Baixe Capítulo Imuno Básica e outras Notas de estudo em PDF para Biotecnologia, somente na Docsity!
| 19
CapÌtulo 1
ImunologiaImunologiaImunologiaImunologiaImunologia
AntÙnio Teva JosÈ Carlos Couto Fernandez Valmir Laurentino Silva
- IntroduÁ„o ‡ Imunologia1. IntroduÁ„o ‡ Imunologia1. IntroduÁ„o ‡ Imunologia 1. IntroduÁ„o ‡ Imunologia1. IntroduÁ„o ‡ Imunologia
A imunologia È uma ciÍncia recente. Sua origem È atribuÌda, por alguns autores, a Edward Jenner, que, em 1796, verificou proteÁ„o induzida pelo cowpox (vÌrus da varÌola bovina) contra a varÌola humana, nomeando tal pro- cesso da vacinaÁ„o. No entanto, È sabido que, na antiguidade, os chineses j· inalavam o pÛ das crostas secas das p˙stulas de varÌola ou as inseriam em pequenos cortes na pele, em busca de proteÁ„o.
O sistema imune È o conjunto de cÈlulas, tecidos, Ûrg„os e molÈculas que os humanos e outros seres vivos usam para a eliminaÁ„o de agentes ou molÈculas estranhas, inclusive o c‚ncer, com a finalidade de se manter a homeostasia do organismo. Os mecanismos fisiolÛgicos do sistema imune con- sistem numa resposta coordenada dessas cÈlulas e molÈculas diante dos orga- nismos infecciosos e dos demais ativadores, o que leva ao aparecimento de respostas especÌficas e seletivas, inclusive com memÛria imunit·ria, que tambÈm
20 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
pode ser criada artificialmente, atravÈs das vacinas. Na ausÍncia de um sistema imune funcional, infecÁıes leves podem sobrepujar o hospedeiro e lev·-lo ‡ morte. PorÈm, mesmo com um sistema imune funcional, o homem, por exem- plo, pode adquirir uma doenÁa infecciosa ou um c‚ncer, pois a resposta imune especÌfica, diante de um agente agressor, leva tempo para se desenvolver e, alÈm disso, tanto organismos estranhos, como cÈlulas neopl·sicas, desenvol- vem mecanismos de evas„o para fugir da resposta imune.
Neste capÌtulo, ser„o abordados conceitos b·sicos dos principais com- ponentes do sistema imune, os mecanismos de resposta especÌfica ante os diversos agentes infectoparasit·rios, como tambÈm a investigaÁ„o dos vestÌgios da passagem desses agentes, por meio de mÈtodos laboratoriais para pesquisa de antÌgenos e anticorpos especÌficos, principal propÛsito desse texto, uma vez que se destina a alunos de escolas tÈcnicas de nÌvel mÈdio.
- ”rg„os, tecidos e cÈlulas2. ”rg„os, tecidos e cÈlulas2. ”rg„os, tecidos e cÈlulas 2. ”rg„os, tecidos e cÈlulas2. ”rg„os, tecidos e cÈlulas envolvidos na resposta imunit·riaenvolvidos na resposta imunit·riaenvolvidos na resposta imunit·riaenvolvidos na resposta imunit·riaenvolvidos na resposta imunit·ria
2.1. CÈlulas que participam do sistema imunit·rio
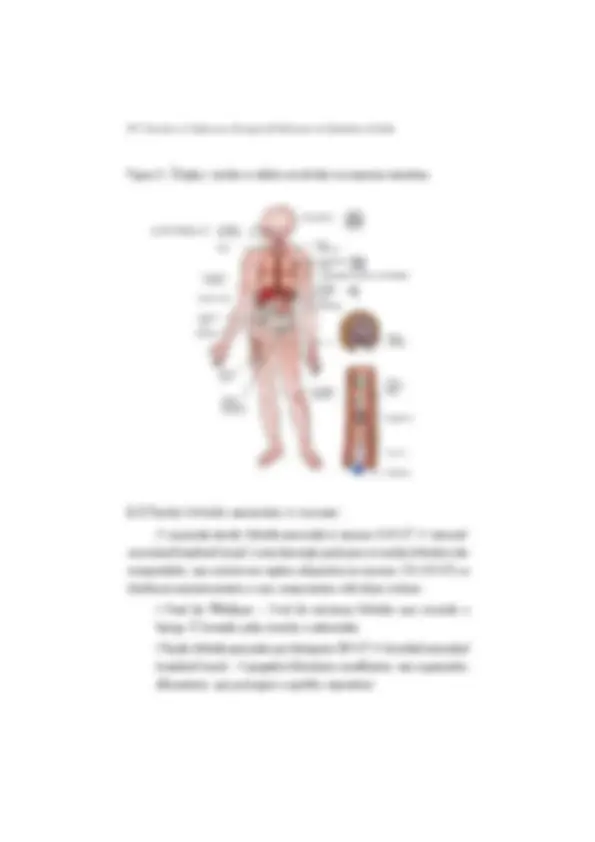
