¡Descarga Resumen inmuno y más Apuntes en PDF de Inmunología solo en Docsity!
¿Qué entiende por repertorio inmunológico? Especifique a nivel celular y molecular. ¿Qué es la teoría de selección clonal y cómo se aplica a la respuesta inmune normal? Discuta acerca de la relevancia del antígeno en los procesos de generación del repertorio de linfocitos B y de memoria inmunológica. ¿Se ven afectados los BCR?
1. a Repertorio inmunológico es el conjunto de ESPECIFICIDADES del que dispone el individuo para enfrentar los posibles ataques externos. Esto incluye la cantidad de clones diferentes de linfocitos, tanto T como B, otras células especializadas como macrófagos, células NK, células dendríticas, y moléculas sintetizadas, expresadas y/ o liberadas por estas células (citoquinas, anticuerpos, quimoquinas, etc).
1b Teoría de selección clonal establece que el sistema
inmune específico está formado por un numeroso conjunto de linfocitos, cada uno con un tipo determinado de receptor para antígenos, único y exclusivo de ese clon (una sola especificidad) y se selecciona en cada respuesta el más afín al antígeno. Esta selección de clones se basa en las características del antígeno.
Ojo esto es distinto a la maduración del linfocito B donde la selección es negativa para evitar la autoinmunidad
El antígeno + selección de clones por afinidad del antígeno al BCR y apoptosis de , destino de Cels B
memoria y efectores
2) ¿Cuáles serían las funciones de los organos linfoides primarios y secundarios? ) Describa detalladamente el
molécula. Así el SI se las arregla para acercar los elementos celulares, desde zonas de menor concentración a zonas de mayor concentración. Una vez que llegan al lugar, estas mismas células poseen otros receptores capaces de interaccionar con moléculas de superficie de células ubicadas en el ganglio, permitiendo ahora su retención. Estas células son específicamente los endoteliocitos de la vénula (ver mas abajo anexo vénulas altas) que secretan primeramente quimioquinas (para atraer a los L vírgenes) y luego expresan en membrana el ligando de selectina-L que es capaz de interaccionar con la molécula selectina-L de la membrana de los linfocitos, reteniendo y acumulando las células vírgenes en el órgano linfoide. Una vez que se han activado y diferenciado, el patrón de expresión de moléculas de membrana del Linfocitos cambia y empiezan a adquirir otros receptores de quimioquinas y receptores de membrana. Esto permite su desplazamiento hacia otros órganos. Estas nuevas moléculas son la selectina-P, selectina-e que interaccionan con moléculas como ICAM-1 y VCAM-1 presentes en el endotelio de los sitios infectados. Entonces el patrón de expresión molecular de los linfocitos varía según sea una célula virgen que esta migrando hacia un linfonodo y es retenida o como un linfocito que ha sido activado y que sale del ganglio hacia el sitio infectado para establecerse allí. El fundamento molecular de estos procesos se divide en dos conceptos:
- atracción por quimioquinas que las orientan y desplazan a sitios determinados,
- presencia de moléculas en dichos sitios que las retienen.
Cuando las células dendríticas están en la periferia se encuentran en una condición inmadura. En este estado presentan receptores para quimioquinas que las direccionan hacia sitios periféricos infectados y además receptores para otras moléculas que las concentran allí. Cuando una de estas células capta un patógeno, lo procesa y expone sus ag lo que lleva a la célula a una condición madura. En este nuevo estado el patrón molecular de membrana cambia absolutamente. Esto varía el direccionamiento que esta célula tiene, y empieza a migrar hacia el ganglio. Cuando entran al linfonodo presentan sus ag en las MHC y esperan la entrada de los linfocitos vírgenes que vienen a través del torrente sanguíneo, produciéndose el encuentro tan ansiado que va a detonar la expansión clonal. Con esta expansión el L se transforma en un L efector, la composición proteica de su membrana cambia y su destino ahora se encuentra en la periferia, donde deberá controlar y combatir a los patógenos.
3.- Funciones de la inmunidad innata en el organismo. Describa características y componentes de la inmunidad innata, comparándolas con la respuesta inmune adaptativa. Explique qué son los "modelos moleculares asociados a los patógenos" ("pathogen asociated molecular patterns" o PAMPs) y los "receptores de modelos moleculares asociados a los patógenos" ("pattern recognition receptors" o PRRs). Señale dónde se encuentran. Indique cual es el objetivo primordial de su acción en el organismo. Dé 2 ejemplos de cómo se obtiene este objetivo.
3.a Funciones de la inmunidad innata en el organismo.
