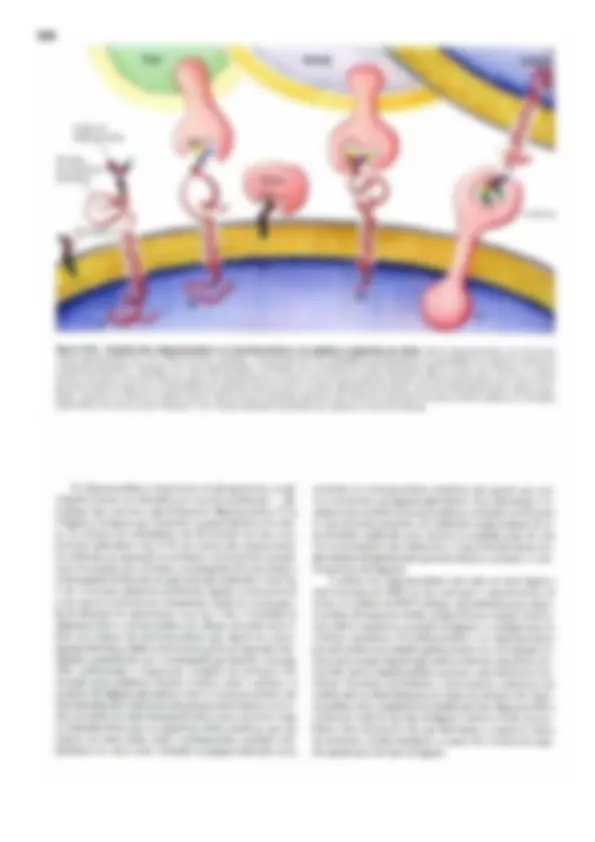
Pré-visualização parcial do texto
Baixe Carboidratos e outras Notas de estudo em PDF para Engenharia Agronômica, somente na Docsity!
CAPÍTULO 9 Carboidratos e Glicoconjugados Os carboidratos são as biomoléculas mais abundantes na face da Terra. A cada ano, a fotossíntese converte mais de 100 de toneladas de CO; e HO em celulose e outros produtos vege- tais, Certos carboidratos (açúcar comum e amido) são a base da dieta na maior parte do mundo e a oxidação dos carboidratos é a principal via metabólica fornecedora de energia para a maio- ria das células não-fotossintéticas. Polímeros insolúveis de car- boidratos funcionam tanto como elementos estruturais quanto de proteção nas paredes celulares bacterianas e de vegetais e nos tecidos conjuntivos de animais, Outros polímeros de carboidra- tos agem como lubrificantes das articulações esqueléticas e par- ticipam do reconhecimento e da coesão entre as células. Políme- ros mais complexos de carboidratos, ligados covalentemente a proteínas ou lipídios, agem como sinais que determinam a loca- lização intracelular ou o destino metabólico dessas moléculas hí- bridas, denominadas glicoconjugados. Este capítulo apresenta as principais classes de carboidratos e glicoconjugados e fornece al guns exemplos de seus variados papéis estruturais e funcionais. Os carboidratos são, predominantemente, poliidroxialdeí- dos ou poliidroxicetonas cíclicos, ou substâncias que liberam esses compostos por hidrólise, Muitos carboidratos, mas não todos, têm fórmulas empíricas (CH;0),; alguns também con- tém nitrogênio, fósforo ou enxofre. Os carboidratos estão divididos em três classes principais, de acordo com o seu tamanho: monossacarídeos, oligossacari- deos e polissacarídeos (a palavra “sacarídeo” é derivada do grego sakcharon que significa “açúcar”). Os monossacarídeos, ou açú- cares simples, consistem em uma única unidade de poliidroxial- deido ou cetona, O monossacarídeo mais abundante na natureza éo açúcar com seis átomos de carbono na molécula, a D-glicose, também chamada dextrose. Os oligossacarídeos são compostos por cadeias curtas de unidades monossacarídicas, unidas entre si por ligações cara terísticas, chamadas ligações glicosídicas. Os mais abundantes são os dissacarídeos, formados por duas unidades de monossa- carídeos. À sacarose, ou açúcar de cana, é o representante típico dessa classe. Ela é composta de dois monossacarídeos, com seis átomos de carbono, D-glicose e D-frutose. Todos os monossaca- rídeos e dissacarídeos comuns têm nomes que terminam com o sufixo “-ose”. Nas células, a maioria dos oligossacarídeos com trés ou mais unidades não ocorre como entidades livres, mas é unida a moléculas de não-açúcares (lipídios ou proteínas), for- mando glicoconjugados. Os polímeros de açúcares têm uma variação progressiva de tamanho de cadeia. Aqueles que contêm mais de 20 unidades são chamados polissacarídeos. Os polissacarídeos podem ter cadeias contendo centenas ou milhares de unidades monossaca- rídicas. Alguns, como a celulose, têm cadeias lineares, enquanto outros, como o glicogénio, tém cadeias ramificadas. Os polissa- carídeos de origem vegetal, amido e celulose, consistem em uni- ões Algodão pronto para a colheita. A fibra do algodão é formada de celulo- se, um polissacarídea com propriedades físico-químicas que lhe dão as caracteristicas têxtil desejáveis. O algodão tem