As respostas imunes s„o mediadas por uma variedade de cÈlulas e por molÈculas que estas cÈlulas expressam (Figura 1). Os leucÛcitos s„o as cÈlulas que desempenham as principais aÁıes, mas outras cÈlulas, que se encontram nos tecidos, tambÈm participam da resposta imunit·ria, enviando sinais e rece- bendo estÌmulos dos leucÛcitos. As cÈlulas que participam do sistema imunit·rio se originam na medula Ûssea, onde muitas evoluem para a fase adulta. A partir da medula, e por meio de vasos sanguÌneos, elas migram junto com todos os elementos celulares do sangue. Inclusive as hem·cias, que transportam o oxigÍ- nio, e as plaquetas que participam da coagulaÁ„o, uma vez que estes elemen- tos se originam das cÈlulas-tronco progenitoras da medula. As cÈlulas que derivam do progenitor mieloide e do progenitor linfoide s„o as que mais
22 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
Os eosinÛfilos parecem ser importantes, principalmente na resposta
diante de infecÁıes parasit·rias ou processos alÈrgicos, j· que seu n˙mero
aumenta no curso destas reaÁıes.
A funÁ„o dos basÛfilos provavelmente È similar e complementar ‡ dos
eosinÛfilos e mastÛcitos.
Os mastÛcitos, cujo precursor parece ser comum aos basÛfilos,
devido a semelhanÁas funcionais, tambÈm se diferenciam ao chegar aos
tecidos onde residem. Eles se localizam principalmente ‡ margem dos
vasos sanguÌneos e liberam mediadores que agem nas paredes vasculares
quando ativados.
Figura 1. CÈlulas que participam do sistema inunit·rio
| 23
O progenitor linfoide comum d· origem aos linfÛcitos. Os linfÛcitos s„o as cÈlulas que reconhecem, especificamente, os antÌgenos. Sua morfologia tÌpica consiste em uma pequena cÈlula redonda com n˙cleo esfÈrico. Apesar da aparÍn- cia uniforme ‡ microscopia Ûtica, v·rios tipos de linfÛcitos podem ser distinguidos com base nas suas propriedades funcionais e proteÌnas especÌficas que expressam. A distinÁ„o mais fundamental consiste na classificaÁ„o destas cÈlulas em duas linhagens principais, conhecidas como linfÛcitos B e linfÛcitos T.
Os linfÛcitos B, tambÈm chamados de cÈlulas B (de bursa ou bolsa de Fabricius, nas aves, e derivadas da medula Ûssea, nos mamÌferos), quando ativados, proliferam e se diferenciam em cÈlulas plasm·ticas ou plasmÛcitos, que s„o as cÈlulas efetoras da linhagem B, cuja funÁ„o principal È a secreÁ„o de anticorpos. Os linfÛcitos T, ou cÈlulas T (derivados do timo), se apresentam em duas classes principais. Uma se diferencia, quando ativada, em cÈlulas T CD8+ ou citotÛxicas, que matam as cÈlulas infectadas, ao passo que a outra classe de cÈlulas T, chamadas de cÈlulas T CD4+ ou auxiliares, atuam na ativaÁ„o de outras cÈlulas, como os linfÛcitos B e os macrÛfagos, alÈm de coordenar a resposta imunit·ria.