Muy antiguo, estable filogénicamente 1.- primeras líneas de defensa
- Otra gran función de la inmunidad innata, es la participación en la estimulación de las respuestas inmunes adaptativas,
3b Describa características y componentes de la inmunidad innata, comparándolas con la respuesta inmune adaptativa. Puntos de comparación: Especificidad, naturaleza de los patógenos, diversidad de receptores, receptores (afinidad x), distribución de receptores (clonal o no clonal), participación en barreras físicas y/o químicas, proteínas sanguíneas, células participantes, mediadores solubles, tiempo de respuesta, localización acción
3c Explique qué son los "modelos moleculares asociados a los patógenos" ("pathogen asociated molecular patterns" o PAMPs) y los "receptores de modelos moleculares asociados a los patógenos" ("pattern recognition receptors" o PRRs). Señale dónde se encuentran. Indique
cuál es el objetivo primordial de su acción en el organismo. Dé 2 ejemplos de cómo se obtiene este objetivo. (Para contestar esta última pregunta, integrar las acciones de los PRRs con los PAMPs para describir un mecanismo de reconocimiento). Los PAMPs, que son, filogenia, objetivo, ejemplo ( flagelina del flagelo bacteriano peptidoglicano de bacterias gram-positivas, lipopolisacáridos (LPS o endotoxinas) de bacterias gram-negativas, ARN de doble cadena (algunos virus de animales y plantas tienen genoma dsARN o ARN convertido en huésped a dsARN, ADN no metilado (varias de las islas CpG presentan adosados grupos metilo). Los PRRs Que son, Receptores del huésped que se han seleccionado a través de la evolución para reconocer a los PAMPs., Algunos circulan en sangre y linfa (MBL, Mannose binding lectin, proteína sintetizada por el higado).
Tipos de receptores:
- Receptores Scavenger , moléculas que se unen a proteínas de baja densidad.(LDL) que ya no puede unirse con el receptor LDL clásico. Lectinas se encuentran anclados a las membranas plasmáticas de macrófagos y monocitos del huésped. Presenta 3 dominios una lectina simple: extracelular (reconoce carbohidratos), dominio transmembrana y otro intracelular (con ITAM (Immunoreceptor Tyrosine-based Activating Motif), que permite la fosoforilación). ■ Receptor de manosa: se expresa en macrófagos y endotelio. Se une a manosa y fucosa de glucolípidos y glucoproteínas del patógeno, capturando sustancias extrañas y además participa del reconocimiento del tráfico celular. ■ Receptor de B- glucano
■ CR
- Toll- Like Receptors (TLR) son homólogos a una proteína de Drosophila llamada Toll, que participa en la polaridad dorso-ventral de la mosca. Presenta un dominio extracelular distinto al humano; un dominio transmembrana y un dominio intracelular, con 80% de homología, lo que indica una similitud en la vía de transducción. La principal función de este recptor, es ser una señal de alarma frente a productos microbianos, de manera de responder con citoquinas en mamíferos. Ejemplos: TLR-2,3,4,5,6,7,8,9.
Cuáles son las etapas de la reacción inflamatoria? Señale los mecanismos inductores de cada etapa. Indique cuales son los resultados de la reacción inflamatoria desde el punto de vista de la defensa orgánica.
Presencia de patogenos, injuria (Por ejemplo necrosis), presencia de cuerpos extraños desencadenan procesos inflamatorios.
Lo macrófagos portan PRRs que se activan frente a antigenos los fagocitan y secretan citoquinas responsables de la inflamación.
- Vasodilatación y aumento de la permeabilidad :
- Expresión de moléculas de adhesión:
- Quimiotaxis : Citoquinas (quimioquinas), componentes del complemento, productos
Ej.: C3a, C4a y C5a, tiene afinida por por receptores de mastocitos, basófilos y neutrófilos, de manera que al ocurrir la unión, se liberan factores promotores de la inflamación, así como aumentar la actividad leucotáxica de neutrófilos.
3. Citólisis medida por la formación del MAC en las superficies celulares.
Ej.: Cada una de las 3 vías del complemento, terminan por activar al Complejo de Ataque a Membrana (explicado anteriormente), que lisa al agente patógeno.
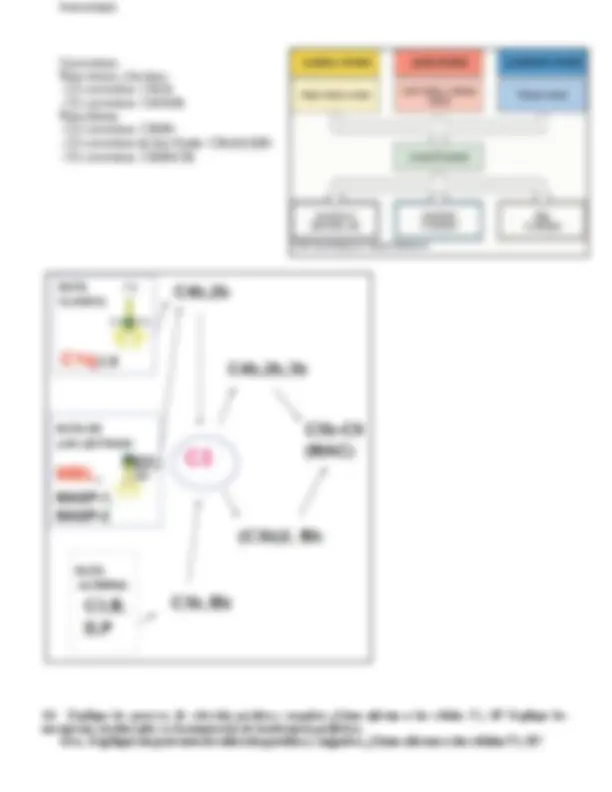
6.- Moléculas y células que actúan como nexos entre la inmunidad innata y adquirida. Diferencias cualitativas y cuantitativas entre la respuesta inmune primaria y secundaria. Describa la respuesta inmune que se generaría en respuesta a una infección viral, mencionando el rol de la respuesta inmune innata y adaptativa. 6A.- Moléculas y células que actúan como nexos entre la inmunidad innata y adquirida.
