


UNIVERSIDADE FEDERAL DA BAHIA
FACULDADE DE FARMÁCIA
DEPARTAMENTO DO MEDICAMENTO
FARMACOLOGIA INTEGRADA
REVISÃO DO SISTEMA CARDIOVASCULAR
SALVADOR
2011








































Estude fácil! Tem muito documento disponível na Docsity

Ganhe pontos ajudando outros esrudantes ou compre um plano Premium

Prepare-se para as provas
Estude fácil! Tem muito documento disponível na Docsity
Prepare-se para as provas com trabalhos de outros alunos como você, aqui na Docsity
Encontra documentos específicos para os exames da tua universidade
Prepare-se com as videoaulas e exercícios resolvidos criados a partir da grade da sua Universidade
Responda perguntas de provas passadas e avalie sua preparação.
Ganhe pontos para baixar
Ganhe pontos ajudando outros esrudantes ou compre um plano Premium
REVISÃO DO SISTEMA CARDIOVASCULAR
Tipologia: Trabalhos
1 / 55

Esta página não é visível na pré-visualização
Não perca as partes importantes!
Trabalho entregue como instrumento de avaliação da disciplina Farmacologia Integrada ministrada pela professora Cristiane Villarreal.
Potenciais de ação e função dos íons Ca2+ O potencial de membrana em repouso do músculo cardíaco normal é de aproximadamente -85 a -95 milivolts (mV) e de cerca de -90 a -100 mV nas fibras de condução especializadas, as fibras de Purkinje. O potencial de membrana se eleva de seu valor normalmente muito negativo para um valor ligeiramente positivo, de +20mV. Em seguida, após a ponte inicial, a membrana permanece despolarizada por cerca de 0,2s no músculo atrial, e 0,3s, no músculo ventricular, formando o chamado platô , seguido por repolarização abrupta ao final do platô. No músculo cardíaco, o potencial de ação é causado pela abertura de dois tipos de canais: os canais rápidos de sódio, que são responsáveis quase que integralmente pelo potencial de ação do músculo esquelético – por súbita abertura de um grande número destes canais e influxo de um grande número de íons sódio – e por outra população inteira dos chamados canais lentos de cálcio. Estes permanecem abertos por alguns décimos de segundos e, durante este período, grande quantidade de íons sódio, como de íons cálcio, flui por esses canais lentos para o interior da fibra muscular cardíaca e isso mantém a despolarização por período prolongado, ocasionando o platô do potencial de ação. Além disso, os íons cálcio que penetram no músculo durante esse potencial de ação, saindo dos retículos sarcoplasmáticos, tem um papel importante ajudando a excitar o processo de contração muscular.
Regulação do bombeamento cardíaco Regulação intrínseca do bombeamento cardíaco. A regulação intrínseca de adaptação do coração à alteração no volume de sangue que entra é denominada mecanismo de Frank-Starling do coração. Este mecanismo indica que quanto mais o coração se enche, durante a diástole, maior vai ser a quantidade de sangue bombeada para a aorta. Quanto mais distendido o músculo cardíaco, mais força de contração. Ou seja, dentro dos limites fisiológicos, o coração bombeia todo o sangue que chega até ele, sem permitir acúmulo excessivo de sangue nas veias.
Curvas de função ventricular. As curvas de função ventricular são maneiras práticas de expressar a capacidade funcional do músculo cardíaco no bombeamento de sangue dos ventrículos. Um tipo é a curva do trabalho sistólico, que sinaliza o aumento do trabalho sistólico, até o limite da capacidade cardíaca, de acordo com o aumento da pressão atrial. Outra é a curva do débito-minuto ventricular, que demonstra o aumento do débito por minuto do volume ventricular de acordo com a elevação da pressão atrial. Ambas as curvas estão ilustradas na figura abaixo.
Controle cardíaco pelos nervos simpático e parassimpático. A capacidade de bombeamento do coração é muito controlada pelos nervos simpáticos e para simpáticos. A estimulação simpática aumenta igualmente a força com que o coração se contrai, aumentando também, como consequência tanto o volume bombeado como a pressão de ejeção. Uma forte estimulação parassimpática pode, de fato, fazer cessar por alguns segundos os batimentos cardíacos, mas depois o coração geralmente retoma a atividade. Além disso, a forte estimulação parassimpática diminui em até 20 a 30% a força de contração do coração. Não é uma grande diminuição, porque as fibras parassimpáticas distribuem-se principalmente para os átrios e pouco para os ventrículos onde ocorre à contração motriz do coração.
