10-6^ eventos por ló- cus, por geração, ou a uma taxa média de 10-9^ a 10- de modificação por par de bases, por geração. A taxa
16 CAPÍTULO 1 OS GENES SÃO DNA
ilustra o exemplo da bromouracila (BrdU), um aná- logo de timina que contém um átomo de bromo no lugar do grupo metil da timina. A BrdU é in- corporada no DNA no lugar da timina. Entretan- to, essa base apresenta propriedades ambíguas de pareamento, devido à presença do átomo de bro- mo que permite a ocorrência de uma mudança na qual a base altera sua estrutura de um grupo ceto (=O) para uma forma enol (-OH). A forma enol pode parear com guanina, o que leva à substituição do par original A-T por um par G-C. O pareamento incorreto pode ocorrer tanto du- rante a incorporação original da base ou no ciclo de replicação subseqüente. A transição é induzida com uma certa probabilidade a cada ciclo replicati- vo; por isso, a incorporação de BrdU tem efeitos continuados na seqüência de DNA.
As mutações pontuais foram consideradas du- rante um longo tempo como a principal forma de modificação de genes individuais. No entanto, sa- bemos que as inserções de segmentos de material adicional são bastante freqüentes. A fonte de mate- rial inserido normalmente corresponde a elemen- tos transponíveis , que são seqüências de DNA ca- pazes de se mover de um sítio para o outro (Ver Capítulo 21, Transposons, e Capítulo 22, Retroví- rus e Retrotransposons). Uma inserção geralmente abole a atividade de um gene. Quando tais inser- ções ocorrem, deleções de parte ou de todo o ma- terial inserido, e algumas vezes de regiões adjacen- tes, podem ocorrer subseqüentemente. Uma diferença significativa entre mutações pon- tuais e inserções/deleções é que a freqüência de mu- tações pontuais pode ser aumentada por compos- tos mutagênicos, e a ocorrência de modificações causadas por elementos transponíveis não é afeta- da. No entanto, inserções e deleções podem tam- bém ocorrer por outros mecanismos – por exem- plo, aqueles envolvendo erros durante replicação ou recombinação –, apesar de, provavelmente, se- rem menos comuns. Além destes, uma classe de compostos mutagênicos denominados acridinas in- troduz inserções e deleções (muito pequenas).
1.13 Os Efeitos das Mutações Podem Ser Revertidos
Conceitos Essenciais
- As mutações diretas inativam um gene, e as mutações reversas (ou revertentes) revertem seus efeitos.
- As inserções podem ser revertidas pela deleção do mate- rial inserido, ao passo que as deleções não são revertidas.
- A supressão ocorre quando uma mutação em um segun- do gene anula o efeito da mutação do primeiro gene.
A FIGURA 1.25 mostra que o isolamento de rever- tentes é uma característica importante que distin- gue as mutações pontuais e inserções das deleções:
- Uma mutação pontual pode ser revertida pela restauração da seqüência original ou pelo ganho de uma mutação compensatória em outro local do gene.
- Uma inserção de material adicional pode ser revertida pela deleção do material inserido.
- Uma deleção de parte de um gene não pode ser revertida.
Mutações que inativam um gene são denomi- nadas mutações diretas. Seus efeitos são revertidos por mutações reversas , que são dois tipos: rever- sões verdadeiras ou reversões de segundo sítio.
FIGURA 1.24 Mutações podem ser induzidas pela incorporação de análogos de bases no DNA.
N N
C
C
C
HC
H
Açúcar
Br
N N
C
C
C
HC
OH
Açúcar
Br
N N
C
C
C
HC
C
N
N
C N
CH
C CN
H
H
H
H
N
Açúcar
Açúcar
Br
A mudança de ceto para enol permite a BrdU parear com G
Mudança ceto-enol
O
O O
O
O
O
Par BrdU-G
BrdU pareia-se com G na replicação
HC
HC
N
N
N
N
N
N
CH
CH
N
N
C
C
C
C
N
N
N
N
C
C
C
C
C
C
HC
HC
H
H
N
N
H
H
H
H
Açúcar
Açúcar
Açúcar
Açúcar
Br
C
C
O
O
O
O
Par BrdU-A
Par T-A
CH 3
BrdU pareia-se com A na replicação
BrdU provoca a troca do par A-T por um G-C
Uma reversão exata da mutação original é cha-
mada de reversão verdadeira. Assim, se um par A-
T foi substituído por um par G-C, uma outra mu-
tação que restaure o par A-T irá regenerar exata-
mente a seqüência selvagem.