sido cultivado e usado em produtos têxteis há milhares de anos. dades repetitivas de D-glicose, mas diferem entre si no tipo de ligação glicosídica, consequentemente têm propriedades e fun- ções biológicas diferentes. Monossacarídeos e Dissacarídeos Os carboidratos mais simples, os monossacarídeos, são aldeí- dos ou cetonas que contêm um ou mais grupos hidroxila na molécula; os monossacarídeos com seis átomos de carbono, gli- cose e frutose, têm cinco grupos hidroxila. Os átomos de carbo- no, nos quais os grupos hidroxila estão ligados, são geralmente centros qui os quais originam os numerosos açúcares este- reoisômeros encontrados na natureza. Iniciaremos descrevendo as famílias de monossacarídeos com esqueletos de três a sete áto- mos de carbono — suas estruturas e suas formas estereoisomé- ricas — e as maneiras de representar suas estruturas tridimensi- onais sobre o papel. Discutiremos, então, as diversas reações qui- micas dos grupos carbonila dos monossacarídeos. Uma dessas reações é a adição de um grupo hidroxila da mesma molécula, 225 226 que produz as formas cíclicas dos açúcares de cinco e seis áto- mos de carbono. (as quais predominam em solução aquosa) e cria um novo centro quiral, aumentando o grau de complexida- de estereoquímica dessa classe de compostos. A nomenclatura que especifica, de maneira não ambígua, a configuração ao re- dor de cada átomo de carbono na forma cíclica e a forma de representar essas estruturas no papel estão descritas com alguns detalhes a seguir; isso será muito útil mais tarde, quando da dis- cussão do metabolismo dos monossacarídeos. Apresentaremos aqui também alguns importantes deriva- dos de monossacarídeos, que serão encontrados nos capítulos seguintes. As duas famílias de monossacarídeos: aldoses e cetoses Os monossacarídeos são compostos incolores, sólidos cristali- nos, naturalmente solúveis em água, porém insolúveis nos sol- ventes não-polares. A maior parte deles tem sabor doce. O es- queleto molecular dos monossacarídeos comuns é constituído por uma cadeia carbônica não-ramificada na qual todos os áto- mos de carbono estão unidos entre si por ligações covalentes simples. Na forma de cadeia aberta, um dos átomos de carbono é unido por uma ligação dupla a um átomo de oxigênio para formar um grupo carbonila; cada um dos outros átomos de car- bono tem um grupo hidroxila. Se o grupo carbonila está em HZ 5 H—c—oH H y H—c—oH HO—C—H H—C—oH a ai a H—C—o0H H H CH;OH Gliceraldeído, Diidroxiacetona, D-Glicose, uma aldose uma cetose uma aldoexose fa) uma das extremidades da cadeia carbônica (isto é, em um aldeí- do), o monossacarídeo é uma aldose; se o grupo carbonila está em qualquer outra posição (em uma cetona), o monossacari- deo é uma cetose. Os monossacarídeos mais simples são as duas trioses com três átomos de carbono: o gliceraldeído, uma aldo- triose, e a diidroxiacetona, uma cetotriose (Fig. 9-1a). Os monossacarídeos com quatro, cinco, seis e sete átomos de carbono em seus esqueletos moleculares são chamados, res- pectivamente, tetroses, pentoses, hexoses e heptoses. Existem aldoses e cetoses correspondentes a cada um desses comprimen- tos de cadeia: aldotetroses e cetotetroses, aldopentoses e ceto- pentoses, e assim por diante. As hexoses, que incluem a aldoexo- se D-glicose e a cetoexose D-frutose (Fig. 9-1b), são os monos- sacarídeos mais comuns na natureza. As aldopentoses D-ribose e 2-desoxi-D-ribose (Fig. 9-1c) são componentes dos nucleotí- deos e dos ácidos nucléicos (Capítulo 10). Os monossacarídeos possuem centros assimétricos Todos os monossacarídeos, exceto a diidroxiacetona, contêm um ou mais átomos de carbono assimétrico (quiral) e, assim, ocor- rem em formas isoméricas opticamente ativas (págs. 45-47). A aldose mais simples, o gliceraldeído, contém um centro quiral (o átomo de carbono central); portanto, tem dois isômeros óp- ticos diferentes, ou enantiômeros (Fig. 9-2). Por convenção, uma dessas duas formas é designada isômero D e a outra H trem €=0 HO—C—H H—C—oH H—C—OH cHoH CH,0H D-Frutose, D-Ribose, 2-Desoxi-D-ribose, uma cetoexose uma aldopentose uma aldopentose tb) fe) Figura 9-1 - Monossacarídeos representativos (a) Duas trioses, uma aldose e uma cetose, O grupo carbonila está sombreado em vermelho em cada uma das estruturas. (b) Duas hexoses comuns. (c) As pentoses componentes dos ácidos nucléicos. A D-ribose é componente do ácido ribonucléico (RNA) e a 2-desoxi-D-nbose é componente do ácido desoxirribonucléico (DNA). Espelho CcH,OH Modelo bola-e-bastão HG CH,0H D-Gliceraldeido 1-Gliceraldeído Fórmulas em perspecti Figura 9-2 - Três maneiras de representar os dois isômeros do gliceraldeído. Os estereoisômeros são imagens especulares um do outro. O modelo bola-e-bastão mostra a configuração real das moléculas. Por convenção, na fórmula de projeção de Fischer, as ligações horizontais são projetadas para a frente do plano do papel, as ligações verticais para a parte de trás do plano do papel. Relenibrando (veja Fig. 3-7) que, nas fórmulas em perspectiva, as ligações em forma de cunha se projetam para a frente do plano do papel e as pontilhadas para trás do plano do papel 228 po carbonila. Cada uma das oito D-aldoexoses, as quais diferem estereoquimicamente em C-2, C-3 e C-4, têm nomes próprios: D-glicose, D-galactose, D-manose, etc. (Fig. 9-3a). As cetoses de quatro e cinco átomos de carbono são designadas pela inserção de “ul” no nome da aldose correspondente; por exemplo: D-ribu- lose é a cetopentose correspondente à aldopentose D-ribose, En- tretanto, existem cetoexoses que são denominadas contrariando essa regra como a frutose (do latim fructus, que significa “fruta”; as frutas são ricas nesse açúcar) e a sorbose (de Sorbus, gênero de árvore de montanha, a qual tem frutas que são ricas em sorbitol, um álcool derivado de açúcar). Dois açúcares que diferem somente na configuração ao redor de um único átomo de carbono são cha- mados epímeros; a D-glicose e a D-manose, que diferem estereo- quimicamente apenas em C-2, são epímeros, como também o são a D-glicose e a D-galactose (as quais diferem em C-4) (Fig. 9-4). 'cHo 'cHO o H=c—oH o HOC—H H+C—oH H+ÇC—oH HO-LC—H H=C—oH SSH FG=oR *CH;OH *CH;OH *CH,0H D-Manose D-Glicose D-Galactose fepímero em €-2) fepímero em C-4) 7 Figura 9-4 - Epímeros. A D-glicose e dois de seus epimeros são mostra dos em fórmulas de projeção. Cada epímero difere da D-glicose na confi- guração de apenas um centro quiral (sombreado em vermelho). Alguns açúcares ocorrem naturalmente na forma L; exem- plos são a L-arabinose e os L-isômeros de alguns derivados de açúcares (discutidos a seguir), componentes comuns dos glico- conjugados. H o Y H—C—oH HO—C—H HO—C—H CH,0H L-Arabinose Os monossacarídeos comuns possuem estruturas cíclicas Para simplificar, representamos as estruturas das várias aldoses e cetoses em forma