Freqüência e Contratilidade cardíaca. Após a freqüência cardíaca elevar-se acima de um nível crítico, por exemplo, a força do próprio coração diminui, presumivelmente devido ao uso excessivo de substratos metabólicos
Figura 1. (A) Curvas do trabalho sistólico em um cão, mostrando o trabalho sistólico em função das pressões atriais médias, esquerda e direita, e (B) Curvas aproximadas do débito-minuto ventricular esquerda e direito, para o coração humano.
ligando esses eletrodos a sistemas adequados de registro, as voltagens elétricas geradas durante cada batimento cardíaco podem ser registradas.
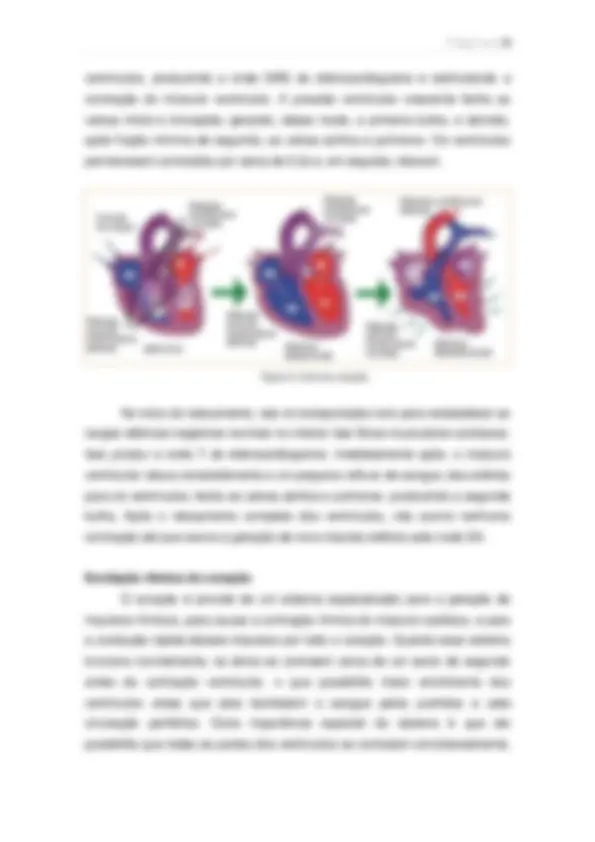
A onda P é causada pela disseminação da despolarização pelos átrios, e isso é seguido pela contração atrial. As ondas QRS resultam da despolarização elétrica dos ventrículos, o que inicia a contração ventricular. Portanto, o complexo QRS se inicia pouco antes do inicio da sístole ventricular. A onda T representa o estagio de repolarização dos ventrículos, quando suas fibras musculares começam a relaxar. Funcionamento das Valvas. As valvas A-V (tricúspide e mitral) evitam o refluxo de sangue dos ventrículos para os átrios durante a sístole, e as valvas semilunares (pulmonar e aórtica) impedem o refluxo da aorta e das aterias pulmonares para os ventrículos durante a diástole. Essas valvas se abrem e fecham passivamente, ou seja, se fecham quando um gradiente de pressão retrograda força o sangue de volta, e se abrem quando um gradiente de pressão para diante leva o sangue à frente. Átrios e Ventrículos como bombas. Cerca de 80% do sangue flui diretamente dos átrios para os ventrículos, mesmo antes da contração atrial. Então essa contração representa 20% adicionais para acabar de encher os ventrículos. Desse modo, os átrios funcionam simplesmente como bomba de
Figura 2. No eletrocardiograma são mostradas as ondas P,Q,R,S e T.