O segundo tipo de mutação reversa – a rever-
são de segundo sítio – pode ocorrer em outro local
no gene e seu efeito compensa a primeira mutação.
Por exemplo, uma modificação de um aminoácido
em uma proteína pode abolir a função de um gene,
mas uma segunda alteração pode compensar a pri-
meira e restaurar a atividade da proteína.
Uma mutação direta resulta de qualquer mu-
dança que inative um gene; uma mutação rever-
sa deve restaurar a função de uma proteína dani-
ficada por uma mutação direta em particular. A
taxa de mutação reversa é correspondentemente
inferior à de mutação direta, tipicamente na or-
dem de ~10.
Mutações também podem ocorrer em outros ge- nes, de modo a contornar os efeitos da mutação no gene original. Esse efeito é denominado supressão. Um lócus no qual uma mutação suprime o efeito de uma mutação em outro lócus é chamado de su- pressor.
1.14 As Mutações São Concentradas
em Hotspots
Conceitos Essenciais
- A freqüência de uma mutação em qualquer par de bases é determinada por uma flutuação estatística, exceto nos hotspots (“sítios quentes”), onde a freqüência é aumentada em pelo menos uma ordem de grandeza.
Até agora lidamos com mutações em relação às mudanças individuais na seqüência do DNA que influenciam a atividade da unidade genética na qual estas ocorrem. Quando consideramos as mutações no que tange à inativação do gene, a maioria dos genes de uma espécie revela taxas mais ou menos similares de mutação em relação ao seu tamanho. Isso sugere que o gene pode ser considerado como um alvo de mutação, e que danos em qualquer par- te de um gene podem abolir sua função. Como re- sultado, a suscetibilidade à mutação é aproximada- mente proporcional ao tamanho do gene. Ao con- siderarmos os sítios de mutações no interior da se- qüência de DNA: todos os pares de bases em um gene são igualmente suscetíveis, ou existem alguns mais prováveis de serem mutados que outros? O que acontece quando isolamos um grande número de mutações independentes em um mes- mo gene? Muitos mutantes são obtidos. Cada um é resultado de um evento mutacional individual. Então, o sítio de cada mutação é determinado. A maioria das mutações ocorrerá em sítios diferentes, mas algumas ocorrerão na mesma posição. Duas mutações independentemente isoladas em um mes- mo sítio podem corresponder exatamente à mesma alteração no DNA (neste caso, o mesmo evento mu- tacional ocorreu em mais de uma ocasião), ou po- dem corresponder a modificações diferentes (três mutações pontuais diferentes são possíveis para cada par de bases). O histograma da FIGURA 1.26 revela a freqüên- cia com a qual mutações são encontradas em cada par de bases do gene lacI de E. coli. A probabilida- de estatística de mais de uma mutação ocorrer em um sítio particular é dada pela cinética de acerto alea- tório (como observado na distribuição de Poisson). Assim, alguns sítios apresentarão uma, duas ou três mutações, e outros, nenhuma. Alguns sítios apre-
FIGURA 1.25 Mutações pontuais e inserções podem ser rever- tidas, mas deleções não são revertidas.
Mutações pontuais
Reversão
Inserção
Reversão por deleção
ATCGGACGGTTA
TAGCCTGCCAAT
Nenhuma reversão é possível
Algumas mutações podem ser revertidas
ATCGGACTTACCGGTTA
TAGCCTGAATGGCCAAT
ATCGGACTTACCGGTTA
TAGCCTGAATGGCCAAT
ATCGGACTTXXXXXACCGGTTA
TAGCCTGAAYYYYYTGGCCAAT
ATCGGACTTACCGGTTA
TAGCCTGAATGGCCAAT
ATCGGAC0ACCGGTTA
TAGCCTGAGTGGCCAAT
ATCGGACTTACCGGTTA
TAGCCTGAATGGCCAAT
ATCGGACTTACCGGTTA
TAGCCTGAATGGCCAAT
1.14 AS MUTAÇÕES SÃO CONCENTRADAS EM HOTSPOTS 17
do próximo ciclo de replicação, ocorrerá uma mu-
tação. Na replicação seguinte, as bases incorretas
do par G-T serão separadas e formarão pares com
novas bases, produzindo um par G-C selvagem e
um par mutante A-T.