O receptor de antÌgeno da cÈlula B (BCR) (Figura 2) È uma forma de anticorpo ligada ‡ membrana que a cÈlula B passa a produzir, apÛs sua ativaÁ„o e diferenciaÁ„o em cÈlula plasm·tica. Os anticorpos s„o molÈculas agrupadas em uma classe de subst‚ncias denominadas imunoglobulinas, e o receptor de antÌgeno do linfÛcito B È tambÈm conhecido como imunoglobulina de mem- brana. A imunidade humoral È a principal funÁ„o das cÈlulas B e dos plasmÛcitos, e consiste em secretar anticorpos no sangue e em outros lÌquidos org‚nicos, resultando efeitos protetores, mediados por lÌquidos teciduais.
O receptor de antÌgeno da cÈlula T (TCR) (Figura 2) constitui uma classe heterogÍnea de proteÌnas de membrana que, embora estejam relaciona- das evolutivamente com as imunoglobulinas, s„o diferentes delas, j· que est„o
Imunologia
| 25
Algumas das cÈlulas-filhas retomam ao estado de repouso, tornando-se cÈlu- las de memÛria, que podem sobreviver por v·rios anos. Estes linfÛcitos de memÛria representam uma grande proporÁ„o das cÈlulas do sistema imunit·rio. A outra progÍnie do linfÛcito virgem ativado diferencia-se em cÈlulas efetoras, que sobrevivem apenas alguns dias, mas que, durante este perÌodo, executam atividade que resultam em defesa.
Outra classe de cÈlulas linfoides, chamada de cÈlulas matadoras natu- rais ou cÈlulas natural killer (NK), È desprovida de receptores antÌgeno- especÌficos, sendo parte do sistema imune inato. Essas cÈlulas circulam no sangue como grandes linfÛcitos, com diferentes gr‚nulos citotÛxicos, e s„o capazes de reconhecer e matar algumas cÈlulas anormais, tais como cÈlulas tumorais e cÈlulas infectadas por vÌrus. E parecem ser importantes na defesa contra biopatÛgenos intracelulares na imunidade inata.
2.2. Os Ûrg„os linfoides e a rede linf·tica
Os Ûrg„os linfoides (Figura 3) s„o tecidos organizados que contÍm grandes quantidades de linfÛcitos em um ambiente de cÈlulas n„o linfoides. Nesses Ûrg„os, as interaÁıes que os linfÛcitos tÍm com as cÈlulas n„o linfoides s„o importantes, tanto para o desenvolvimento dos linfÛcitos e o inÌcio da resposta imune adaptativa, como para a manutenÁ„o dos mesmos. Tais Ûr- g„os podem ser divididos em Ûrg„os linfoides centrais ou prim·rios, produ- tores de linfÛcitos, e Ûrg„os linfoides perifÈricos ou secund·rios, que de- sempenham a funÁ„o de maximizar o encontro entre os linfÛcitos e os produtos processados pelas cÈlulas apresentadoras de antÌgenos, dando inÌ- cio ‡ resposta imune. Os Ûrg„os linfoides centrais s„o a medula Ûssea vermelha e o timo, um grande Ûrg„o localizado na porÁ„o superior do tÛrax. Tanto os linfÛcitos B como as cÈlulas T surgem na medula Ûssea, mas apenas os linfÛcitos B ali se diferenciam. Os linfÛcitos T migram para o timo para sofrer seu processo de diferenciaÁ„o. Uma vez completada sua maturaÁ„o
Imunologia
26 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
celular, os dois tipos de linfÛcitos entram na corrente sanguÌnea, migrando para os Ûrg„os linfoides perifÈricos. Durante a vida intrauterina, o fÌgado fetal desempenha o papel que a medula Ûssea vermelha passa a desenvol- ver plenamente apÛs o nascimento.
Os Ûrg„os linfoides perifÈricos s„o especializados na captura do antÌgeno para possibilitar o inÌcio das respostas imunes adaptativas. Os microrganismos patogÍnicos podem penetrar no hospedeiro por muitas portas de entrada, instalando o processo infeccioso em qualquer sÌtio, mas o encontro do antÌgeno com os linfÛcitos acontecer· nos Ûrg„os linfoides perifÈricos: os nÛdulos linf·ticos, o baÁo e v·rios tecidos linfoides associa- dos ‡s superfÌcies das mucosas. Os linfÛcitos est„o em contÌnua recirculaÁ„o entre esses tecidos, para os quais o antÌgeno tambÈm È carreado, vindo de todos os locais de infecÁ„o, primariamente dentro de macrÛfagos e cÈlulas dendrÌticas. Dentro dos Ûrg„os linfoides, cÈlulas especializadas, como as cÈlulas dendrÌticas maduras, apresentam o antÌgeno para os linfÛcitos.