escova (primer pump), melhorando a eficácia do bombeamento ventricular por, no máximo 20%. Imediatamente após o inicio da contração ventricular a pressão ventricular sobre, de modo abrupto, fazendo com que as valvas A-V se fechem. É necessário mais 0,02 segundo para que o ventrículo gere pressão suficiente para abrir as valvas semilunares (aórtica e pulmonar). Esse é o período de contração isovolumétrica, onde há contração, mas não há esvaziamento. Após a sístole, as valvas aórtica e pulmonar se fecham (quando diminui a pressão nos ventrículos). Durante 0,06 segundo o músculo ventricular continua relaxando, mesmo que o volume não se altere, originando o período de relaxamento isovolumétrico. Durante esse período, as pressões intraventriculares diminuem rapidamente, de volta a valores diastólicos. É então que as valvas A-V se abrem para inicia novo ciclo de bombeamento ventricular. Relação entre os sons e o bombeamento cardíaco. Ao auscultar o coração com um estetoscópio, não se ouve a abertura das valvas. Porém, quando elas se fecham, os folhetos valvares e o liquido que as banham vibram, originando sons. Quando os ventrículos se contraem, ouve-se primeiro o som causado pelo fechamento das valvas A-V. Essa vibração é chamada de primeiro som cardíaco, ou primeira bulha. Quando as valvas aórticas e pulmonar se fecham, ao final da sístole, ouve-se o segundo som cardíaco, ou segunda bulha. Conceitos de pré-carga e pós-carga. Para a contração cardíaca, a pré- carga é geralmente considerada como a pressão diastólica final quando o ventrículo está cheio. A pós-carga do ventrículo é a pressão na artéria à saída do ventrículo. Assim, a pressão durante o enchimento do ventrículo é a pré- carga, e a pressão arterial contra a qual o ventrículo deve exercer a contração é a pós-carga. Contração Cardíaca. O ciclo cardíaco começa como a gênese do impulso rítmico no nodo AS. Em seguida, a transmissão desse impulso pelo e para o coração faz com que as fibras musculares se contraiam. Dessa forma, a onda P do eletrocardiograma ocorre imediatamente antes do inicio da onda de pressão atrial, causada pela contração atrial. Aproximadamente 0,16s após o início da onda P, o impulso elétrico já concluiu sua passagem pelos átrios, pelo nodo AV e pelo feixe AV. Logo começa a se propagar rapidamente pelos
o que é essencial para a geração efetiva de pressão nas câmaras ventriculares. Infelizmente, porém, este sistema rítmico e condutor do coração é muito suscetível a danos por doenças cardíacas, especialmente a isquemia dos tecidos cardíacos decorrente do fluxo sanguíneo coronário insuficiente. A consequência é, muitas vezes, um ritmo cardíaco desordenado ou uma seqüência anormal da contração das câmaras cardíacas, e a eficácia do bombeamento cardíaco muitas vezes fica gravemente afetada, a ponto de causar a morte do paciente. O nodo sinusal. O nodo sinusal é uma pequena tira achatada e elíptica de músculo especializado. As fibras deste nodo quase não têm filamentos contrateis e são contínuas com as fibras atriais, de modo que qualquer potencial de ação que se inicia no nodo sinusal espalha-se imediatamente para os átrios. Ritmicidade automática das fibras sinusais. Muitas fibras cardíacas têm a capacidade de auto-excitação. Isto é particularmente verdadeiro para as fibras do sistema especializado de condução do coração; a parte desse sistema que apresenta o maior grau de auto-excitação são as fibras do nodo sinusal que controla normalmente a freqüência de batimento de todo o coração. O potencial da fibra nodal sinusal entre as descargas tem negatividade de apenas -55 a -60 mV, em comparação com -85 a -90 mV para a fibra ventricular. A causa dessa negatividade reduzida é que as membranas celulares das fibras sinusais são naturalmente permeáveis a íons sódio. Auto-excitação das fibras do nodo sinusal. Os íons sódio tendem naturalmente a vazar para dentro das fibras do nodo sinusal por múltiplos canais na membrana, e esse influxo de cargas positivas também causa elevação do potencial de