NEXOS ENTRE RII Y RIA (celular(CD) y molecular(Quimioquinas y citoquinas) Células dendríticas (DC):
- activa linfocitos T naive mediante el MHC con el epitopo y el coestimulador B CD8 mediante MHC I y CD4 mediante MHC II
- permiten la activación de los linfocitos B que reconocen antigenos proteicos, mediante la activación de linfocitos Th1.
- Homing Quimioquinas: Son moléculas proteicas, pequeñas, vida media muy corta, por esta razón tienen efectos rápidos no muy lejos del lugar donde son producidas. Quimioquinas atraviesan el endotelio y se distribuyen generando un gradiente de concentración que va a seguir el leucocito. (homing) Rantes, MIP-1α, MIP-1β: atrae DC inmaduras (CCR1, 5 y 6) y células NK (CCR5) SLC: atrae DC maduras (CCR7) a los vasos linfáticos aferentes. Se expresa en el endotelio de estos vasos IP-10: atrae células T activadas y celulas NK (CXCR3) IL-8: atrae neutrófilos (CXCR ½) *entre paréntesis los receptores de quimioquinas
DC y macrófagos tisulares activados producen Rantes, MIP-1α, MIP-1β, IL-8, IP-10. Th1 vuelve a los tejidos por CCR5 y CXCR3. Th2 vuelve a los tejidos con CCR3, CCR4 y CCR
Citoquinas: Proteínas de bajo peso molecular, pueden actuar de forma autocrina, paracrina y endocrina.
IL-12 e INF-γ: promueve la actividad Th
IL-4, 5 y 13: promueve la actividad Th IL-23, TGF-β, IL-6: promueve actividad Th (presente en enfermedades autoinmunes cronicas) IL-10: citoquina inmunosupresora, promueve anergia clonal (DC tolerogénica)
Complemento: El C3d unido al antígeno potencia la respuesta del linfocito B por crosslink (tiene receptores BcR para el
antigeno y receptores CR2 para el C3d)
Macrófagos vigilan los tejidos del cuerpo y cuando detectan un antígeno por medio de sus PRRs (como los TLRs por ejemplo), lo fagocitan y liberan citoquinas y quimioquinas, estas últimas para reclutar más células, entre ellas, las células dendríticas. Estas últimas al ser vírgenes son altamente fagocíticas, pero cuando capturan al antígeno maduran, ya no fagocitan y pasan a ser APC especializadas, cambiando sus receptores de quimioquinas para ahora poder llegar a los Nódulos Linfáticos regionales por vía linfática aferente. Aquí, presentarán un fragmento de proteína del patógeno en contexto MHC I y II activando (junto con las
DC
inmunogenic a
citoquinas que liberan) a los linfocitos vírgenes o “naive” en T CD8 y T CD4 respectivamente (presentación cruzada). Tanto los macrófagos como las DC al reconocer a un patógeno generan una señal intracelular que activara al factor de transcripción NF-kB para que se expresen genes implicados en la respuesta inmune, incluyendo a las citoquinas, las que inducirán la inflamación o ayudaran en la activación de linfocitos T y B.
6B.- Diferencias cualitativas y cuantitativas entre la respuesta inmune primaria y secundaria.