A rede linf·tica consiste em um extenso sistema de vasos que coletam o lÌquido intersticial, fazendo-o retornar para o sangue. Esse lÌquido intersticial È produzido continuamente pela passagem de ·gua e solutos de baixo peso molecular atravÈs das paredes vasculares que pe- netram no espaÁo intersticial, pela secreÁ„o celular e outros fatores de excreÁ„o. Ao ser parcialmente drenado para os vasos linf·ticos, passa a ser chamado de linfa. A linfa flui lentamente pelos vasos linf·ticos prim·- rios, des·gua em vasos linf·ticos de calibre progressivamente maior, que convergem para o ducto tor·cico, e desemboca na veia cava superior, que, por sua vez, devolve todo o volume para a corrente sanguÌnea, num fenÙmeno denominado recirculaÁ„o.
Localizados em pontos de convergÍncia da rede vascular, os nÛdu- los linf·ticos constituem uma sÈrie de Ûrg„os encapsulados em forma de ìcaroÁo de feij„oî, que se distribuem ao longo dos vasos linf·ticos. Os
28 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
Figura 3. ”rg„os, tecidos e cÈlulas envolvidos na resposta imunit·ria.
2.3.Tecido linfoide associado ‡ mucosa
A express„o tecido linfoide associado ‡ mucosa (MALT =mucosal- associated lymphoid tissue) È uma descriÁ„o geral para os tecidos linfoides n„o encapsulados, que existem nas regiıes subjacentes ‡s mucosas. Os MALTs se distribuem anatomicamente e seus componentes individuais incluem:
- Anel de Waldeyer - Anel de estruturas linfoides que circunda a faringe. … formado pelas tonsilas e adenoides.
- Tecido linfoide associado aos brÙnquios (BALT =bronchial-associated lymphoid tissue) - Agregados linfocit·rios semelhantes, mas organizados difusamente, que protegem o epitÈlio respiratÛrio.
| 29
- Tecidos linfoides associados ao intestino (GALT =gut-associated lymphoid tissues) - Incluem folÌculos linfoides isolados e o apÍndice cecal, alÈm de estruturas especializadas do intestino delgado, as placas de Peyer.
- Tecido linf·tico urogenital
- Entre outros MALTs (Figura 3). Coletivamente, estima-se que o sistema imune de mucosa contenha tantos linfÛcitos quanto o resto do corpo. Esses linfÛcitos formam um grupo especial de cÈlulas que seguem leis um tanto diferentes. Embora notavelmente diferentes em sua aparÍncia, os nÛdulos linf·ticos, o baÁo e os tecidos linfoides associados ‡ mucosa demonstram a mesma arquitetura b·sica. Cada um deles opera segundo o mesmo princÌpio, capturando o antÌgeno nos locais de infec- Á„o e apresentando-o a pequenos linfÛcitos migratÛrios para, assim, induzirem as respostas imunes adaptativas. Os tecidos linfoides perifÈricos tambÈm proveem sinais de sobrevivÍncia aos linfÛcitos que n„o encontram seu antÌgeno especÌfi- co. Isto È importante para manter o n˙mero correto de linfÛcitos T e B circulantes, e assegura que somente os linfÛcitos com o potencial de responder ao antÌgeno estranho sejam mantidos.
2.4. RecirculaÁ„o de linfÛcitos
Os pequenos linfÛcitos T e B que se diferenciaram na medula Ûssea e no timo, mas que ainda n„o se encontraram com o antÌgeno, s„o referidos como linfÛcitos virgens ou em repouso. Estes elementos circulam continua- mente do sangue para os tecidos linfoides perifÈricos, nos quais penetram por meio de interaÁıes adesivas especiais com os capilares e retornam para o sangue atravÈs dos vasos linf·ticos ou, no caso do baÁo, diretamente ao sangue. Na presenÁa de uma infecÁ„o, os linfÛcitos que reconhecem o agente infeccioso s„o retidos no tecido linfoide, onde proliferam e se diferenciam em cÈlulas efetoras, capazes de controlar a infecÁ„o.
