¡Descarga La pared celular y más Apuntes en PDF de Fisiología de las Plantas solo en Docsity!
Tema 2: La pared celular
La pared celular 3.1. Funciones y tipos: PC primaria y PC secundaria 3.2. Composición 3.2.1. Celulosa: estructura y síntesis 3.2.2. Polisacáridos de la matriz: hemicelulosas y sustancias pécticas 3.2.3. Proteínas: estructurales y enzimáticas 3.2.4 Lignina 3.4. Biogénesis de la pared celular 3.5. Plasmodesmos 3.6. La pared celular en la defensa de las plantas.
3. Pared celular 3.1. Funciones y tipos: PC primaria y PC secundaria 1. (^) Estructura celular externa de elevada resistencia mecánica 2. Red tridimensional de microfibrillas de celulosa en una matriz hidratada de polisacáridos, proteínas, fenoles y otros en una solución ligeramente ácida. 3. Muy dinámica 4. Continuidad molecular con plasmalema y citoesqueleto. 5. Su adquisición supuso la conquista el medio terrestre por las plantas. 6. No exclusiva de plantas: procariotas, hongos, algas.
La pared celular es una estructura común a todos los organismos del reino Planta, su composición varía entre especies, entre tejidos de una misma especie, entre células... En la célula vegetal, la pared se va depositando como una serie de capas. Así, durante la división celular se deposita la primera capa originando la lámina media , formada fundamentalmente por polisacáridos pécticos, que mantiene la unión entre las células adyacentes. Entre la lámina media y la membrana plasmática, se continúa depositando el material que va a constituir la pared celular. Una vez que la placa celular está completa, en las células hijas tiene lugar el depósito del material que constituye la pared celular primaria , de escaso grosor (0.1-1.0 μm). Las paredes primarias son de gran importancia en el proceso de extensión o expansión celular, pues controlan el crecimiento. En algunos tipos de células especializadas, una vez que dejan de crecer, se depositan nuevas capas de material en la cara interna de la pared, lo que constituye la pared secundaria , más gruesa que la primaria. La pared celular es una estructura altamente organizada, organizada por una red tridimensional de microfibrillas de celulosa embebida en una matriz constituida por polisacáridos (hemicelulosas y pectinas), proteínas y fenoles en una solución ligeramente ácida. A diferencia de lo que sugiere su nombre, no es una estructura estática sino un compartimento metabólico dinámico y mantiene continuidad molecular con la membrana plasmática y el citoesqueleto. La pared tiene una gran resistencia mecánica, y define el tamaño y la forma de la célula, al controlar en gran parte el crecimiento celular. Tiene también un papel importante en la defensa de las plantas frente a organismos potencialmente patógenos, al actuar como barrera física y como fuente de moléculas con actividad biológica (oligosacarinas), implicadas en el control de mecanismos de defensa.
Funciones esenciales para el crecimiento, desarrollo, mantenimiento y reproducción :
- (^) Define la forma de la célula
- Resistencia mecánica: 2.a. Unión entre células 2.b.Vegetales gran porte 2.c. Resistencia tensión del xilema
- Regula el volumen celular: control del crecimiento
- Relaciones hídricas de la planta: Presión de Turgencia.
- (^) Reserva: polisacáridos en PC endospermo y cotiledones de las semillas
- Defensa: como barrera física, y como fuente oligosacarinas. Son fragmentos de pared celular biológicamente activos, que pueden estar implicados en respuestas defensivas o relacionadas con procesos
- Gran importancia económica e industrial (leña, carbón, papel, tejidos, fibras en la dieta humana).
PC: tamaños, formas y composición muy variadas Dos grandes tipos en función de su desarrollo: 1.- PC primaria: Células jóvenes en crecimiento. Delgadas y simples (excepciones: colénquima y epidermis). 2.- PC secundaria: Después de elongación celular. 1 ó más paredes gruesas. Altamente especializada en estructura y composición Fuerza y dureza → crecimiento vertical. Existencia de huecos: sólo PC1 (Pit /pit fields: en la pared celular primaria hay grupo de agujeros. Estos agujeros se llaman plasmodesmos y van a través de la pared celular primaria. Un grupo de plasmodesmos se denomina campo pit primaria).
Lámina media: 1 capa fina Zona de contacto en PC de células vecinas. Muy rica en pectinas, y otras proteínas ≠ Origen: placa celular.