Respuesta 1ª Respuesta 2ª Tipo de linfocito T y B predominante
Naive Memoria
Tiempo que demora en responder
5-10 días 1-3 días
Tamaño de la respuesta Nº de anticuerpos (peack)
pequeño Grande
Clase de anticuerpos
IgM > IgG IgG IgA IgE Afinidad anticuerpo Baja Alta Inducido por Todos los inmunógenos
Solo proteínas antigénicas Dosis requerida Alta Baja Pendiente de curva de anticuerpos
Achatada Pronunciada (mayor velocidad de producción de anticuerpos) *Ojo que en ambas puede haber RII y RIA
Una respuesta inmune secundaria (2ª exposición al mismo antígeno) es siempre mayor y más rápida que una primaria y esto se debe a que la inmunidad adaptativa presenta memoria inmunológica. Esto significa que cuando el organismo se enfrenta por 2ª vez al mismo antígeno, esta mejor preparado para responder ya que tiene células de memoria, que ya han sufrido cambios de isotipo y maduración de afinidad, por lo que se requerirá una cantidad mucho menor de antígenos para desatar una respuesta de mayor intensidad en menos tiempo. La memoria inmunológica, necesaria para que haya diferencias entre la respuesta inmune 1ª y 2ª, solo existe para antígenos proteicos o T-dependientes. Otra característica de la respuesta inmune 2ª, es la concentración de anticuerpos en el plasma contra antígenos, la cual se mantiene elevada por mucho más tiempo. 6c.- Describa la respuesta inmune que se generaría en respuesta a una infección viral, mencionando el rol de la respuesta inmune innata y adaptativa. El MHC I está presente en todas las células nucleadas (excepto células gonadales y algunas placentarias) El MHC II está presente en DC, macrófagos, linfocitos B, células endoteliales y epitelio tímico.
Cuando las células fagocitan algo (p. ej. bacterias), lo presentan en contexto MHC II; y cuando son antígenos endógenos (creados en la propia célula, como los virus) los presentan en MHC I. Las DC pueden presentar lo que fagocitan como MHC I y II.
a. Anticuerpo
Esta molécula puede ser de superficie o secretada sólo en las cél. Linf. B , por lo tanto la de superficie será BCR (receptor de cél. B) y la secretada serán los anticuerpos o Ig liberados, la diferencia entre estas ésta en que el de superficie tiene 2 dom. Transmembrana y 2 colitas citoplasmática, y la secretada no; esta colita no permite llevar a cabo la activación de mecanismos transductores intracelulares, por tanto requiere del acoplamiento de un molecula que fosforile Ig-alfa e Ig-beta. Y esto forma el BCR (Ig de superficie + estas dos mol. Señalizadoras). En la secretada la mólecula que determina su función efectora es Fc( fragmento cristalizado). Y la región de reconocimiento al antígeno se llama Fab que abarca el dom V + Ch-1 + CL.
b. TCR (receptor de la cél T):
Es muy similar a la Ig. Esta molécula reconoce en un contexto de compl. mayor de histocompatibilidad, al antígeno, Está formado por dominios del tipo de Ig, de 110 aminoácidos unidos por puentes disulfuro, formado por 2 cadenas, también tenemos una región variable y una constante. El reconocimiento está dado por las dos porciones variables. Tiene una cadena αβ y la otra posibilidad es γδ, de esta manera tenemos linfocitos T αβ y linfocitos T γδ. La gran mayoría de los linfocitos T circulantes son αβ y en la periferia, mucosas e intestinos, tenemos γδ. tiene receptores diferentes. Linfocitos αβ, los receptores tienen la cadena β que se comporta como la región variable de la cadena pesada de las Igs,. La cadena α se comporta como la región variable de la cadena liviana. El receptor de linfocito T es similar a la región Fab de la Ig, por lo tanto sólo tiene un dominio constante y la parte genética es más fácil. No hay cambios de clase como en las Ig y los de la cadena β incluso están en otro cromosoma, son genes totalmente diferentes α y β. Requieren al igual que las Ig de moléculas que se acoplen y sean capaz activar una respuesta intracelular, estas son la CD3.
7.2) Comparar la estructura y funcionalidad de las clases más importantes de anticuerpos y sus propiedades efectoras:
En cuanto a la estructura lo que más varía es en relación al número de dominios que tiene la molécula: La Ig G tiene 3 dominios, la región bisagra varía mucho entre ellas. Ig M tiene 4 dominios, adopta a veces estructura pentamérica o hexamérica Circula unida a otras cuatro moléculas por una cadena J. IgD fundamentalemente de mb. muy baja en plasma Ig E tiene 4 dominios IgA: es dimérica , tiene 3 dom. En la región cte y está unida entre ella por una cadena j y además tiene estructura que la protege de la degradación proteolítica, y está estabilizada por una proteína secretora, puede ser monomérica no en secreción.
En cuanto a sus propiedades efectoras:
Ig G: se encuentra en gran concentración en el suero; puede atravesar la placenta, por lo tanto importante en la inmunidad fetal; muy abundante en la respuesta inmune secundaria ; participa en funciones efectoras en el sistema de complemento, de activación de citotoxicidad dependiente de anticuerpo. IgM: Importante en la respuesta inmune primaria. IgA: Está presente en las secreciones mucosas; importante para la inhibición de virus, importante como primera barrera del antígeno en una respuesta inmune secundaria.
Ig E: participa en los procesos alérgicos, monta respuesta segura frente a los parásitos que entran en el cuerpo.