de cadeia linear (Figs. 9-3 e 9-4), Na realida- de, em soluções aquosas, as aldotetroses e todos os monossaca- | £ | Figura 9.5 - Formação de hemiacetais e he- RE + micetais. Um aldeído ou uma cetona pode rea- H gir com um álcool, na proporção 1:1, produzin- ditas do um hemiacetal ou um hemicetal, respectiva- mente, criando um novo centro quiral no carbono da carbonila. A substituição de uma segunda mo- fécula de álcool produz um acetal ou cetal. Quan- do o segundo álcool faz parte de uma outra mo- RL-C=0 + | lécula de açúcar, forma-se a ligação glicosidica | (pão. 231) | Cetona rídeos com cinco ou mais átomos de carbono na cadeia ocor- rem, predominantemente, como estruturas cíclicas (anel) nas quais o grupo carbonila forma uma ligação covalente com o oxigênio de um grupo hidroxila ao longo da cadeia. A formação dessas estruturas em anel é resultado da reação geral entre aldeí- dos ou cetonas e álcoois, formando derivados chamados hemia- cetais ou hemicetais (Fig. 9-5), que apresentam um carbono as- simétrico adicional e assim podem existir em duas formas este- roisoméricas. Por exemplo: a D-glicose existe, em solução, como um hemiacetal intramolecular, no qual a hidroxila livre em C-5 reagiu com C-1 da função aldeído, formando um novo centro assimétrico e produzindo dois estereoisômeros, chamados a e (Fig. 9-6), Esses anéis de seis elementos são conhecidos por pi- ranoses, porque eles se assemelham ao anel de seis elementos do pirano (Fig. 9-7). A nomenclatura sistemática para os dois anéis da D-glicose são a.-D-glicopiranose e f-D-glicopiranose, As aldoexoses também existem em formas cíclicas com anéis de cinco átomos, as quais, devido à semelhança com o compos- to cíclico furano que possui estrutura química com cinco áto- o chamadas furanoses. Entretanto, o anel de seis átomos aldopiranosídico é muito mais estável que o anel aldo- furanosídico e predomina nas soluções de aldoexoses. Apenas aldoses contendo cinco ou mais átomos de carbono podem for- mar anéis piranosídicos, As formas isoméricas dos monossacarídeos, que diferem uma da outra apenas em sua configuração ao redor do átomo de car- bono do hemiacetal ou hemicetal, são chamadas anômeros. O átomo de carbono da carbonila ou hemiacetal é chamado de. carbono anomérico. Os anômeros a e f3 da D-glicose intercon- vertem-se, quando em solução aquosa, por meio de um proces- so chamado mutarrotação. Assim, uma solução que inicialmente contém apenas a-D-glicose e uma outra solução que contém apenas -D-glicose formarão, quando atingirem o equilíbrio, misturas idênticas, exibindo propriedades ópticas idênticas. Essa mistura consiste de aproximadamente um terço de a-D-glicose, dois terços de f-D-glicose e quantidades muito pequenas da for- ma linear e de anel de cinco carbonos (glicofuranose). As cetoexoses também ocorrem nas formas anoméricas a e B. Nesses compostos, o grupo hidroxila em C-5 (ou C-6) reage com o cetogrupo em C-2, formando um anel furanosídico (ou piranosídico) contendo uma ligação hemicetal (Fig. 9-5). A D- frutose forma predominantemente um anel furanosídico (Fig. 9-7); sendo que o anômero mais comum, em formas combina- das ou derivadas, é a P-D-frutofuranose. As fórmulas em perspectivas de Haworth, como aquelas na Figura 9-7, são comumente usadas para mostrar a forma em anel dos monossacarídeos. Porém, o anel piranosídico com seis áto- mos não é plano, como a perspectiva de Haworth sugere, mas tende a assumir uma das duas conformações em “cadeira” (Fig. 9-8). 