Las células vegetales están rodeadas por paredes celulares rígidas Una diferencia fundamental entre las plantas y los animales es que cada célula vegetal está rodeada por una pared celular rígida. En los animales, las células embrionarias pueden migrar de una posición a otra, dando lugar al desarrollo de tejidos y órganos que contienen células que se originan en diferentes partes del organismo. En las plantas, tales migraciones celulares no ocurren porque la pared de cada célula y su vecina están unidas a través de la lámina media. En consecuencia, el desarrollo vegetal, a diferencia del desarrollo animal, depende sólo de los patrones de división y crecimiento celular. Las células vegetales tienen dos tipos de paredes: primaria y secundaria (Figura 1.2). Las paredes celulares primarias suelen ser finas (menos de 1 mm) y son características de células jóvenes en crecimiento. Las paredes celulares secundarias son más gruesas y resistentes que las paredes primarias y se depositan cuando la mayor parte de la elongación celular ha finalizado. Las paredes celulares secundarias deben su fuerza y resistencia a la lignina, un material quebradizo similar a la cola. Las discontinuidades en esta pared es lo que llaman “simple pits”; cuando coincide con un simple pit de la ´pared de la célula vecina se le llama “pit-pairs”. La evolución de las paredes celulares secundarias lignificadas proporcionó a las plantas la estructura reforzada necesaria para crecer verticalmente sobre el suelo y colonizar la tierra. Las briófitas, que carecen de paredes celulares lignificadas, son incapaces de elevarse más de unos pocos centímetros sobre el suelo. La lámina media es una capa fina que se observa con frecuencia en la zona en la que las paredes de las células vecinas entran en contacto. Tiene una composición distinta del resto de la pared: se caracteriza por ser muy rica en pectinas, y por tener proteínas distintas del resto de la pared. Su origen es la placa celular, que se forma cuando se separan dos células hijas.
(d) Arabinogalactano-proteínas (AGP) ii) Proteínas enzimática a. (^) Oxidorreductasas (peroxidasas, lacasas, NADPH-oxidasa) b. Transglicosidasas (XET;XEH) c. _Expansinas 1.α-expansinas 2.β-expansinas
- Ligninas
- Agua_
3.2 Composición
3.2.1. Polisacáridos CELULOSA: Estructura y síntesis Muchas de las características fisiológicas y morfológicas de los tejidos vegetales se deben a las propiedades de las paredes de sus células; por eso, las células que realizan las diferentes funciones tienen paredes con distinta composición y estructura. Pero, en general, podemos considerar que una pared celular está formada mayoritariamente por polisacáridos (celulosa, hemicelulosas y pectinas), que constituyen alrededor del 90 % del peso seco de las paredes celulares primarias y entre el 65 y el 85 % de las secundarias. Las proteínas, tanto estructurales como enzimáticas, constituyen hasta un 10 % del peso seco de las paredes, mientras que las ligninas pueden tener una presencia importante (hasta un 35 %) en algunos tipos de paredes secundarias. Microfibrillas de celulosa : Estructura cristalina → estabilidad Hemicelulosas: Flexibles, unen moléculas de celulosa. Pectinas : Gel que embebe red de celulosa y hemicelulosa. Evita el colapso. Proteínas estructurales: Fuerza mecánica y ensamblaje. Ligninas: compacta e impermeabiliza
Esquema de los principales componentes estructurales de la pared celular primaria y su probable ordenamiento. Las microfibrillas de celulosa están cubiertas por hemicelulosa (como el xiloglucano) que se entrecruza para mantener las microfibrillas unidas una a otras. Las pectinas forman un gel que actúa como matriz de interconexión, entrando probablemente en relación con las proteínas estructurales.
El papel preciso delas proteína estructurales es incierto, pero es posible que aporten soporte mecánico y ayuda en el ensamblaje de otros componentes en la pared celular.
La celulosa es una microfibrilla fuertemente empaquetada de cadenas lineales de β-D-glucosa con enlaces (1→4). Cada resto de glucosa presenta una rotación de 180º con respecto a los restos contiguos. Debido a ello, la unidad que se repite en la celulosa es la celobiosa : dímero de β-D-glucosas que están unidas por enlace (1→4). A cada cadena de unidades de glucosa unidas se le puede denominar β-glucano. Las microfibrillas de celulosa tienen una longitud indeterminada y varían mucho en anchura y grado de ordenación, según su origen. Esta diversidad en la anchura corresponde a las diferencias en el número de
La celulosa está constituida por una larga secuencia lineal de restos de D-glucosa unidos por enlaces β (1— 4) (fig. 1.6). Cada resto presenta una rotación de 180º respecto a los restos contiguos, estabilizada por puentes de hidrogeno intramoleculares. El grado de polimerización medio en las paredes secundarias es alto, aproximadamente 104, mientras que en las paredes primarias es mas bajo, entre 2.6 y 6 103. Debido a su estructura, las cadenas de celulosa (40-70) se unen por puentes de hidrogeno intermoleculares formando agregados ( microfibrillas ), en su mayor parte cristalinos, en los que las cadenas individuales de celulosa se disponen de forma paralela. Las microfibrillas están separadas entre sí unos 30 nm; ese espacio está ocupado por los polisacáridos matriciales y por lignina o suberina, lo que confiere gran resistencia mecánica a la pared. Las microfibrillas de celulosa están presentes en las paredes de todas las plantas superiores, y constituyen el 30-40% del peso de las paredes de los tejidos leñosos, mientras que en las paredes primarias se encuentran en menor proporción.