7.3) describa características de Ig útiles en una variedad de ensayos serológicos, inmunoterapia, y diagnóstico: (no cachaba mucho acerca de estos métodos y no lo encontré en las clases así es q apliqué google, sorry, si alguien cacha más de estas técnicas que ayude )
a)En ensayos serológicos encontré Ig G -la seroconversión comienza con una respuesta de la inmunoglobulina M (IgM) contra unas proteínas, con un cambio de los anticuerpos de tipo IgM por los IgG , sólo encontraba estudios basados en estas 2 Ig y se deben usar por su función en el desarrollo de la respuesta inmune primaria tanto como en la secundaria. b )-La Inmunoterapia (IT) se define como la administración gradualmente ascendente de un preparado alergénico hasta lograr la dosis que permita al paciente una mayor exposición al alérgeno identificado sin que se presenten síntomas alérgicos .uno de los mecanismos usados son:
- Disminución a largo plazo de niveles de Ig E específica (por su efecto en el desarrollo de la reacción alérgica) también junto a la Ig G4.
C- Diagnósticos :
Ig A antitransglutaminasa para el diagnóstico de la enfermedad celíaca (cuando es autoinmuno sd de mala absorción) IgE en procesos alérgicos, asmáticos, etc
7.4) ¿Qué son anticuerpos monoclonares, quiméricos, humanizados, y completamente humanos?. Discuta las ventajas comparativas entre ellos en función de su uso.
I)Anticuerpos monoclonares : es un anticuerpo homogéneo producido por una célula híbrida producto de la fusión de un clon de linfocitos B descendiente de una sola y única célula madre y una célula plasmática tumoral. ( Ojo son de células de ratón) Los anticuerpos monoclonales, son anticuerpos idénticos porque son producidos por un solo tipo de célula del sistema inmune, es decir, todos los clones proceden de una sola célula madre. Es posible producir anticuerpos monoclonales que se unan específicamente con cualquier molécula con carácter antigénico.
II) anticuerpos quiméricos: segunda generación de anticuerpos monoclonales, basada en la humanización de los anticuerpos monoclonales de ratón mediante ingeniería genética, evitando así el rechazo del sistema inmune al ser introducidos en el organismo. Son los llamados anticuerpos quiméricos. Un anticuerpo quimérico es creado de tal manera que incorpora parte animal y parte humana. La parte animal o hipervariable (un 30%) es indispensable para que el anticuerpo reconozca la sustancia extraña (antígeno) y la parte humana (un 70%) es responsable de que el sistema inmunológico pueda contribuir a añadir efectividad a su acción. III) Anticuerpo monoclonal humanizado : significa que contiene un 90% de material humano, lo que reduce la inmunogenicidad de los anticuerpos, es decir, el rechazo del sistema inmunológico. La humanización es una técnica que se basa en la estructura terciaria del sitio de combinación con el antígeno, el paratopo , donde existen unas regiones responsables de la unión al antígeno mientras que otras zonas sólo sirven de soporte estructural al paratopo. Por lo tanto las regiones estructurales se obtienen de un anticuerpo humano mientras que las regiones responsables de la unión al antígeno proceden del anticuerpo del ratón. IV)Anticuerpos monoclonares completamente humanos : recién se están empezando a producir por técnicas de ADN recombinante anticuerpos monoclonales completamente humanos, que resuelven en buena parte no sólo el problema
En cuanto a la diversidad genética de MHC , están todos en el mismo cromosoma(6 en humano), en el mismo gen, salvo la beta-2 microglobulina. Se organizan de esta forma, primero, los de clase II, luego clase III y después clase I.
Debido a que heredamos un haplotipo materno y otro paterno, tenemos 6 posibilidades de moléculas de clase I, ya que el MHC clase I codifica sólo cadenas F 06 1F 02 0F 02 83 maternas, 3 paternas) F 02 E Clase II codifica para F 06 1F 02 0 y F 02 0F 06 2F 02 C aquí las combinaciones aumentan. Los libros dicen que existen alrededor
de 20 combinaciones distintas para estas cadenas F 06 1F 02 0y^ F 02 0F 06 2F 02 C pero la profe sólo ha identificado 12. Así que existen por lo menos 12 combinaciones distintas.
Las moléculas clásicas (HLA A, HLA B, HLA C, HLA DP, HLA DQ, HLA DR) son muy polimórficas, se podría decir que nos hace únicos; mientras que las no clásicas, no lo son tanto. Debido a este gran polimorfismo existen tantos problemas en el transplante de órganos. Es casi imposible que existan dos personas con el mismo HLA.