membrana. Assim, o potencial "de repouso" eleva-se gradualmente entre cada dois batimentos cardíacos. Ao atingirem a voltagem limiar, de cerca de -40 mV, os canais de cálcio-sódio são ativados, levando à entrada muito rápida tanto de íons cálcio como sódio e causando, assim, o potencial de ação. Assim sendo, é basicamente a permeabilidade intrínseca das fibras do nodo sinusal aos íons sódio que causa sua auto-excitação. Vias internodais e transmissão do impulso cardíaco pelos átrios. As extremidades das fibras do nodo sinusal fundem-se às fibras musculares atriais circundantes e os potenciais de ação que se originam no nodo sinusal dirigem-
se para adiante, por meio dessas fibras. Desse modo, o potencial de ação se propaga por toda a massa muscular atrial, e acaba por chegar também ao nodo A-V. A condução é mais rápida em vários pequenos feixes de fibras musculares atriais. A causa da velocidade de condução mais rápida nesses feixes é a presença de certo número de fibras de condução especializadas misturadas ao músculo atrial. Essas fibras são semelhantes às fibras de Purkinje dos ventrículos, de condução muito rápida. Transmissão no sistema de Purkinje. A transmissão muito rápida dos potenciais de ação pelas fibras de Purkinje é considerada como sendo causada pela maior permeabilidade das junções abertas nos discos intercalados entre as sucessivas células cardíacas que constituem as fibras de Purkinje. Nesses discos, os íons são facilmente transmitidos de uma célula para a seguinte, aumentando, assim, a velocidade de transmissão. As fibras de Purkinje também têm muito poucas miofibrilas, o que quer dizer que elas mal se contraem durante a transmissão de impulsos. Controle da excitação e condução no coração. Vimos que o impulso se origina normalmente no nodo sinusal. Entretanto, isso muitas vezes não ocorre, em condições anormais, pois outras partes do coração podem apresentar contrações rítmicas da mesma forma que as fibras do nodo sinusal; isto é particularmente verdadeiro para as fibras de Purkinje do nodo A-V. Então surge a pergunta: por que é o nodo sinusal que controla a ritmicidade do coração, e não o nodo A-V ou as fibras de Purkinje? O nodo sinusal controla, portanto, o batimento do coração porque sua freqüência de descarga rítmica é maior do que a de qualquer outra parte do coração. Portanto, o nodo sinusal é o marca-passo normal do coração. Papel do Sistema de Purkinje na causa da contração sincrônica do músculo ventricular. Através do sistema de Purkinje é que o impulso cardíaco chega a quase todas as partes dos ventrículos em intervalo de tempo muito curto, excitando a primeira fibra muscular ventricular apenas 0,06 s antes da excitação da última fibra muscular ventricular. Isso faz com que todas as partes do músculo ventricular de ambos os ventrículos comecem a se contrair quase que exatamente ao mesmo tempo. O bombeamento efetivo pelas duas câmaras requer esse tipo sinerônico de contração. Se o impulso cardíaco trafegasse muito lentamente pelo músculo ventricular, grande parte da massa
Mecanismo do efeito simpático. A estimulação dos nervos simpáticos libera norepinefrina nas terminações nervosas simpáticas, acredita-se que ele aumenta a permeabilidade da membrana das fibras ao sódio e ao cálcio. No nodo sinusal, um aumento da permeabilidade ao sódio ocasiona um potencial de repouso mais positivo e uma elevação mais rápida do potencial de membrana até o nível limiar de auto-excitação, acelerando o início da auto excitação, aumentando, portanto, a freqüência cardíaca. No nodo A-V, a maior permeabilidade ao sódio torna mais fácil ao potencial de ação excitar a parte subseqüente da fibra de condução, o que diminui o tempo de condução dos átrios para os ventrículos. O aumento da permeabilidade aos íons cálcio é pelo menos, parcialmente responsável pelo aumento da força contrátil do músculo cardíaco sob influência da estimulação simpática, porque os íons cálcio têm papel muito importante no desencadeamento do processo contrátil das miofibrilas.