7 3 | on HOR OR | = | R-C—OR H Hemiacetal HOR" R-C—OR + HO H Acetal HOR Alcool oH HOR! OR! HOR! RI-C—OR R HOR! RI-C—OR' + HO R Álcool Hemicetal Cetal H=C—oH HO-C—H D-Glicose H+C—OH D HC | dA mutarrotação 3H i HO N E 4 H H P-p-Glicopiranose Figura 9-6 - Formação das duas estruturas cíclicas da D-glicose. A reação entre o grupo aldeido em C-1 e o grupo hidroxila em €-5 forma uma ligação hemiacetal, gerando dois estereoisômeros, os anômeros a e P, que diferem somente na estereoquímica em torno do carbono do hemiacetal. A interconversão dos anômeros a e B é chamada mutarrotação. f-D-Glicopiranose HOcH 0 0H HH 1a H CHOH ho HH oH H P-D-Frutofuranose Furano t * q-D-Frutofuranose 9-7 - Piranoses e furanoses. As formas piranosídicas da D-gli- e as formas furanosídicas da D-frutose são mostradas aqui nas fór- em perspectiva de Haworth. As bordas do anel mais próximas do estão representadas por linhas grossas. Na perspectiva de Haworth, ps grupos hidroxila que estão abaixo do plano do anel devem estar do direito nas fórmulas de projeção de Fischer (compare com a Fig. 9- O pirano e o furano são mostrados para comparação 'H,0H o, pe HA H E He Pi H H e H H a-D-Glicopiranose (tb) Figura 9-8 - Fórmulas conformacionais das piranoses. (a) Duas for- mas em “cadeira” do anel piranosídico. Os substituintes nos carbonos do anel podem ser axiais (ax), projetando-se paralelos com o eixo verti- cal em relação ao plano do anel, ou, então, equatoriais (eq), projetan- do-se de maneira quase perpendicular ao mesmo eixo. Geralmente os substituintes nas posições equatoriais sofrem menos impedimento es- térico por substituintes vizinhos; portanto, as conformações com subs- tituintes volumosos na posição equatorial são favorecidas. Uma outra conformação em “barco” (não mostrada) somente aparece em deriva- dos com substituintes muito volumosos. (b) A conformação em “cadei- ra” da a-D-glicopiranose. 229 à esquerda). Além desses derivados ácidos das hexos: car ácido com nove átomos de carbono merece menç N-acetilneuramínico (ácido siálico), um derivado da N-acetil- manosamina, é um dos componentes de muitas glicoproteínas e glicolipídios de origem animal. Os grupos carboxila desses derivados de açúcar estão ionizados em pH 7, e esses compostos são, portanto, corretamente nomeados de carboxilatos, por exemplo, glucuronato, galacturonato, e assim por diante, Durante a síntese e a degradação metabólica dos carboidra- tos, frequentemente os intermediários não são os próprios açú- cares, mas os seus derivados fosforilados. A condensação do áci- do fosfórico com um dos grupos hidroxila de um açúcar forma um éster fosfórico, como na glicose-6-fosfato (Fig. 9-9). Os açú- cares fosfatados são relativamente estáveis em pH neutro e pos- suem uma carga negativa. Um dos efeitos da fosforilação do açú- car no interior das células é impedir a difusão desse açúcar para fora dela; a membrana plasmática das células, em geral, não pos- sui um transportador específico para essas moléculas. A fosfori- lação também ativa os açúcares para as transformações quími- cas subsequentes. Vários derivados fosforilados importantes dos açúcares serão discutidos no próximo capítulo. Os monossacarídeos são agentes redutores Os monossacarídeos podem ser oxidados por agentes oxidantes relativamente suaves, tais como os fons férrico (Fe'') e