En cuanto a la variabilidad de TCR : -Hay loci separados que codifican la cadena del TCR, las cadenas , del TCR y la cadena del TCR. Todos estos loci contienen segmentos V y J, y sólo los loci de cadenas y del TCR, poseen segmentos génicos D. Las recombinaciones somáticas de los loci de TCR implican la unión de segmentos D y J, seguido de la unión de un segmento V a los segmentos DJ recombinados. Este proceso está mediado por un complejo enzimático recombinasa que incluye los componentes linfocito específicos RAG1 y RAG2. La diversidad de los repertorios TCRs se genera por las asociaciones recombinacionales de múltiples genes V, D y J de la línea germinal y la diversidad de unión generada por la adición de nucleótidos al azar en los sitios de recombinación. Estos mecanismos generan la mayor diversidad en la unión de las regiones V y C que forman las terceras regiones hipervariables de las cadenas polipeptídicas TCRs. Receptor de celula T
c. ¿es posible que un linfocito B mantenga la especificidad por un epitopo y cambie su función efectora?
- Si ES POSIBLE dado esto se logra a través del Cambio de Clase o Switching Isotípico En linfocito B naive, será IgM masivamente y en menor medida IgD. Si el linfocito ha sido expuesto al antígeno (ya es un linfocito efector), lo que hará molecularmente es la producción de IgM secretora, transcribiendo hasta la señal de término TT0. La respuesta inmune no es 1:1, sino que un linfocito que ha encontrado un antígeno genera una respuesta clonal. Entonces los hijos de ésta son los que atacan, son células efectoras: producen IgM secretora. Otras células sin embargo, van a tomar el camino de las células de memoria. Esta memoria está asociada, por lo general, al cambio de la cadena constante para hacer IgG generalmente.
Cuando llega el antígeno, el BCR cambia de IgM a cualquier otra Ig. Los exones río abajo tienen una señal llamada señal S (representada por un círculo negro en el diagrama; éstas llevan el nombre del segmento al que anteceden: Sγ 3 , Sγ 1 , S (^) α 1 , S (^) γ 2 , etc.), que viene de switching , cambio. Esta secuencia de DNA es muy repetitiva de aproximadamente 5 Kpb. Estas secuencias son complementarias con la del círculo lila, denominada S (^) μ. Sμ es más amplia, por lo que tiene distintos sitios de afinidad para cada uno del resto de las
señales S. En la progenie que se convertirá en células de memoria, S μ se apareará con la secuencia Sγ 1 , para formar IgG-1 por ejemplo, y dejará al exón γ1 contiguo al exón μ para que el promotor tome todo esto y desaparezca μ. Esto está dado por las citoquinas.
Hoy se sabe que el linfocito T ayuda al B. El linfocito T empieza a secretar citoquinas y, dependiendo de la naturaleza de éstas, se decidirá cuál es el apareo que genera esta señal. Por ejemplo, en presencia de altos niveles de IFN-γ, se produce la unión de Sμ a S (^) γ 1 , dando paso a la producción de γ1 en el citoplasma, que se unirá a cadenas livianas verdaderas, que será la misma de la célula original. Sólo se hace un cambio del acompañamiento de las cadenas livianas. Esta célula ya no tendrá IgM como BCR, sino sólo γ1. Pero algunas de estas células liberan anticuerpos a pesar de estar en la línea de las células de memoria, produciendo IgG-1. Hay otros cambios de clase dependiendo del contexto: IgA, por ejemplo, abunda en respuestas contra virus. En una infección vírica, entonces, habrá tanto IgA como IgG.
IgG
Consecuencias para la RIA de la interacción PAMP-PRR.
- Esta interacción genera moléculas coestimuladoras en las APC (B7.1 y B7.2 o CD80 Y CD86)
- (^) Producción de señales patógeno-específica: 1º Señal: interacción MHC-péptido-TCR 2º Señal: CD28-B
- Si la célula blanco es una célula somática (todas excepto GR) las CD8+ las matarán ( gran parte de la sintomatología de una enfermedad viral es por la destrucción de células propias que contienen al no propio )
- Si la célula blanco es una APC, CD40L del LT la activará para que exprese mayores niveles de B7.
- En ausencia de la señal mediada por CD28, el LT se hace tolerante o anérgico, quizás por elegir involucrar a CTLA4 en su interacción con la APC, o sea, hay un cambio de ligando de CD28 a CTLA4.
Moléculas Accesorias. (esto no lo preguntan pero no está demás saberlo, yo me lo saltaria)
“Los linfocitos T requieren de moléculas accesorias que ayuden al linfocito T a transducir la información a la membrana.” Hay un TCR por cada 2 CD3 (2 ε, δ, γ) y lo poseen todos los linfocitos T, tanto los CD4+ o CD8+. En la intermembrana hay distribución de cargas diferentes, ya sea que el TCR estará cargado + y las moléculas ayudadoras -, entonces entre ambas se atraen, e interaccionan, esta unión ocurre cuando la APC se une al LT. Existen ayudadoras ζ (z) que está entre el TCR. El CD3 y z tienen dominios ITAM (motivación de activación de tirosina) y comienzan a haber fosforilaciones. Por cada TCR hay 10 sitios ITAM. Cuando se une el TCR al MHC II (en este caso) el CD4+ a través de su región bisagra irá a ponerse en contacto con el segmento de MHC II que está pegado a la membrana de la APC, esto porque no hay otra forma de unir algo que está tan lejos. Cuando esto sucede comienza la fosforilación de los residuos de tirosina y vendrán moléculas adaptadoras Lck (quinasa) que adaptará la fosforilación y reclutará a Zap-70, la cual se unirá a parte de las cadenas z, mediante sus dominios de alta afinidad con tirosina fosforilada (dominio SH2) y el otro dominio (SH3) tiene afinidad por prolina y esta será un ancla que conectará con otro dominio río abajo. La presentación necesita a las moléculas coestimuladoras, si éstas no están pueden caer en estado de anergia que implica estar activado pero no responder.