Imunologia
| 31
cÈlulas T CD4 ou CD8 ocorre na medula tÌmica, e as cÈlulas T maduras s„o liberadas para a circulaÁ„o.
3.1. Receptores de antÌgenos e molÈculas acessÛrias dos linfÛcitos T
Os linfÛcitos T respondem aos antÌgenos peptÌdicos, que s„o expos- tos pelas cÈlulas apresentadoras de antÌgenos (APCs). O inÌcio desta res- posta requer o reconhecimento especÌfico do antÌgeno pelas cÈlulas T, a ades„o est·vel das cÈlulas T ‡s APCs e a transduÁ„o dos sinais ativadores. Cada um desses eventos È mediado por molÈculas distintas, expressas pelas cÈlulas T. As molÈculas de MHC e os peptÌdeos formam um complexo na membrana plasm·tica das APCs. O receptor que reconhece esse complexo peptÌdeo-MHC È o TCR (Figura 2), que È distribuÌdo clonalmente, ou seja, os clones de linfÛcitos que apresentam diferentes especificidades ex- pressam distintos TCRs. Os sinais bioquÌmicos, que s„o acionados na cÈlula T pelo reconhecimento do antÌgeno, n„o s„o transduzidos pelo TCR, mas por proteÌnas n„o vari·veis chamadas CD3 edzeta (z), que est„o ligadas de forma n„o covalente ao receptor do antÌgeno para formar o complexo TCR. Portanto, nas cÈlulas T, o reconhecimento do antÌgeno È basicamente realiza- do por dois grupos de molÈculas: um receptor para o antÌgeno altamente vari·vel, o TCR, e proteÌnas sinalizadoras n„o vari·veis (CD3 e cadeia z). Outras molÈculas acessÛrias funcionam como molÈculas de ades„o para esta- bilizar a ligaÁ„o das cÈlulas T ‡s APCs, permitindo que o TCR mantenha Ìntimo contato com o antÌgeno durante o tempo suficiente para a transduÁ„o dos sinais necess·rios ‡ ativaÁ„o dessas cÈlulas.
As cÈlulas T que expressam o TCR °d pertencem a uma linhagem distinta das cÈlulas T restritas ao MHC. A percentagem das cÈlulas T °d È muito vari·vel nos diferentes tecidos das diferentes espÈcies, normalmente n„o excedendo mais do que 5%. Elas n„o reconhecem os antÌgenos peptÌdeos
Imunologia
32 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
associados ‡s molÈculas MHC e n„o s„o restritas ao MHC. Alguns clones dessas cÈlulas reconhecem uma pequena molÈcula que pode ser apresentada por molÈculas similares ‡s da classe I do MHC, ou seja, uma apresentaÁ„o n„o cl·ssica de molÈculas normalmente encontradas nas microbactÈrias e em outros microrganismos. A diversidade limitada das cÈlulas °d sugere que os ligantes desses receptores s„o bem conservados. Elas podem iniciar a resposta imune contra um pequeno n˙mero de microrganismos antes mesmo do recrutamento das cÈlulas T antÌgeno-especÌficas ab.
AlÈm dos componentes do complexo TCR, as cÈlulas T apresentam v·rias proteÌnas de membrana, as quais exercem papel crucial na resposta destas cÈlulas no reconhecimento do antÌgeno. Essas molÈculas presentes na membrana de linfÛcitos ligam-se especificamente a outras molÈculas da membrana de outras cÈlulas, como as APCs, cÈlulas do endotÈlio de vasos e da matriz extracelular. Essas molÈculas n„o apresentam regiıes vari·veis, n„o s„o polimÛrficas, s„o idÍnticas em todas as cÈlulas T de todos os indivÌduos de uma mesma espÈcie, e s„o respons·veis pela transduÁ„o de sinais bioquÌmicos para o interior das cÈlulas T. Essa propri- edade assegura que as cÈlulas T e as APCs permaneÁam ligadas o tempo suficiente para permitir aos TCRs a oportunidade de localizar, reconhecer e responder ao complexo peptÌdeo-MHC na APC.