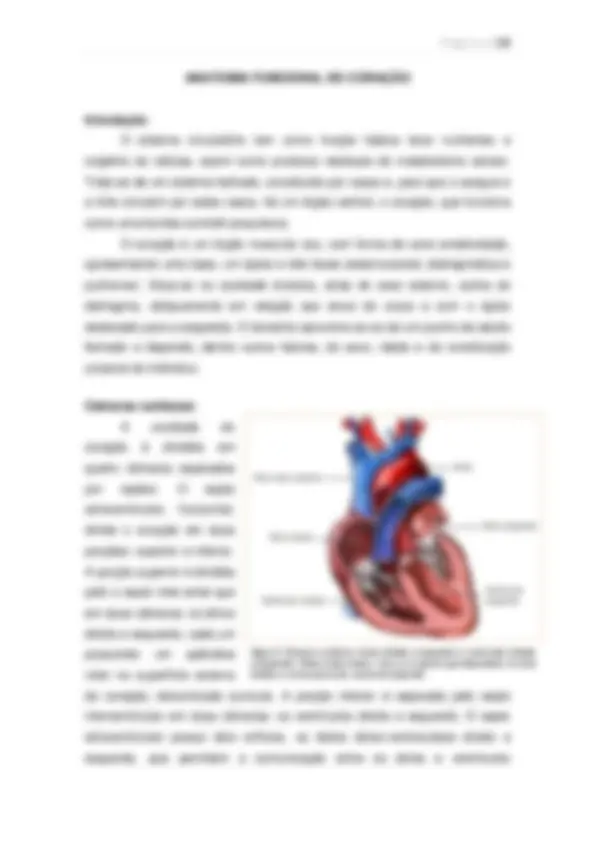
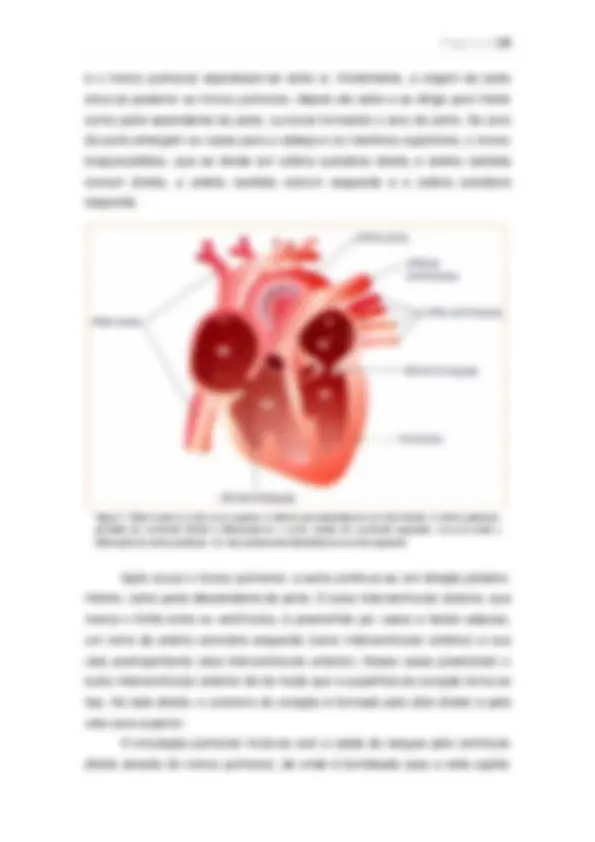
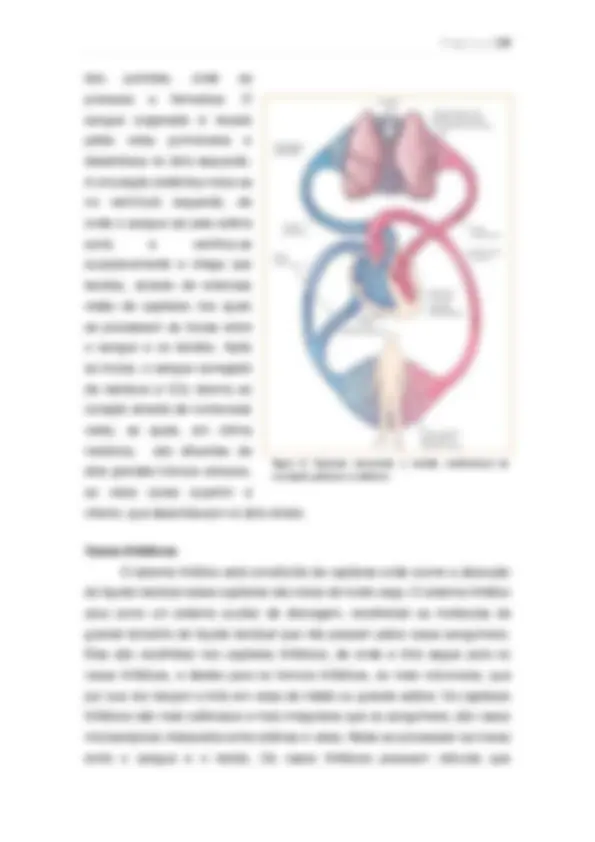
Visão geral da circulação A função geral da circulação é a de suprir as necessidades corporais – transportar até os tecidos os nutrientes, eliminar os produtos do metabolismo, levar hormônios de uma parte do corpo para a outra e, de modo geral, manter o ambiente apropriado em todos os líquidos teciduais do organismo para que as células sobrevivam e funcionem de maneira ótima. Partes funcionais da circulação: Artérias: Transportam o sangue, sob alta pressão, para os tecidos. Têm forte parede vascular e alta velocidade. Arteríolas: Pequenos ramos finais do sistema arterial; agem como condutos de controle, pelos quais o sangue é liberado para os capilares. Possuem forte parede vascular. Capilares: Sua função é a troca de líquidos, nutrientes, eletrólitos, hormônios e demais substâncias entre o sangue e o líquido intersticial. As paredes dos capilares são finas e possuem inúmeros poros permeáveis à água.