Sinapsis Inmunológica. (repetición de lo de arriba no lo saco por si le intereza a alguien)
La interacción entre un Linfocito T y su APC es muy estrecha y está compuesta de la interacción entre MHC-TCR , de la coestimulación B7-CD28 y moléculas de adhesión como ICAM1-LFA1 , que generan un complejo de unión que tiene dos zonas definidas, la periférica donde están las moléculas de adhesión y se
llama B-Mac y la central donde están MHC-TCR. Esta sinapsis inmunológica dura casi una hora en la cual se producen todas las señales conocidas. Las conexinas estarían participando en la sinapsis. Cuando hay activación del linfocito T aumenta el Ca++^ citosólico lo que es importante y se puede medir para saber si un linfocito T ha sido activado o no activado, ya que el Ca ++^ también genera fosforilaciones. Lo importante es que todas las vías convergen a la activación genética, muchas veces se necesita que
todas las vías se unan para activar un gen. Funcionamiento del CTLA4: Puede competir por el B7 y si se une inactiva a la célula o si se une paralelamente con CD28, generará factores que inhiben las señales que generan las cascadas para activar genes. Cuando estos receptores (TCR) y coestimuladores (CD28) son utilizados permite que se unan a él unas ligasas que permiten la unión de ubiquitina que es una señal proteolítica.
9. B Colaboración de los linfocitos T “helper” a otras células del sistema inmune. (MENCIONE DETALLADAMENTE AL MENOS TRES ACTIVIDADES DE AYUDA O COLABORACION QUE LOS LINFOCITOS T HELPER A OTRAS CELUAS DEL SISTEMA INMUNE)
Cuando los patógenos están en vesículas de macrófagos, serán presentados por MHC II y generarán linfocitos Th1. La respuesta inmune humoral se da cuando el lugar de ubicación son los fluidos extracelulares presentado por MHC II y genera Th2 (principalmente) y Th1 que activará linfocitos B.
Ambos linfocitos Th son ayudadores (ambos son CD4+) pero en el contexto de diferentes patógenos:
- Linfocito Th1 : patógenos en vesículas del macrófago, esta ayuda hace que la bacteria se destruya dentro del macrófago.
- Linfocito Th2: un linfocito B es reconocido y lo activa para que libere anticuerpos. Los linfocitos T CD8+ (citotóxico) que utilizan para activarse Th1 induce la destrucción de la célula que contiene al patógeno.
Detalle de lo anterior (la respuesta queda contestada de lo anterior, pero este es el detalle)
Respuesta inmune celular mediada por Th1.
Cuando el CD4+ interactúa con CD o macrófago cobra vital importancia CD40-CD40L. Con esto habrá estimulación de estas células y producción de IL-12 que puede:
- Estimular CD4+ naives a diferenciarse a Th1.
- Estimular NK para que produzcan INFγ que actuará en el receptor de INFγ en la CD o macrófago para que se produzca IL-12 y en el macrófago también estimula la actividad patogénica.
Fase efectora de Th1:
- Interacción con el macrófago con MHCII-TCR, CD40-CD40L y el Th1 producirá IFNγ que activará al macrófago pero también sirve para mantener el fenotipo de Th1.
- Sin inflamación no son capaces de migrar
- Su función principal es transportar al patógeno.
Las APC más especializadas son las células dendríticas. Tienen receptores de membrana que las transportan desde la periferia al ganglio linfático, pueden hacer presentación cruzada:
Cuando las células fagocitan algo, lo presentan en MHCII. Cuando son endógenos (creados en la propia célula, como los virus) lo presentan en MHCI. Sin embargo, las CD pueden presentar algo que fagocitaron en MHCI (presentación cruzada). Las CD están en todas partes pero en bajas cantidades, siempre va a una zona de peligro y acá puede estar inmadura o madura.
- Inmaduras: acción fagocítica
- Maduras: fenotipo distinto con muchas prolongaciones y gran cantidad de MHC.