A desaminação da 5-metilcitosina é a causa mais
comum de formação de pares G-T incorretos no
DNA. Sistemas de reparo que atuam em erros do
tipo G-T tendem a repor o T por um C (ao invés
da reposição alternativa de repor o G por um A), o
que ajuda a reduzir a taxa de mutação (ver Seção
20.7, O Controle da Direção do Reparo de Parea-
mentos Incorretos). No entanto, esses sistemas não
são tão efetivos quanto a remoção do U do par G-
U incorreto. Assim, a desaminação da 5-metilcito-
sina promove mutações com muito mais freqüên-
cia que a desaminação da citosina.
A 5-metilcitosina também cria hotspots no DNA
eucariótico. Isto é comum em dinucleotídeos CpG
que estão concentrados em regiões denominadas
ilhas CpG (ver Seção 24.19, Ilhas CpG São Alvos
Regulatórios). Embora a 5-metilcitosina correspon-
da a apenas cerca de 1% das bases do DNA huma-
no, sítios contendo esta base modificada são res-
ponsáveis por ~30% de todas as mutações pontuais.
Isso torna o estado de 5-metilcitosina um determi- nante particularmente importante de mutações em células animais. A importância dos sistemas de reparo na redu- ção da taxa de mutações é enfatizada pelos efeitos decorrentes da eliminação da enzima MBD4 de camundongo, uma glicosilase que remove T (ou U) de pares formados incorretamente com G. O resul- tado é o aumento, com um fator de 3×, da taxa de mutação nos sítios CpG. (Esse efeito não é ainda maior porque MBD4 é apenas um dos vários siste- mas que atuam em pares incorretos G-T; podemos imaginar que a eliminação de todos os sistemas aumentaria muito mais a taxa de mutações.) A atuação desses sistemas traz uma luz interes- sante em relação ao uso de T no DNA comparado ao uso de U no RNA. Talvez essa propriedade este- ja relacionada à necessidade de estabilidade da se- qüência no caso do DNA; o uso de T implica que quaisquer desaminações em C são imediatamente reconhecidas, pois geram uma base (U), a qual em geral não está presente no DNA. Isso aumenta mui- to a eficiência com a qual os sistemas de reparo po- dem atuar (se comparado à situação onde estes de- vem reconhecer erros G-T, os quais podem ser pro- duzidos também por situações onde a remoção do T não seria a resposta adequada). Além disso, a li- gação fosfodiéster do esqueleto é mais lábil quando a base é U.
1.16 Alguns Agentes Hereditários São extremamente Pequenos
Conceitos Essenciais
- Alguns agentes hereditários muito pequenos não codi- ficam proteínas e consistem em RNA ou proteína com propriedades hereditárias.
Viróides são agentes infecciosos que causam doen- ças em plantas superiores. Estes correspondem a mo- léculas muito pequenas e circulares de RNA. Dife- rentemente dos vírus – para os quais os agentes in- fecciosos correspondem a um vírion , um genoma encapsulado em um envoltório protéico –, o RNA do viróide é, ele mesmo, o agente infeccioso. O viróide consiste somente em RNA, que exibe um extenso, porém imperfeito, pareamento de bases, e assume uma forma de bastão característica, como mostra- do na FIGURA 1.29. Mutações que interferem com a estrutura de bastão reduzem a sua infectividade. Um RNA de viróide consiste em uma única es- pécie molecular, a qual é replicada de forma autô- noma nas células infectadas. Sua seqüência é preci- samente perpetuada em seus descendentes. Virói-
U
A remoção da uracila impede mutações
Me C
Desaminação oxidativa
G
A uracila é removida
Me-C é substituída por T
T pareia com A
A citosina é inserida
T
T C
C C
A G
e
Não há mutação
C
G
T
Mutação
Replicação
G G
G G
G G
T C
G G
C é substituída por U
FIGURA 1.28 A desaminação da 5-metilcitosina produz timina (por transição C-G para T-A); a desaminação da citosina produz uracila (que normalmente é removida e então é substituída por citosina).