Vênulas: Coletam o sangue dos capilares e, de forma gradual, coalescem, formando veias progressivamente maiores. Veias: Funcionam como condutos para o transporte de sangue das vênulas de volta ao coração, além de atuarem como reservatório de sangue extra, de acordo com as necessidades da circulação. A sua parede é fina, mas suficiente para contrair e expandir.
Teoria básica da função circulatória Intensidade do Fluxo Sanguíneo. Quando os tecidos estão ativos, precisam de grande incremento do suprimento de nutrientes e, portando, de fluxo sanguíneo muito maior. Ainda assim o coração não pode aumentar seu débito por mais que quatro a sete vezes acima dos valores de repouso. Dessa forma, não é possível aumentar o fluxo sanguíneo em todas as partes do corpo quando um tecido particular demanda fluxo aumentado. Em vez disso, os microvasos, em cada tecido, monitoram, de modo contínuo, as necessidades teciduais, tais como a disponibilidade de oxigênio. Débito Cardíaco. Quando o sangue flui para um tecido, este sangue retorna imediatamente, pelas veias, para o coração. O coração responde de forma automática ao aumento da chegada de sangue, bombeando-o imediatamente para as artérias, de onde veio. Dessa forma, o coração age como um autômato, respondendo às demandas dos tecidos. No entanto é preciso auxílio na forma de sinais nervosos especiais, contribuindo para bombear a quantidade necessária de fluxo sanguíneo.
Figura 4. Visão geral da circulação.
somente durante a sístole cardíaca, e não ocorre fluxo durante a diástole. Entretanto, a complacência da árvore arterial reduz os pulsos de pressão, de modo que quase não ocorrem pulsos quando o sangue atinge os capilares. Assim, o fluxo sanguíneo tecidual é, essencialmente, contínuo, com pulsações muito pequenas. Fatores principais que afetam a pressão do pulso: Débito sistólico cardíaco; Complacência (distensibilidade total) da árvore arterial. Ressalta-se que a ejeção a partir do coração durante a sístole também é um fator, no entanto, menos importante. Quanto maior o débito sistólico, maior será a quantidade de sangue que deve ser acomodada na árvore arterial a cada batimento, portanto, maiores serão o aumento e a queda de pressão durante a sístole e a diástole, representando em maior pressão de pulso. Por outro lado, quanto menor for a complacência do sistema arterial, maior será o aumento da pressão, provocado por um dado volume de sangue bombeado, em cada batimento, para as artérias. A pressão de pulsos é determinada, em termos aproximados, pela proporção entre o débito sistólico e a complacência da árvore arterial. Logo, qualquer condição que afete esses fatores, afetará também a pressão do pulso.