3.2. Correceptores CD4 e CD8: Receptores envolvidos na ativaÁ„o
As molÈculas CD4 e CD8 s„o proteÌnas das cÈlulas T que se ligam ‡s regiıes n„o polimÛrficas das molÈculas de MHC e transduzem os sinais que, juntamente com os sinais liberados pelo complexo TCR, iniciam a ativaÁ„o das cÈlulas T. Normalmente, as cÈlulas T ab maduras expressam CD4 ou CD8, embora existam referÍncias da express„o de ambos os marcadores. Esses correceptores interagem com as molÈculas de MHC, quando o TCR reconhe-
34 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
4.2. RelaÁ„o filogenÈtica dos antÌgenos
A estimulaÁ„o de linfÛcitos de galinhas com proteÌna de pato resulta em uma resposta imune muito baixa. Por outro lado, se inoculadas em galinhas, proteÌnas de coelho, a resposta imune È bastante elevada. Isto acontece porque quanto mais prÛxima for a relaÁ„o filogenÈtica, menor ser· o estÌmulo e vice- versa. Existe pouca diferenÁa entre as proteÌnas de galinhas e patos e muita diferenÁa entre as proteÌnas de aves e mamÌferos. Embora este conceito da relaÁ„o filogenÈtica reflita boa parte das aplicaÁıes imunolÛgicas, n„o pode ser tomado como regra. A induÁ„o de uma resposta imune muito especÌfica È funÁ„o direta da semelhanÁa biolÛgica entre a fonte do antÌgeno e o animal receptor, ainda que seja menos intensa. Lebres e coelhos pertencem ‡ mesma famÌlia e s„o bastante semelhantes, tanto morfolÛgica quanto fisiologicamente. Portanto, ao se injetar proteÌnas de coelho em lebre, poder· se obter anticorpos muito especÌfi- cos, ou seja, anticorpos que sÛ reagem contra proteÌna de coelho.
4.3. Peso molecular e complexidade molecular
Na maioria dos antÌgenos, quanto maior for a molÈcula, maior ser· o n˙mero de epitopo; e quanto maior a complexidade, maior ser· a imunogenicidade. Um antÌgeno complexo contÈm v·rios determinantes antigÍnicos, onde alguns dos quais s„o mais eficientes na induÁ„o da resposta imune e s„o chamados imunodominantes.
4.4. ConfiguraÁ„o espacial e acessibilidade
A imunogenicidade e a antigenicidade de uma proteÌna n„o depende apenas de sua estrutura prim·ria (isto È, da sequÍncia de amino·cido), mas tambÈm das estruturas secund·rias, terci·rias e atÈ quatern·rias. Assim, se tratar- mos uma proteÌna pelo calor, ou agentes quÌmicos desnaturantes, e inocularmos esta em um animal, poderemos obter a formaÁ„o de anticorpos com especificidade diferente do que se inocul·ssemos a proteÌna intacta. A configuraÁ„o espacial de
| 35
diversos epitopos em uma ˙nica molÈcula de proteÌna pode influenciar a ligaÁ„o do anticorpo de v·rias formas (Figura 4). A ·rea importante para a imunogenicidade deve ficar acessÌvel, na superfÌcie da molÈcula.
Figura 4. DistribuiÁ„o dos determinantes antigÍnicos sequenciais e n„o sequenciais em uma macromolÈcula proteica
4.5. Forma de administraÁ„o e adjuvantes
A dose do antÌgeno, a via e o esquema de imunizaÁ„o, assim como o uso de adjuvantes, s„o fatores atuantes na induÁ„o da resposta imune. As vias de inoculaÁ„o subcut‚nea, intradÈrmica e intramuscular levam geralmente os imunÛgenos para os nÛdulos linf·ticos regionais, e, mais frequentemente, induzem a imunidade celular. Os antÌgenos inoculados por via endovenosa e intraperitonial acumulam-se predominantemente no baÁo, e mais frequente- mente induzem a uma imunidade humoral. O adjuvante melhora a imunogenicidade de compostos com ele misturado, sem interferir na especificidade da resposta. Em medicina preventiva, s„o muitas vezes adicio- nados ‡s vacinas para reduzir a dose e a frequÍncia de injeÁıes dos antÌgenos utilizados para a imunoprofilaxia de doenÁas infecciosas. Normalmente, o antÌgeno È aprisionado por ele, formando depÛsitos, o qual È liberado aos poucos por perÌodo de tempo mais extenso. Com isso, h· o aumento do tempo de exposiÁ„o do antÌgeno no organismo pelo retardamento de sua
Imunologia
| 37
da resposta imune que, frequentemente, resultam em anular a aÁ„o de biopatÛgenos, por meio da ativaÁ„o do sistema complemento, opsonizaÁ„o dos antÌgenos para fagocitose, citotoxicidade celular dependente de anticorpo (ADCC), em que os anticorpos marcam os microrganismos para serem destruÌdos pelas cÈlulas do sistema imune inato e reaÁıes de hipersensibilidades, entre outras ocorrem.