1.16 ALGUNS AGENTES HEREDITÁRIOS SÃO EXTREMAMENTE PEQUENOS 19
20 CAPÍTULO 1 OS GENES SÃO DNA
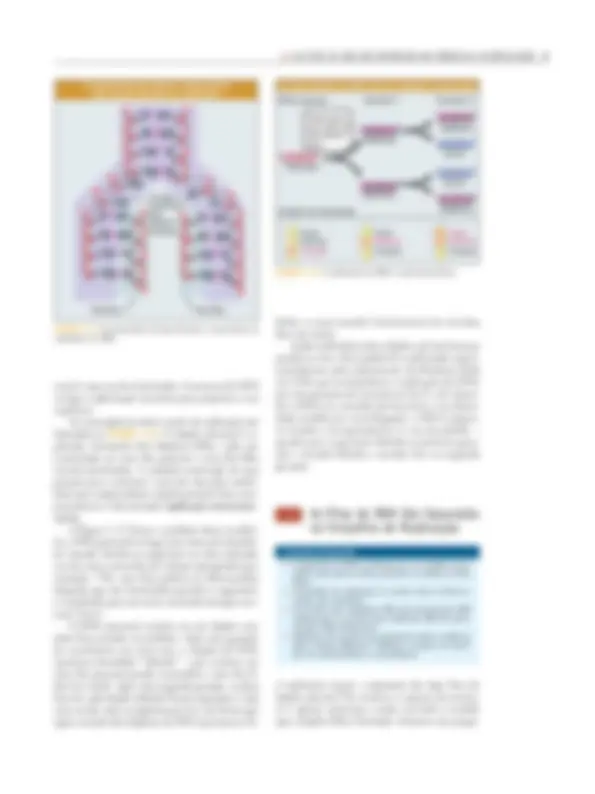
des são classificados em diferentes grupos. Um de- terminado viróide é identificado como membro de um grupo devido à similaridade da sua seqüência com a seqüência de outros membros do grupo. Por exemplo, quatro viróides relacionados ao PSTV (viróide do Tubérculo Afilado de Batata, do inglês potato spindle tuber viroid ) apresentam de 70 a 83% de similaridade de seqüência com PSTV. Diferen- tes isolados de uma mesma linhagem de viróide variam entre si, e a modificação pode afetar o fenó- tipo das células infectadas. Por exemplo, as linha- gens suave e severa de PSTV diferem em três substi- tuições de nucleotídeos. Os viróides se assemelham aos vírus por possuí- rem genomas de ácidos nucléicos herdáveis. Assim, preenchem os critérios em relação à informação gené- tica. Entretanto, os viróides, que por vezes são cha- mados de patógenos subvirais , diferenciam-se dos ví- rus tanto na estrutura quanto na função. O RNA do viróide parece não ser traduzido em proteínas e, desta forma, não é capaz de codificar as funções necessárias à sua sobrevivência. Essa situação leva a duas ques- tões: como o RNA do viróide é replicado e como o viróide afeta o fenótipo da célula vegetal infectada? A replicação deve ser realizada pelas enzimas da célula hospedeira, subvertidas de suas funções nor- mais. A natureza herdável da seqüência do viróide indica que o RNA do viróide atua como molde. Os viróides são presumivelmente patogênicos porque interferem com os processos celulares nor- mais. Esta ação é realizada de forma relativamente aleatória, por exemplo, seqüestrando uma enzima essencial para a sua própria replicação, ou interferin- do com a produção de RNAs celulares necessários. Alternativamente, os viróides podem se comportar como moléculas regulatórias anormais, exercendo efei- tos particulares na expressão de genes individuais. Um agente ainda mais incomum é o scrapie – o causador de uma doença neurológica degenerativa
em ovelhas e cabras. A doença está relacionada às doenças humanas kuru e síndrome de Creutzfeldt- Jacob, que afetam a função cerebral. O agente infeccioso do scrapie não contém ácido nucléico. Esse extraordinário agente é chamado de príon (agente infeccioso proteináceo). Este corres- ponde a uma glicoproteína hidrofóbica de 28 kDa, PrP. PrP é codificada por um gene celular (conser- vado nos mamíferos) que é expresso no cérebro nor- mal. A proteína é encontrada sob duas formas: o produto encontrado no cérebro normal é denomi- nado PrP c^ e é totalmente degradado por proteases. A proteína encontrada em cérebros infectados é denominada PrP sc^ e é extremamente resistente à degradação por proteases. PrPc^ é convertida em PrPsc por uma modificação ou alteração conformacional que confere resistência a proteases, a qual ainda não está completamente definida. O agente do scrapie , PrPsc^ deve, de algum forma, modificar a síntese do seu equivalente celular normal, tornando-o infeccioso, ao invés de inócuo (ver Seção 31.12, Príons Causam Doenças em Mamíferos). Ca- mundongos que não possuem o gene PrP não são in- fectados e não desenvolvem scrapie , demonstrando que PrP é essencial ao desenvolvimento da doença.