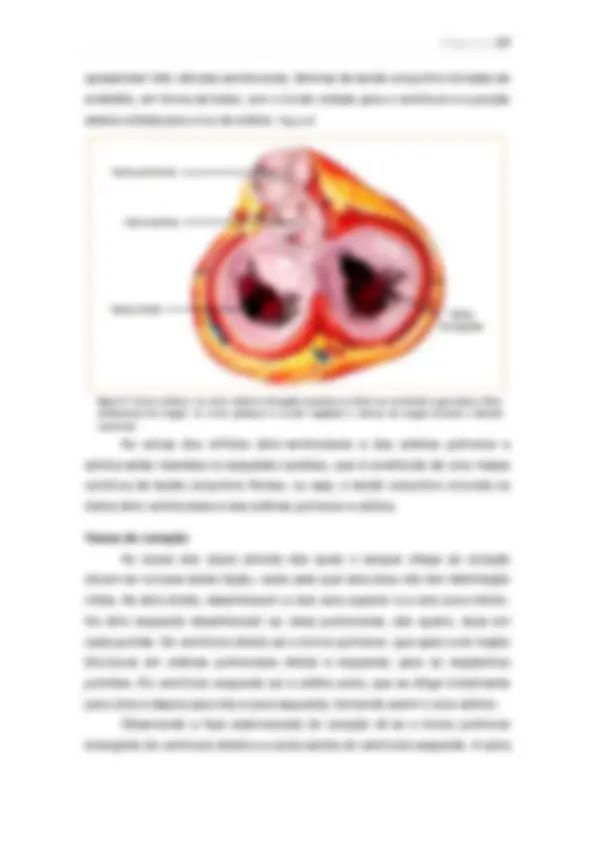
Microcirculação A microcirculação é responsável pelo transporte de nutrientes do sangue para os tecidos e a remoção dos produtos do metabolismo celulares. Apresenta organização conforme as necessidades específicas de cada órgão, mas o sangue entra nos capilares por meio de uma arteríola, passa para metarteríola até atingir os capilares verdadeiros de pequeno calibre, a seguir o sangue entra nas vênulas e retorna à circulação geral. A parede dos capilares é formada por uma única camada de células endoteliais e apresentam poros. A difusão é o principal meio de transporte entre as substâncias e o líquido intersticial, sendo que a medida que o sangue atravessa o capilar a água e substâncias dissolvidas difundem pela parede capilar. As partículas lipossolúveis como oxigênio e gás carbônico difundem diretamente sem passar pelos poros, por isso sua intensidade de transporte é maior que as substâncias hidrossolúveis como íons sódio e cloreto, água e glicose que só podem difundir
através dos poros. A intensidade dessa difusão depende da diferença de concentração entre o plasma e o liquido intersticial transportando do meio de maior concentração para o de menor, bastando pequena diferença de concentração para produzir transporte. O líquido intersticial tem composição parecida com a do plasma, mas apresenta uma quantidade menor de proteínas, fica entre filamentos de proteoglicanos e também fluindo livremente sem proteoglicanos. O movimento do líquido através da membrana capilar é determinado por quatro forças: a pressão capilar, que tende a forçar o líquido passar pelos poros para os espaços intersticiais; a pressão do liquido intersticial, tende a forçar o líquido para o interior do capilar; a pressão coloidosmótica do plasma, produzida por proteínas dissolvidas no plasma, que promove osmose de líquido para o capilar e a pressão coloidosmótica do capilar, que promove osmose de líquido para o espaço intersticial. A pressão coloidosmótica é produzida por proteínas dissolvidas no plasma, principalmente a albumina, pois não atravessam com facilidade os poros dos capilares sendo a concentração de proteínas no plasma maior que no líquido intersticial. A pressão do capilar na extremidade arterial é maior que na extremidade venosa, assim o líquido eflui na extremidade arterial e é reabsorvido (influi) na extremidade venosa. Sistema Linfático. É responsável pela reabsorção do líquido que flui para fora dos capilares arteriais de volta para o sangue pelos capilares linfáticos e também de proteínas que podem atravessar os capilares linfáticos sem impedimento. Quando a concentração protéica do liquido intersticial aumenta o volume e a pressão desse líquido, o retorno da proteína e do líquido pelo sistema linfático torna-se grande para balancear a intensidade do seu vazamento em um equilíbrio na concentração. Controle local do fluxo sanguíneo. Os tecidos controlam seu fluxo sanguíneo conforme suas necessidades metabólicas e quando ocorrem alterações na disponibilidade de oxigênio (que quando diminuída, o fluxo sanguíneo para os tecidos aumenta). Esse é o controle agudo, regulado pela adenosina, que provoca vasodilatação, pela dilatação natural do vaso conforme a demanda de oxigênio e nutrientes como a glicose. Também existe auto- regulação do fluxo sanguíneo com o aumento da pressão arterial e a dilatação