Son muy difíciles de conseguir de la sangre porque son sólo el 1%. Existen métodos para producirlas a partir de daño periférico, entonces a partir de monocitos de una fracción de la sangre se pueden utilizar factores de crecimiento que permitan la diferenciación a CD que primero serán inmaduras. Estas células presentan antígeno, pero no son atacadas por los linfocitos, porque desde la periferia las CD migran al ganglio, razón por la cual no se puede encontrar con los linfocitos en la periferia. La diferencia de las CD inmaduras y maduras es a parte de la forma y la función, los marcadores de membrana que se pueden medir por densitometría de flujo con Ac monoclonales contra Ag específicos que emiten fluorescencia, esto se mide para caracterizarlo. Las CD se comunican entre sí por gap junctions (conexina 43). Para comprobar si se pasaban información la una a la otra se teñía una célula con cierto colorante y se le microinyectaba colorante de otro color y se veía que este se expresaba en su célula vecina. Se ha descrito que se puede traspasar Ag, así una CD periférica puede viajar al ganglio y traspasarle el Ag a una célula que reside allí.
Macrófagos (fagocitos mononucleares).
- Reconocen PAMPs a través de PRR (gran cantidad)
- Función es reconocer al patógeno en el momento que ingresa al organismo
- Fagocitan al patógeno para eliminarlo
- Al activarse secretan una serie de sustancias (citoquinas) que sirven para atraer más refuerzos contra el patógeno.
Los macrófagos que han ingerido microorganismos exhiben antígenos microbianos a los linfocitos T efectores diferenciados. A continuación, estas células activan a los macrófagos para destruir los microorganismos. Este proceso es un mecanismo muy importante de la inmunidad celular contra los microorganismos intracelulares. Los que han captado microorganismos también son importantes en la activación de linfocitos T vírgenes para desencadenar las respuestas primarias contra los antígenos microbianos. También tienen funciones efectoras en la inmunidad innata y adaptativa. En la innata consiste en fagocitar microorganismos y sintetizar citoquinas que reclutan y activan otras células inflamatorias. En la adaptativa, los linfocitos T estimulados activan los macrófagos para destruir los microorganismos fagocitados. En la humoral, los anticuerpos recubren u opsonizan los microorganismos y estimulan su fagocitosis mediante receptores de superficie para los anticuerpos presentes en los macrófagos. Secretan 4 citoquinas principalmente que tienen importancia en su acción sobre el endotelio vascular:
- TNF
- (^) IL-
- IL-
- Quimiioquinas Células B. Los linfocitos B actúan como APC para los linfocitos Th durante la respuesta inmune humoral. Son las células productoras de anticuerpos. En el ganglio, en el folículo linfoide hay LB reaccionando con un Ag,
paralelamente está el LT reaccionando con el Ag mediante APC. Luego de esto el LB endocita el Ag, viaja a la zona T, al unirse el LB al LT podrá proliferar generando células efectoras y células de memoria. El linfocito B al unirse con el LT nunca sale del folículo. Los MHC nunca están solos en la superficie porque son inestables, entonces siempre tiene que estar con algún péptido y como no siempre hay Ag, la mayoría de las veces está presentando péptidos propios. El LB se une al Ag, lo endocita, luego une esta vesícula a fagosomas y lo degrada para unir los péptidos a MHC-II que se pondrán en la membrana para unirse a LT- CD4+. Cuando un Ag se une por primera vez a una célula, las posteriores uniones ya sean a células efectoras, a LT-CD4+ serán siempre con el mismo Ag. El LB con el LT se unen mediante el TCR-péptido-MHCII, más la unión del CD40-CD40L y la interacción del receptor de citoquinas con IL-2, con esto el LB queda listo para comenzar a proliferar. El LT al producir IL-2 da señal para que prolifere el LB. El cambio de clase va a estar mediado por la activación del LT por una determinada CD.
10 b.- Comente, con la mayor información posible, acerca de las vías de procesamiento y presentación antigénica de clase I y II.
MHC-I.
- Formado por cadena α con 3 dominios (1,2,3) más la cadena β-microglobulina que da la estabilidad.
- Bolsillo formado por α1 y α2 (formado por cadenas β sheet) y sus aniones son por interacciones con péptidos del antígeno (9-11aá)
- Se expresa en todas las células nucleadas (excepto las gonadales y algunas placentarias)
- Cadena β no es polimórfica
- Presenta péptidos codificados endógenamente (propias, virales, bacterias intracelulares). Los patógenos endógenos están libres en el citosol.
MHC-II.
- 2 cadenas α-β ambas polimórficas
- Bolsillo α1-β1 (más abierto) permite que sean péptidos más grandes (17-30aá)
- Como ambas cadenas son polimórficas podemos tener mínimo 6 tipos y máx 12.
- Se expresa en APC (dendríticas, macrófagos, LB)
- Presentan péptidos exógenos (patógenos extracelulares)que pueden encontrarse dentro de la célula pero en vesículas.
La presentación necesita a las moléculas coestimuladoras, si estas no están pueden caer en estado de anergia que implica estar activado pero no responder. PRIMING: es la sensibilización de un linfocito maduro no activado, requiere gran cantidad de antígenos en contexto de MHC más coestimuladoras.