La microscopía electrónica ha aportado evidencias que indican que las microfibrillas de celulosa son sintetizadas por grandes complejos proteicos llamados partículas rosetas o complejos terminales , que están embebidos en la membrana plasmática. Estas estructuras contienen muchas unidades de celulosa sintasa , el enzima que sintetiza los (1→4) β-D-glucanos individuales que forman las microfibrillas. La celulosa sintasa, que está localizada en el lado citoplasmático de la membrana plasmática, transfiere un residuo glucosa de un nucleótido dador de azúcar a la cadena creciente de glucano. Los glucósidos esteroles (esteroles unidos a una cadena de dos o tres residuos de glucosa) que actúan como aceptores iniciales, comienzan la síntesis de la cadena de glucano. El esterol es unido al glucano por una endoglucanasa y la cadena de glucano creciente es entonces empujada fuera de la membrana hacia el exterior de la célula, donde, junto con otras cadenas de glucano, cristaliza en una microfibrilla e interacciona con los Xiloglucanos y otros polisacáridos de la matriz. El nucleótido dador del azúcar es probablemente la uridinadifosfato-D-glucosa (UDP-glucosa). Tres genes diferentes codifican las proteínas que constituyen los monómeros que forma la celulosa sintasa
21 -La dirección en que se depositan las microfibrillas sintetizadas puede estar guiada por los microtúbulos adyacentes a la membrana -Las fibrillas son secretadas a un ambiente (la pared en formación) en el que hay otros polisacáridos (hemicelulosa, xiloglucanos..) que pueden unirse a la superficie de la celulosa y modificar la estructura general.
3.2.2. Polisacáridos HEMICELULOSAS: Estructura y síntesis Las hemicelulosas son un grupo heterogéneo de polisacáridos que se unen estrechamente a la PC. En la PC se encuentran varias clases de hemicelulosas. En la pared primaria de dicotiledóneas la hemicelulosa más abundante es el xiloglucano. Como la celulosa, este polisacárido tiene un esqueleto de residuos de β-D-glucosa con enlaces (1→4). No obstante, a diferencia de la celulosa, el xiloglucano tiene cadenas laterales cortas que contienen xilosa unida mediante enlace α (1→6), galactosa , y con frecuencia, aunque no siempre, una fucosa terminal. Estas sustituciones
3.2.2. Polisacáridos PECTINAS: Estructura y síntesis .
Los polisacáridos pécticos o pectinas son unos de los polímeros conocidos más complejos, constituidos por una mezcla de polímeros ácidos y neutros. Están presentes en las paredes celulares de todas las plantas, aunque su importancia cuantitativa varía desde alrededor del 35% en paredes primarias de dicotiledóneas hasta el 5% de las gramíneas. Se caracteriza por su capacidad de formar geles y se considera que su presencia en las paredes celulares determina su porosidad, proporciona superficies cargadas que modulan el pH y el balance iónico y sirven como moléculas de reconocimiento a señales tales como organismos simbióticos, patógenos e insectos. Las pectinas ácidas están constituidas por una alta proporción de restos de ácido D-galacturónico (forma ácida del monosacárido galactosa) unidos mediante un enlace α(1→4). Las sustancias pécticas son el constituyente fundamental de la lámina media, la primera capa que se forma y que luego queda como el cemento que une las paredes celulares adyacentes. Contienen otras formas ácidas de azúcares. Son los polisacáridos más solubles de la pared celular y se pueden extraer con agua caliente (mermeladas) o con quelantes de calcio. Las pectinas forman geles hidratados en los que los grupos carboxilos (carga -) se unen de forma iónica a Ca 2+.^ Se forma una red de pectinas unidas entre sí por puentes de calcio, ya que un calcio se puede unir a dos carboxilos. También existen otros tipos de unión. Las pectinas están sujetas a modificaciones que pueden alterar su conformación y uniones, produciéndose alteraciones en la carga y en el estado de gelificación, que puede afectar a las actividades de las enzimas de la pared celular.
- -Las hemicelulosas y las sustancias pécticas están altamente hidratadas y, a pesar de la formación de enlaces con las fibrillas de celulosa, la pared permanece lo suficientemente porosa como para permitir el paso de agua y la mayoría de los solutos en disolución. El control fino de la permeabilidad lo ejerce el plasmalema.
- -Aunque estos polisacáridos no tienen estructura cristalina, al estar altamente ramificados y ser más cortos que las que componen las microfibrillas de celulosa, tienden a ordenarse parcialmente en la pared, presumiblemente debido a la tendencia a alinearse a lo largo de los ejes de celulosa. Las cadenas constituidas únicamente por ácido galacturónico reciben el nombre de homogalacturonanos y su longitud es muy variable. Los grupos carboxilo de los restos galacturonosil pueden encontrarse esterificados con metanol o libres. Los grupos carboxilo libres pueden estar disociados y dar lugar a la formación de puentes de Ca 2 +, que constituyen la estructura conocida como caja de huevos.