cúprico (Cu?!) (Fig. 9-10a). O carbono do grupo carbonila é oxidado a carboxila, A glicose e outros açúcares capazes de reduzir os íons férrico ou cúprico são chamados de açúcares redutores. Essa propriedade é a base da reação de Fehling, um teste qualitativo para a presença de açúcares redutores. É também possível deter- minar a concentração de um açúcar redutor pela medida da quantidade de agente oxidante que é reduzido pela solução des- se mesmo açúcar, Por muitos anos, esse teste foi usado para de- tectar e medir as elevações dos níveis de glicose no sangue e na urina no diagnóstico do diabetes melito. Atualmente, métodos mais sensíveis para a medida da glicose sangúínea empregam uma enzima, a glicose oxidase (Fig. 9-10b). Os dissacarídeos contêm uma ligação glicosídica Os dissacarídeos (como a maltose, a lactose e a sacarose) são constituídos por dois monossacarídeos unidos covalentemente entre si por uma ligação O-glicosídica, a qual é formada quan- do um grupo hidroxila de um molécula de açúcar reage com o átomo de carbono anomérico da outra molécula de açúcar (Fig. 9-11). Essa reação representa a formação de um acetal a partir de um hemiacetal (como a glicopiranose) e de um álcool (um grupo hidroxila de uma segunda molécula de açúcar) (Fig. 9-5). Quando um carbono anomérico participa de uma ligação gli- cosídica, ele não pode mais ser oxidado por um fon cúprico ou férrico. Esse açúcar contendo o átomo de carbono anomérico ligado não pode existir na forma linear e não pode mais agir como um açúcar redutor. Na descrição dos dissacarídeos ou polissacarídeos, a extremidade de uma cadeia que tem um car- bono anomérico livre (isto é, não envolvido em uma ligação gli- cosídica) é comumente chamada de extremidade redutora da cadeia. As ligações glicosídicas são facilmente hidrolisadas por ácido, mas resistem à clivagem por base. Assim, os dissacarídeos podem ser hidrolisados para liberar os seus componentes mo- nossacarídicos livres por aquecimento com ácido diluído. Um outro tipo de ligação glicosídica reúne o átomo de carbono ano- mérico de um açúcar a um átomo de nitrogênio em uma glico- proteína (veja Fig. 9-25). Essas ligações N-glicosil são também encontradas em todos os nucleotídeos (Capítulo 10). H OH B-D-Glicose HO D) vó id Do H=C—oH Ep ou HoÇ-H Nut >; H-/c—oH 6-0 H=C—oH H=t-0n *CH;0H cH;oH D-Glicose D-Gluconato (forma linear) (a) glicose ovidne D-Glicose + O, D-Glucono-d-lactona + H;O; (b) Figura 9-10 - Açúcares como agentes redutores. (a) A oxidação do carbono anomérico da glicose e outros açúcares é a base da reação de Fehling: o íon cuproso (Cu*) produzido em condições alcalinas forma o “óxido cuproso que é, um precipitado vermelho. Na forma hemiacetal (em anel), o C-1 da glicose não pode ser oxidado pelo Cu”. Entretanto, a forma em cadeia aberta está em equilibrio com a forma em anel e, eventualmen- te, a reação de oxidação chega a ser completa. A reação com Cu?