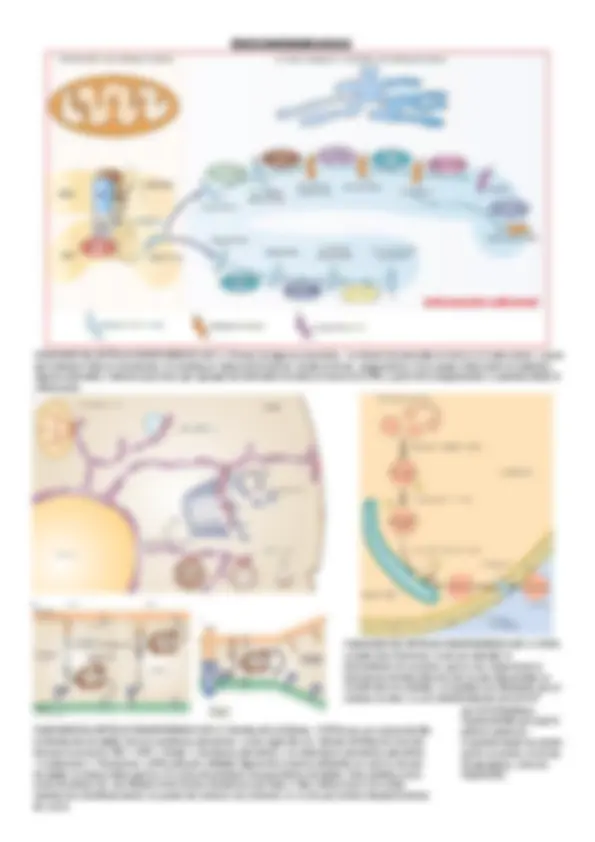
ENDOMEMBRANAS
Esquemageneraldecómoseintercambian
vesículasentrelosdiferentescompartimientos
delsistemadeendomembranas
Tiposdevesículascubiertasysuslocalizaciones.COPI
(COated Protein I),COPIIycubiertasdeclatrina (CCV,
Clathrin Coated Vesicles)