Sin embargo, las cadenas de homogalacturonano no son muy frecuentes en las paredes celulares ya que normalmente presentan restos de L-ramnosa intercalados, enlazados en α(1→2). El ramnogalacturonano I (RG I) presenta una mayor proporción de ramnosa, a menudo en posición alternante, y son estos restos de ramnosa los puntos de anclaje de las cadenas laterales. Así, aproximadamente la mitad de los mismos puede estar unida en posición C-4 a cadenas constituidas por L-arabinosa y D-galactosa, con un grado de polimerización variable. Debido a que los restos de ramnosa se presentan con una frecuencia variable, dentro de las pectinas hay regiones mucho más ramificadas que otras. El ramnogalacturonano II (RG II) es un polisacárido relativamente pequeño (grado de polimerización aproximado de 60), con una estructura muy compleja ya que es el que presenta mayor número de azúcares y enlaces diferentes. Así, está constituido por ácido galacturónico, ramnosa, arabinosa y galactosa junto a pequeñas cantidades de azúcares poco frecuentes como 2-O-metilfucosa, 2-O-metilxilosa, apiosa, 3-C-carboxi-5-deoxi-L-xilosa y ácido 3-
deoxi-mano-octulosónico (KDO). A diferencia del RG I, los restos de ramnosa pueden presentar enlaces en 3, 3,4 y 2,3,4 o ser terminales. También se pueden formar enlaces con boro entre cadenas del RG II, posiblemente a través de la apiosa.
Las pectinas de naturaleza neutra están constituidas por L-arabinosa, D-galactosa o ambas. Los arabinanos están formados por restos de L-arabinosa unidos mayoritariamente por un enlace α(1→5), algunos unidos en 2 y 3 a otros restos de arabinosa. Los galactanos están formados por una cadena de β (1-4)-D-galactano con algunas ramificaciones de galactosa enlazadas en 6. Puede haber sustituciones de α (1-5)arabinano en proporción variable en el carbono 3 de los restos de galactosa, lo que recibe el nombre de arabinogalactano I. El arabinogalactano II es más abundante y contiene restos de D-galactosa enlazados en 3, 6, y 3, 6, y de L-arabinosa enlazados en 3 y 5. Tanto la arabinosa como la galactosa pueden asociarse a ácido ferúlico mediante un enlace éster, lo que posibilita la formación de enlaces entre distintas cadenas mediante puentes diferulil por la acción de las peroxidasas presentes en las paredes celulares.
3.2.3. Proteínas
Proteínas de la pared celular -Las estructurales normalmente se clasifican de acuerdo con el aminoácido que predomina en su composición: *glicoproteínas ricas en hidroxiprolina (HRGP) *proteínas ricas en prolina (PRP) *proteínas ricas en Gly (GRP) -La naturaleza de las proteínas de la pared celular varía mucho con el tipo celular, maduración, patógenos, heridas…También hay sitios específicos dentro de la célula, en los que predominan determinadas proteínas. Muchas proteínas estructurales tienen estructuras primarias altamente repetitivas, y en ocasiones están altamente glicosiladas. -La expresión de los genes que codifican las HRGP aumenta en respuesta a infecciones o a heridas. Por eso, se piensa que participan en la respuesta defensiva de las plantas aumentando la resistencia de la pared celular.
- Proteínas estructurales Las paredes celulares contienen también diferentes proteínas, tanto estructurales como enzimáticas, que en las paredes primarias llegan a constituir el 10 % de su peso. La mayoría corresponde a glicoproteínas, aunque el grado de glicosilación es muy variable, caracterizándose también por la presencia de secuencias repetitivas que pueden ser compartidas entre varias. Entre las proteínas estructurales, la proteína rica en hidroxiprolina HRGP (extensina) es la mejor caracterizada de todas ellas. Esta glicoproteína debe su nombre a la presencia de hidroxiprolina (Hyp) como aminoácido mayoritario (~46 %), que junto a otros cinco aminoácidos (serina, histidina, valina, tirosina y lisina) constituye más del 90 % de la misma. Está formada por numerosas secuencias repetidas de serina-(Hyp) 4 y tirosina-lisina-tirosina, responsables de su estructura secundaria y terciaria. Los residuos de tirosina pueden formar puentes intramoleculares de isoditirosina , que aumentan la rigidez y el carácter hidrófobo de la molécula. La glicosilación de la proteína tiene lugar a través de serina e hidroxiprolina, de manera que la serina se encuentra unida a un resto de galactosa, mientras que la hidroxiprolina sirve de anclaje a tri o tetraarabinanos que sirven para estabilizar la estructura de la proteína.
- Fuerza de compresión, resistencia a patógenos y herbívoros, impermeabilidad,…
- Pocas células pueden sintetizarla, sin embargo corresponde al 30% del C de la Biosfera.