Já a estimulação parassimpática causa principalmente redução acentuada da freqüência cardíaca e ligeira redução na contratilidade cardíaca. O centro vasomotor, localizado bilateralmente na substância reticular do bulbo e no terço inferior da ponte, transmite impulsos descendentes pela medula e, daí, por todas as fibras vaso constritoras simpáticas para todos ou quase todos os vasos sangüíneos do corpo. Esse centro é responsável pelo tônus vasomotor que trata-se de um estado parcial de contração nos vasos sanguíneos, o que é significante para a manutenção da resistência dos vasos e por conseqüência da pressão arterial. Ao mesmo tempo que está controlando o grau de constrição vascular, o centro vasomotor também pode aumentar ou diminuir a atividade do coração, aumentando-a normalmente ao mesmo tempo que ocorre vasoconstrição em todo o corpo e diminuindo-a normalmente ao mesmo tempo que ocorre inibição da vasoconstrição. O centro vasomotor ainda pode sofrer inibição por diversos centros nervosos superiores como o hipotálamo, córtex cerebral, lobo temporal anterior entre outros, excitando-o ou inibindo-o. O efeito vasoconstritor e também vasodilatador do centro vasomotor é resultante da sua estimulação sobre a supra-renal, fazendo a secretar no sangue circulante tanto epinefrina como norepinefrina. Esses dois hormônios são levados pela corrente sanguínea a todas as partes do corpo, onde agem diretamente sobre os vaso sanguíneos, geralmente causando vasoconstrição, por conta da ação sobre os receptores alfa, mas, ocasionalmente, a epinefrina produz vasodilatação por ter um potente efeito estimulador dos receptores "beta", o que freqüentemente dilata os vasos. Papel do sistema nervoso no controle rápido da pressão arterial. Uma das funções mais importantes do controle nervoso da circulação é sua capacidade de causar aumentos muito rápidos da pressão arterial. Como conseqüência, três alterações principais ocorrem simultaneamente, cada uma das quais ajuda a aumentar a pressão arterial. Elas são as seguintes: Quase todas as arteríolas se contraem, levando ao aumento da resistência periférica. As veias em particular e outros grandes vasos da circulação contraem-se fortemente isso desloca o sangue para o coração, fazendo o bater mais com maior intensidade.
Estimulação do coração, fazendo com que ocorra aumento da freqüência cardíaca e também aumento da força de contração. O controle nervoso da pressão arterial é sem dúvida o mais rápido dos mecanismos de controle da pressão arterial. Os mecanismos reflexos para a manutenção da pressão arterial normal. Existem múltiplos mecanismos nervosos subconscientes para manter a pressão arterial em seu nível normal de operação ou próximo dele. Quase todos eles são mecanismos reflexos de feedback negativo são eles: Reflexo barorreceptor - É desencadeado por receptores de estiramento, denominados barorreceptores, os quais estão localizados na parede das grandes artérias sistêmicas. Uma elevação da pressão distende os barorreceptores e faz com que eles transmitam sinais para o sistema nervoso central, sendo, então, enviados sinais de feedback de volta à circulação, pelo sistema nervoso autonômico, para reduzir a pressão ao nível normal. Reflexo quimiorreceptor - Os quimiorreceptores são células quimiossensíveis a falta de oxigênio e ao excesso de dióxido de carbono ou de íons hidrogênio. Sempre que a pressão arterial cai abaixo de um nível crítico, os quimiorreceptores são estimulados, devido à diminuição do fluxo sanguíneo para eles, com a conseqüente diminuição da disponibilidade oxigênio e o acúmulo excessivo de dióxido de carbono e íons hidrogênio não removido pelo lento fluxo sanguíneo. Os sinais transmitidos dos quimiorreceptores para o centro vasomotor excitam esse centro e isso faz elevar-se a pressão arterial. Evidentemente, este reflexo ajuda a pressão a retornar ao nível normal quando ela cai a um nível muito baixo. Reflexos atriais - A distensão dos átrios causa a dilatação reflexa das arteríolas eferentes nos rins e também são transmitidos ao mesmo tempo sinais ao hipotálamo para diminuir a secreção de hormônio antidiurético, afetando, assim, indiretamente a função renal. A combinação desses dois efeitos causa rápida perda de líquido pela urina, que serve como poderoso meio para fazer o volume sanguíneo voltar ao normal e assim regular a pressão arterial.