* não é tão simples como a equação aqui apresentada; além do D-gluconato, vá- rias ácidos de cadeia curta são produzidos pela fragmentação da glicose (b) A concentração da glicose sangúinea é comumente determinada pela medida da quantidade de H;0; produzida pela reação catalisada pela glico- se oxidase. Na mistura reacional, uma segunda enzima, a peroxidase, Cata- lisa a reação da H;0; com um composto incolor para produzir um compo- nente corado, cuja quantidade é, então, medida por espectrofotometria. H,0H CH;0H O, heminectod H/4 H ) HO OH H OH dy H O a-D-Glicose P-p-Glicose trideólie|| condensação HO HO “cH,0H Maltose a-D-glicopiranosil-(1-»4)-D-glicopiranose Figura 9-11 - Formação da maltose. Um dissacarideo é formado a part de dois monossacarídeos (no caso desta figura, duas moléculas de D-glicose) quan- do uma —OH (álcool) da molécula de glicose (à direita) se condensa com o hemiacetal intramolecular de outra molécula de glicose (à esquerda), com a eliminação de H;O e a formação de uma ligação O-glicosídica. O reverso dessa reação é a hidrólise, ataque da ligação glicosídica pela H,O. A molécula da maltose retém um hemiacetal redutor em C-1, não envolvido na ligação glico- sídica. Asssm sendo, a mutarrotação interconverte as formas a em 3 do hemi- acetal. As ligações nessa posição, algumas vezes, são representadas por linhas onduladas para indicar que a estrutura pode ser tanto a quanto f. 231 232 A maltose, um dissacarídeo (Fig. 9-11), contém duas unida- des de D-glicose unidas por uma ligação glicosídica entre C-1 (o carbono anomérico) de uma unidade de glicose e C-4 da outra. Devido ao fato de o carbono anomérico livre (C-1 da unidade de glicose à direita da Fig. 9-11) poder ser oxidado, a maltose é um dissacarídeo redutor. A configuração ( do átomo de carbono anomérico envolvido na ligação glicose com o carbono anomérico livre é capaz de existir nas formas « e f3-piranosídicas. Várias regras devem ser seguidas para nomear os di deos, como a maltose, de forma clara e precisa, e especialmente para designar os oligossacarídeos mais complexos. Por conven- ção, o nome descreve o composto a partir de seu terminal não- redutor colocado à esquerda, sendo, então, “construído” na se- guinte ordem: (1) A configuração (a ou f) do átomo de carbo- no anomérico que retine a primeira unidade de monossacarídeo (à esquerda) à segunda é escrita. (2) É escrito o nome da unida- de da extremidade não-redutora. Para distinguir as estruturas dos anéis, cinco ou seis átomos, adiciona-se ao nome os termos “furano” ou “pirano”. (3) Os dois átomos de carbono reunidos pela ligação glicosídica são indicados entre parênteses, com uma seta conectando os dois números; por exemplo (1-4) mostra que C-1 da primeira unidade de açúcar está unido ao C-4 da segunda. (4) Escreve-se o nome da segunda unidade. Se existir uma terceira unidade, a segunda ligação glicosídica é descrita por meio das mesmas convenções. Para encurtar a descrição de polissacarídeos complexos são frequentemente empregadas abre- viações de três letras para cada monossacarídeo, como apresen- tado na Tabela 9-1. Seguindo essa convenção para denominar oligossacarídeos, a maltose será nomeada q.-D-glicopiranosil- (154)-B-D-glicopiranose, Como a maioria dos açúcares encon- trada neste livro é enantiômeros D e predomina a forma pirano- sídica das hexoses, usamos geralmente a versão curta do nome formal desses compostos, dando a configuração do carbono ano- mérico e nomeando os carbonos unidos pela ligação glicosídi- ca, Nessa nomenclatura abreviada, a maltose é nomeada Gle(a1-»4)Gle, Tabela 9-1 - Abreviações para os monossacarídeos comuns e al- guns de seus derivados Nome Abreviação | Nome Abreviação Abequose Abe Ácido glucurônico GIA Arabinose Ara Galactosamina GalN Frutose Fru Glucosamina GleN Fucose Fuc N-acetilgalactosamina GalNAc Galactose Gal N-acetilglucosamina GIcNAc Glicose Gle Ácido