- Células conductoras (vasos xilemáticos) y estructurales (fibras) con engrosamiento secundario
Lignina -Polímero fenólico con un patrón de uniones complejo e irregular de subunidades de alcoholes aromáticos. Estas subunidades se sintetizan a partir de la fenilalanina y se secretan hacia la pared, donde se oxidan por las enzimas peroxidada y lacasa. Cuando la lignina se forma en la pared, desplaza el agua de la matriz y forma una red hidrofóbica que une fuertemente la celulosa y evita su alargamiento. (fig. 15.18). La lignina se puede definir como un conjunto de polímeros que derivan de los monolignoles: alcohol cumarílico, alcohol sinapílico y alcohol coniferílico. A su vez, los monolignoles se sintetizan a partir del ácido p -cumarílico. Esos alcoholes se secretan a la pared, y en ella ocurre su polimerización, con intervención de oxidasas como peroxidasas y lacasas. -A medida que se forma lignina en la pared, desplaza el agua de la matriz, y se va creando un conjunto hidrófobo que se une a la celulosa e impide el crecimiento en longitud de la célula. -La lignina aporta fuerza mecánica y disminuye la susceptibilidad de la planta a ataques por patógenos, así como disminuye su digestibilidad para herbívoros. La estructura precisa de la lignina no se conoce debido a lo difícil que es extraerla de las plantas, donde se une covalentemente a la celulosa y a otros polisacáridos de la pared. La lignina se forma normalmente a partir de tres alcoholes fenil-propanoides diferentes: conierilico, coumarílico y sinapílico , alcoholes que se sintetizan a partir de la fenilalanina. Los alcoholes se unen en un polímero a través de la acción de enzimas que generan intermediarios con radicales libres. La proporción de las 3 unidades monoméricas en la lignina varía según la especie, órganos e incluso según las capas de una única pared celular. La lignina se encuentra en las paredes celulares de algunos tipos de tejidos de sostén (fibras) y vasculares (traqueidas), sobre todo en traqueidas y en los vasos del xilema. La rigidez mecánica de la lignina fortalece el tejido del tallo y vascular, posibilitando el crecimiento y permitiendo que el agua y los minerales vayan por el xilema en condiciones de presión negativa sin que el tejido colapse. Además de proveer soporte mecánico, la lignina tiene funciones protectoras importantes en plantas. su dureza física y química hace que sea pobre como alimento y poco digestible. La lignina bloquea el crecimiento de patógenos y es una respuesta frecuente a la infección o a heridas.
- Lignina: Nombre deriva del latín lignum (madera) y se ha empleado para describir la sustancia incrustante no celulósica presente en el tejido leñoso. Es el producto orgánico de plantas más abundante después de la celulosa. La lignificación causa la formación del tejido xilemático secundario en los árboles, así como un refuerzo del tejido vascular en plantas herbáceas. No se conoce un método que permita su aislamiento en estado nativo, sin alterar su estructura. Al contrario que los lignanos, es una mezcla racémica ópticamente inactiva. El nombre proviene del latín: lignum , madera, aunque también hay lignina en plantas no leñosas (tejidos vasculares de las plantas herbáceas). La lignificación supone un aumento de la rigidez y fuerza de compresión, y hace las paredes celulares hidrofóbicas e impermeables. -Después de la celulosa, es el compuesto vegetal más abundante (supone un 30% del C de la biosfera) -Es un polímero de fenilpropano muy ramificado -Función tanto primaria (estructural) como secundaria (defensa: la dureza hace que sea difícil de comer y, además, aumenta la indigestibilidad) -Unida covalentemente a la celulosa y a otros polisacáridos de la pared celular. En resumen, las ligninas son polímeros fenólicos que se encuentran en algunas paredes celulares vegetales. Estos polímeros contribuyen a la fuerza de compresión, resistencia a la degradación por microorganismos y a la impermeabilidad al agua de la matriz polisacárida y proteica de la pared celular. Las células capaces de acumular lignina no son muchas (elementos del xilema y elementos de sostén) (a), y frecuentemente tienen otras modificaciones, como engrosamiento de la pared celular secundaria, con objeto de contribuir al aumento de resistencia. De hecho, las funciones principales de la
lignina son estructurales y de transporte de agua y nutrientes minerales (primarias). Pero hay muchos tipos celulares que pueden sintetizar lignina en respuesta a herida o enfermedad (b) (función secundaria). Objetivos modificación genética de la cantidad de lignina o de la composición de la misma: aumento resistencia madera, aumento digestibilidad de plantas forrajeras, mayor facilidad de separación de la lignina de la celulosa en la fabricación de papel. La lignina de Gimnospermas (madera blanda) deriva fundamentalmente del alcohol coniferílico y, después, del p -coumarilico. En Angiospermas (madera dura), la lignina contiene alcohol coniferílico y sinapílico en proporción similar. En monocotiledóneas, suele haber más unidades del p-cumarílico que en dicotiledóneas. No obstante, hay excepciones a este patrón general. Además, la estructura de la lignina varía de especie a especie, entre los tipos celulares de una misma planta, e incluso entre diferentes partes de la pared celular de una misma célula. Además, también hay variaciones en la proporción en que se encuentran los monolignoles en la lignina sintetizada en respuesta a estrés, tanto abiótico como biótico. La existencia de más de un tipo de lignina y la precisa regulación de la composición de los monómeros de lignina durante el desarrollo y las respuestas a estrés, implica que las distintas ligninas pueden tener diferentes propiedades adaptativas o funcionales. Actualmente, se sabe que las ligninas están formadas por más monómeros en adición a los tres monolignoles. La lignificación es el proceso mediante el cual las unidades se unen entre sí, en reacciones que están acopladas a la formación de radicales. La reacción principal consiste en la adición de un monómero (generalmente un monoliognol) al polímero en crecimiento. La lignificación es un proceso distinto de la dimerización que origina los lignanos..