1.17 Resumo
Dois experimentos clássicos provaram que o DNA é o material genético. O DNA isolado de uma linhagem das bactérias pneumococos conferiu propriedades dessa linhagem em outra linhagem. Além disso, o DNA é o único componente que é herdado pela progênie de fagos a partir dos fagos parentais. O DNA pode ser usado para transfectar novas propriedades em células eucarióticas. O DNA é uma dupla hélice, consistindo em fitas antiparalelas nas quais as unidades de nucleo-
CUG C
G C
A U
A U
C G
U G
AA U
A U
C G
U A
359
CG C
U G
G C
G C
U A
U A
C G
C G
U U
G
U
U C
G C
G C
U G
U A
C U
A U
C G
A U
C G
C G
U G
G A
A
G
C C
C G
U G
C G
C G
U C
G U
A U
G C
C G
A U
G U
A C
A A
A U
A U
G C
A U
A A
A
U
A
C
A
U
A C
G U
A U
A U
G U
G U
C C
G G
G C
C C
U A
C A
G G
G A
A G
G C
G C
A C
G U
C C
G G
C A
U A
U G
C U
A C
G A
G
A
G
C
A
A
U A
C A
C G
C G
C U
G G
G G
G C
G C
A C
A A
A
U
C
C
C
A
U
U
G C
G G
A G
G C
C U
G U
A C
A G
C C
U U
G G
G U
C C
A G
A C
A G
A U
A U
A U
G C
G C
A C
C C
G C
G G
U C
G U
G C
G C
G C
A C
G A
U C
G C
C A
C G
C G
A A
G C
C G
G C
G C
C C
C G
G C
A G
C C
A U
G C
G C
A U
G U
U C
A
C
A
U
U
G
U U
C G
C G
C G
G C
C U
C U
G C
A U
A U
A U
C C
A C
G U
G U
G U
U U
U U
U U A^ CU 330
1 30
300
60
270
90
240
120 150
(^210 )
PSTV severo
CUG C
G C
A U
A U
C G
U G
AA U
A U
C G
U A
359
CG C
U G
G C
G C
U A
U A
C G
C G
U U
G
U
U C
G C
G C
U G
U A
C U
A U
C G
A U
C G
C G
U G
G A
A
G
C C
C G
U G
C G
C G
U C
G U
A U
G C
C G
A U
G U
A C
A U
A U
A U
G C
A U
A A
A
U
A
C
A
U
A C
G U
A U
A U
G U
G U
C C
G G
G C
C C
U A
C A
G G
G A
A G
G C
G C
A C
G U
C C
G G
C A
U A
U G
C U
A C
G A
G
A
G
C
A
A
U A
C A
C G
C G
C U
G G
G G
G C
G C
A C
A
A
A
U
C
C
C
A
U
U
G C
G G
A G
G C
C U
G U
A C
A G
C C
U U
G G
G U
C C
A G
AU G
A U
A U
G U
G C
A C
C C
G C
G G
U C
G U
G C
G C
G C
A C
G A
U C
G C
C A
C G
C G
A A
G C
C G
G C
G C
C C
C G
G C
A G
C C
A U
G C
G C
A U
G U
U C
A
C
A
U
U
G
U U
C G
C G
C G
G C
C U
C U
G C
A U
A U
A U
C C
A C
G U
G U
G U
U U
U U
U U A^ CU
330
1 30
300
60
270
90
240
120 150
210 180
PSTV suave
U (^) C
Os genomas de viróides são de RNA
+U A U
AA U
FIGURA 1.29 O RNA de PSTV é uma molécula circular que forma uma extensa estrutura em dupla fita, interrompida por muitas alças internas. As formas severa e suave diferem em três sítios.