Estas funÁıes s„o estruturalmente separadas na molÈcula e a regi„o de ligaÁ„o ao antÌgeno varia amplamente, sendo conhecida como regi„o vari·vel ou regi„o V. A regi„o molecular que participa da funÁ„o efetora È conhecida como regi„o constante ou C, e n„o varia do mesmo modo, embora apresente cinco formas principais que se especializaram na ativaÁ„o de diferentes mecanismos.
A not·vel diversidade das molÈculas dos anticorpos È consequÍncia de um mecanismo altamente especializado, pelos quais os genes expressos s„o reunidos por rearranjos de DNA, que juntam dois ou trÍs diferentes segui- mentos para formar um gene de regi„o vari·vel durante o desenvolvimento das cÈlulas B. Subsequentes rearranjos nucleicos podem reunir o gene composto da regi„o vari·vel e qualquer gene da regi„o constante, produzindo assim anticorpos de cada um dos 5 isotipos.
Estruturalmente (Figura 5), a imunoglobulina È formada por duas cadeias leves (L-light-leve), idÍnticas, constituÌdas de polipeptÌdeos de cerca de 25 mil Daltons e de duas cadeias pesadas (H-heavy- pesado), tambÈm idÍnticas, com peso molecular de 50 mil Daltons ou mais. Cada cadeia leve est· ligada a uma cadeia pesada por pontes dissulfÌdricas. O n˙mero exato e as posiÁıes destas pontes entre as cadeias diferem entre as classes e subclasses de Imunoglobulinas. AlÈm disso, ambas as cadeias, leves e pesadas, possuem uma regi„o vari·vel e outra constante. Portanto, a imunoglobulina possui na cadeia leve uma regi„o constante (CL) e uma vari·vel (VL). O mesmo na cadeia pesada, uma regi„o constante (CH) e uma vari·vel (VH). Existem dois tipos de cadeias leves, akappa (k) e alambda (l). Em humanos, 60% das
Imunologia
38 | Conceitos e MÈtodos para a FormaÁ„o de Profissionais em LaboratÛrios de Sa˙de
cadeias leves s„o do tipokappa, e 40% s„o do tipolambda. Os primeiros 110, ou mais, amino·cidos da regi„o aminoterminal das cadeias leves ou pesadas variam muito entre os anticorpos de especificidade diferentes e por isto s„o chamadas de regi„o vari·vel.
A molÈcula de imunoglobulina pode ser digerida por enzimas proteolÌticas. A digest„o pela papaÌna quebra a molÈcula em trÍs fragmentos (Figura 5): dois fragmentos chamados Fab (fragment antingen binding), que se liga ao antÌgeno especÌfico, e um fragmento denominado Fc (fragment crystallizable, fragmento cristaliz·vel), por formar cristais quando armazenado em locais frios. Os fragmentos Fab s„o os que contÍm as cadeias leves (L) completas, emparelhadas com os domÌnios V (vari·vel) e C (constante) da cadeia pesada, enquanto o Fc, contÈm apenas o domÌnio C (constante). A papaÌna cliva a molÈcula na porÁ„o aminoterminal das pontes de enxofre, permitindo que as metades carboxiterminais da Fc permaneÁam unidas, dei- xando o fragmento Fc livre. J· a pepsina, cliva na mesma regi„o, mas na porÁ„o carboxiterminal das pontes dissulfrÌdicas, produzindo o (Fab)í 2 , onde os dois braÁos dos Ac permanecem unidos.
Figura 5. Estrutrua b·sica de uma imunoglobina e a formaÁ„o dos fragmentos pela digest„o enzim·tica.