La síntesis de lignina, que tiene lugar en la propia pared celular, parece iniciarse con la oxidación de los monolignoles, presumiblemente por acción de una peroxidasa. In vitro , los radicales libres que resultan de la oxidación de monolignoles pueden reagruparse para formar estructuras diméricas que por oxidación y agrupación formarían los polímeros de lignina, sin necesidad de intervención de otras enzimas. No obstante, es posible que in vivo haya algún mecanismo de regulación de la lignificación Después del transporte de los monolignoles a la pared celular (no se sabe si vía vesículas Golgi o a través de bombas ABC del plasmalema), la lignina se forma a través de la polimerización deshidrogenativa de los monolignoles. La deshidrogenación de los monolignoles para formar los correspondientes radicales se ha atribuido a diversas enzimas: peroxidasas, lacasas, polifenol oxidasa y otras. Las peroxidasas usan H 2 O (^2) para oxidar sus sustratos; tampoco está completamente claro de dónde viene: actividad NADPH-oxidasa, amino-oxidasa dependiente de Cu, oxalato oxidasa, o la propia peroxidasa actuando sobre otros sustratos (NADPH, ascorbato, glutatión…). Las lacasas son glicoproteínas con Cu de la pared celular, que consumen O 2 para la oxidación. In vitro, pueden polimerizarse monolignoles y obtener lignina artificial empleando cualquiera de las dos enzimas. Antes de la síntesis de lignina, las células que van a formar el xilema secundario sufren cambios específicos irreversibles que finalmente llevan a la muerte celular y a la formación de elementos conductores y tejido de sostén (fibras). Estas células experimentan una expansión programada de sus paredes celulares, con deposición de celulosa, hemicelulosa, pectina y proteínas estructurales. Cuando se inicia la síntesis de lignina, los monolignoles son transportados desde el citosol a la pared celular, en determinadas localizaciones, y no de forma homogénea. La deposición controlada de monolignoles específicos crea dominios con distinta configuración estructural. Los monolignoles además pueden formar enlaces con otros polímeros de la pared celular, enlazando polisacáridos y proteínas con la lignina en una red tridimensional muy compleja. La síntesis de lignina se inicia en la lámina media y en las esquinas de la pared celular primaria una vez que se ha iniciado la síntesis de la pared secundaria (hay más lignina en esos sitios de iniciación que en
Las nuevas paredes primarias de ensamblan durante la citocinesis. Las paredes primarias se originan de novo durante las etapas finales de la división celular, cuando la placa celular recién formada separa las dos células hijas y genera una pared estable que es capaz de resistir la carga física de la presión de turgencia. Celulosa: en plasmalema con intervención de la celulosa sintasa. Polisacáridos de la Matriz: la mayoría sintetizados por enzimas unidos al Aparato de Golgi, y transportados hacia la pared en formación vía exocitosis de pequeñas vesículas. Enzimas responsables: azúcar nucleótido-polisacárido-glicosiltransferasas. Transfieren monosacáridos desde azúcares nucleótidos al extremo en crecimiento de la matriz polisacarídica. Proteínas: en el RE rugoso. Cuando son glicosiladas, lo son en el Golgi. -La formación de la pared celular se inicia en la placa celular , que da origen a la lámina media , la primera estructura que separa las dos células hijas. La placa celular se forma cuando las vesículas del aparato de Golgi y las cisternas del RE se agregan en la zona media del huso de la célula en división. Esta agregación está organizada por el fragmoplasto , un complejo ensamblaje de microtúbulos, membranas y vesículas que se forma en la anafase tardía o en la telofase temprana. -Las vesículas del Golgi están cargadas de pectina. Las vesículas se fusionan. Se está formando la placa celular; quedan dos membranas y entre ellas pectina. Mientras que se forma, hay partes que ya están atravesadas por prolongaciones del RE, que formarán los primeros plasmodesmos. Crecimiento centrífugo. Al final, la placa celular contacta con la membrana de la célula parental y se fusiona con ella. -La placa celular da origen a la lámina media, formada principalmente por polisacáridos pécticos. Entre lámina media y plasmalema se sigue depositando material, originándose la pared primaria. En algunos tipos de células especializadas, una vez que la célula ha dejado de crecer se depositan nuevas capas de material, y se forma la pared secundaria. Después de que se forme la pared, ésta puede crecer y madurar a través de un proceso que se puede esquematizar de la siguiente forma: Síntesis componentes → Secreción → Ensamblaje → Expansión (en células en crecimiento) → Cross- linking (entrecruzamiento) → Formación de la pared secundaria. Después de ser secretados al espacio extracelular, los polímeros de la pared deben ensamblarse es un estructura cohesiva; es decir, los polímeros individuales deben realizar el reordenamiento físico y los enlaces característicos de la pared. Tras su secreción los polímeros deben ensamblarse. Dos teorías:
- Autoensamblaje: tendencia a formar agregados de forma espontanea
- Ensamblaje mediado por enzimas: los enzimas de la pared pueden formar parte en la unión de la pared, enzimas: ▲ XET/XEH: Corta/pegan xiloglucanos, tiene la capacidad de romper el esqueleto de un xiloglucano y unir un extremo de un xiloglucano cortado con el extremo libre de otro xiloglucano aceptor, integrando en la pared los xiloglucanos recién sintetizado. ▲ Esterasas : Hidrolizan grupos ésteres que bloquean los grupos carboxilo de las pectinas, al desbloquear estos grupos carboxilos los enzimas aumentan la concentración de grupos ácidos en las pectinas e incrementan la capacidad de las pectinas para formar una red gel con enlaces Ca 2
- (^) (Hidrolizan grupos ésteres→ Aumento [grupos acídicos] de pectinas → aumento unión de Ca 2+ formación de redes gelificadas) ▲ Glucosidasas : Rompen cadenas laterales de hemicelulosas → aumento tendencia a adherirse a la superficie de las microfibrillas de celulosa. ▲ Oxidasas y peroxidadas : catalizan entrecruzamiento de grupos fenólicos en proteínas de la pared, pectinas, ligninas, etc.
3.4 Plasmodesmos
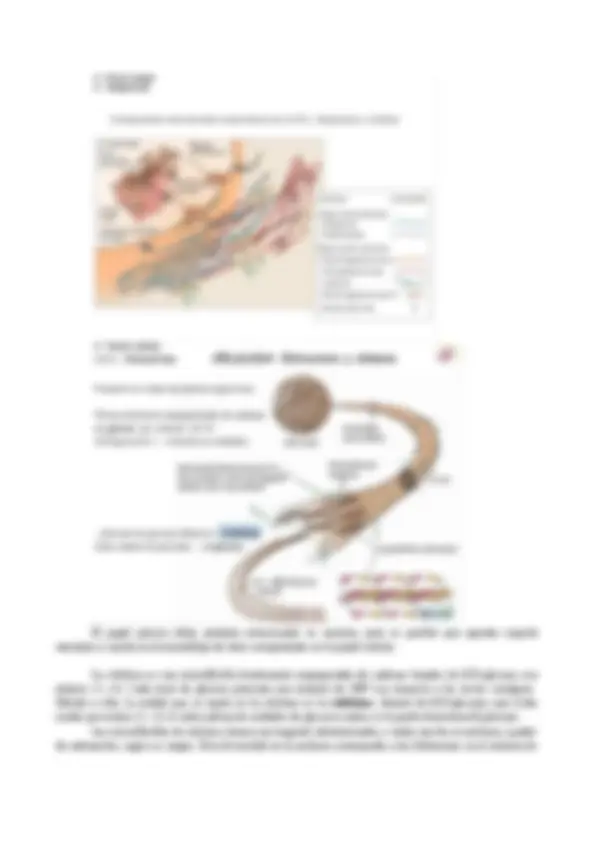
Los plasmodesmos son extensiones tubulares de la membrana plasmática, de 40 a 50 nm de diámetro, que atraviesan la pared celular y conectan el citoplasma de células adyacentes. Como la mayoria de las células vegetales están conectadas de esta forma, sus citoplasmas forman un sistema continuo conocido como simplasto. El transporte intercelular de solutos a través de los plasmodesmos se conoce; como transporte simplástico. Hay dos tipos de plasmodesmos: primarios y secundarios Los plasmodesmos primarios se forman durante la citocinesis, cuando las vesículas procedentes del Golgi que contienen los precursores de la pared celular se fusionan para formar la placa celular (la futura lámina media). Más que una lámina continua ininterrumpida, la placa celular recién depositada está atravesada por numerosos poros (Figura 1.27A), donde los restos del huso mitótico, formados por fragmentos del RE y microtúbulos, interrumpen la fusión de las vesículas. Las posteriores deposiciones de polímeros de la pared aumentan el grosor de las dos paredes celulares primarias a ambos lados de la lámina media generando canales lineales recubiertos con la membrana (Figura 1.27B). El desarrollo de los plasmodesmos primarios proporciona una continuidad y comunicación directa entre las células que están relacionadas clonalmente (es decir, procedentes de una misma célula madre). Los plasmodesmos secundarios se forman entre las células después de que sus paredes celulares se han formado. Se generan bien por evaginación de la membrana plasmática en la superficie celular o bien por ramificación de los plasmodesmos primarios. Además de aumentar la comunicación entre las células relacionadas clonalmente, los plasmodesmos secundarios permiten la continuidad simplástica entre las células que no están relacionadas clonalmente.
Transporte simplástico Como los poros nucleares, los plasmodesmos tienen una estructura interna compleja que funciona regulando el tráfico macromolecular de célula a célula. Cada plasmodesmo contiene un estrecho túbulo de RE llamado desmotúbulo (véase la figura 12.7). El desmotúbulo se continúa con el RE de las células adyacentes. Así, el simplasto une no sólo el citoplasma de células vecinas, sino también el contenido del lumen del RE. Sin embargo, no está claro que el desmotúbulo represente realmente un pasaje, dado que no parece haber un espacio entre las membranas, que están fuertemente comprimidas. Hay proteínas globulares asociadas a la membrana del desmotúbulo y a la membrana plasmática del poro (véase figura 12.7B). Estas proteínas globulares parecen estar interconectadas por extensiones radiales, dividiendo al poro en 8 ó 10 microcanales. Algunas moléculas pueden pasar de célula a célula a través de los plasmodesmos, probablemente fluyendo a través de los microcanales, aunque aún no se ha establecido la ruta exacta de comunicación. Siguiendo el movimiento de moléculas fluorescentes de diferentes tamaños a través de plasmodesmos que conectan las células epidérmicas de la hoja, Robards y Lucas (1990) determinaron que la masa molecular limitante para el transporte es de 700 al 000 daltons, equivalente a un tamaño molecular de 1,5 a 2,0 nm. Este es el tamaño límite de exclusión , o SEL , de los plasmodesmos.
3.5. La pared celular en la defensa de las plantas
_1. Define la forma de la célula
- Regula el volumen celular:_ control del crecimiento
- Relaciones hídricas de la planta
- Aumento de la resistencia mecánica.
- Reserva: polisacáridos en paredes de las semillas
Defensa vegetal contra patógenos Aunque carecen de sistema inmunitario, las plantas muestran una sorprendente resistencia a enfermedades producidas por hongos, bacterias, virus y nemátodos presentes en el ambiente. En esta sección examinaremos diferentes mecanismos desarrollados por las plantas para resistir a la infección, incluyendo la producción de agentes antimicrobianos y un tipo de muerte celular programada llamada respuesta hipersensible. Finalmente, analizaremos un tipo especial de inmunidad vegetal llamada resistencia sistémica adquirida. Una vez infectadas por un patógeno, las plantas despliegan un amplio espectro de defensas contra los microbios invasores. Una defensa común es la respuesta hipersensible , por la que las células que rodean la zona de infección mueren rápidamente, privando al patógeno de nutrientes y evitando su extensión. Después de una respuesta hipersensible satisfactoria, queda una pequeña zona de tejido muerto en el sitio donde se ha iniciado la invasión, pero impide el resto de la planta se vea afectada. La respuesta hipersensible con frecuencia está precedida por la producción de especies reactivas de oxígeno. Las células próximas a la infección sintetizan una enorme cantidad de compuestos tóxicos formados por la reducción del oxígeno molecular, incluidos el anión superóxido (0 2 -^ ) el peróxido de hidrógeno (H 2 O 2 ) y el radical hidroxilo (-OH). Se cree que una oxidasa dependiente de NADPH localizada en la membrana plasmática (Figura 13.25) produce 0 2 - , que es convertido posteriormente en -OH y H 2 02. El radical hidroxilo es el agente oxidante más fuerte de estas especies reactivas de oxígeno y es capaz de iniciar una serie de reacciones en cadena de radicales con una gran variedad de moléculas orgánicas, provocando la peroxidación de Iípidos, la inactivación enzimática y la degradación de ácidos nucleicos. Las especies reactivas de oxígeno pueden contribuir a la muerte celular como parte de la respuesta hipersensible o actuar para matar directamente al patógeno.
Muchas especies reaccionan a la invasión bacteriana o fúngica por síntesis de lignina o calosa. Se cree que estos polímeros sirven de barrera, frenando el avance del patógeno al resto de la planta y bloqueando físicamente su propagación. Una respuesta relacionada es la modificación de las proteínas de la pared celular. Ciertas proteínas de la pared ricas en prolina se llegan a entrecruzar oxidativamente después del ataque por patógenos en una reacción mediada por H 20 2 (véase la figura 13.25). Este proceso refuerza las paredes celulares en las proximidades del sitio de infección, aumentando su resistencia a la digestión microbiana. Otra respuesta de defensa a la infección es la formación de enzimas hidrolíticos que atacan la pared celular del patógeno. La invasión fúngica induce una gran cantidad de glucanasas, quitinasas y otras hidrolasas. La quitina, un polímero de residuos de N-acetilglucosamina, es el principal componente de las paredes celulares fúngicas. Estos enzimas hidrolíticos pertenecen a un grupo de proteínas que están estrechamente relacionadas con la infección patogénica y se conocen como proteínas relacionadas con patogénesis (PR). Las fitoalexinas. Quizás la respuesta mejor estudiada de las plantas a la invasión bacteriana y fúngica es la síntesis de fitoalexinas. Las fitoalexinas son un grupo diverso de metabolitos secundarios con una fuerte actividad antimicrobiana que se acumulan en los alrededores del sitio de infección. La producción de fitoalexinas parece ser un mecanismo común de resistencia a microbios patógenos para un amplio rango de plantas. Sin embargo, diferentes familias de plantas utilizan diferentes tipos de productos secundarios como fitoalexinas. Por ejemplo, los isoflavonoides son las fitoalexinas más comunes en las leguminosas, mientras que en Solanaceae, como la patata, el tabaco y el tomate, las fitoalexinas son diversos sesquiterpenos (Figura 13.26). Generalmente, las fitoalexinas son indetectables en la planta antes de la infección, pero se sintetizan rápidamente después del ataque microbiano debido a la activación de nuevas rutas biosintéticas. El punto de control normalmente es la iniciación de la transcripción génica. De esta forma, las plantas no parecen almacenar ninguna maquinaria enzimática necesaria para la síntesis de fitoalexinas. En su lugar, poco después de la invasión microbiana se transcriben y traducen los RNA (^) m adecuados, sintetizándose los enzimas de novo.
Las proteínas del inhibidor de la poligalacturonasa ( PGIPs ), también conocidas como proteínas de poligalacturonasa-inhibición , son fitoproteínas capaces de inhibir la acción de las enzimas de la poligalacturonasa producidas por los patógenos bacterianos y fungicidas