murâmico Mur Manose Man Ácido N-acetilmurâmico MurZAc Ramnose Rha Ácido N-acetilneuramínico NeuSAc Ribose Rib tácido siálico) Xilose Xyl O dissacarídeo lactose (Fig. 9-12) ocorre naturalmente ape- nas no leite; quando hidrolisado, libera D-galactose e D-glicose. O carbono anomérico da unidade de glicose está disponível para oxidação; portanto, a lactose é um dissacarídeo redutor. O seu nome abreviado é Gal(f31-»4)Gle, A sacarose (o açúcar comum) é um dissacarídeo de glicose e frutose. Ela é formada pelos vege- tais, porém não pelos animais superiores. Em contraste à mal- tose e à lactose, a sacarose não contém átomos de carbono ano- méricos livres; os átomos de carbono anoméricos de ambos os monossacarídeos estão envolvidos na ligação glicosídica (Fi g. 9-12). A sacarose é portanto um açúcar não-redutor. Os dissa- carídeos não-redutores são chamados glicosídeos; as posições H Lactose (forma f) B--galactopiranosil-(1-+4)-f-D-glicopiranose Gal(BI-»4)Gle CHOH Sacarose a-D-glicopiranosil B-D-frutofuranosídeo Gle(a162B)Fru Trealose a-D-glicopiranosil a-D-glicopiranosídeo Glefa tes ta)Gle Figura 9-12 - Alguns dissacarídeos comuns. Da mesma maneira que a maltose na Figura 9-11, esses dissacarideos são mostrados nas fórmu- las em perspectiva de Haworth. Os nomes comuns, os nomes sistemáti- cos completos e as abreviações são dados para cada um deles. ligadas são dos carbonos anoméricos. Na nomenclatura abrevia- da, uma seta dupla conecta os símbolos que especificam os car- bonos anoméricos e suas configurações. Por exemplo, o nome abreviado da sacarose é correto tanto na forma Gle(a.1<52B)Fru, como na forma Fru(B2€+1a.)Glc. A sacarose é o principal inter- mediário da fotossíntese; em muitos vegetais, ela é a principal forma na qual o açúcar é transportado das folhas até as demais partes da planta. A trealose Gle(a Le 1a)Glc (Fig. 9-12) é um dissacarídeo de D-glicose que, como a sacarose, é um açúcar não- redutor, Este é o principal constituinte da hemolinfa, o fluido circulante dos insetos, que serve como um composto de arma- zenamento de energia. Polissacarídeos A maioria dos carboidratos encontrados na natureza ocorre como polissacarídeos, polímeros de média até alta massa mole- cular. Os polissacarídeos, também chamados glicanos, diferem entre si na identidade das suas unidades monossacarídicas e nos tipos de ligação que os unem, no comprimento das suas cadeias e no grau de ramificação destas. Os homopolissacarídeos con- tém apenas um único tipo de unidade monomérica; os hetero- polissacarídeos contêm dois ou mais tipos diferentes de unida- des monoméricas (Fig. 9-13). Alguns homopolissacarídeos ser- vem como forma de armazenamento de monossacarídeos empregados como combustíveis pelas células; o amido e o glico- gênio são homopolissacarídeos desse tipo. Outros homopo! sacarídeos, como a celulose e a quitina, são utilizados como ele- mentos estruturais das paredes celulares vegetais e de exoesque- letos de animais, respectivamente. Os heteropolissacarídeos fornecem suporte extracelular nos organismos de todos os rei- Extremidade não-redutora L, ta) Ponto de ramificação Ramificação (0:16) (b) Extremidades não-redutoras Extremidade redutora o f Amilose Extremidades redutoras Amilopectina te) Figura 9-15 - Os dois polissacarídeos do amido, a amilose e a amilopectina (a) Um pequeno segmento de amilase, um polimero linear composto por unidades de D-glicose com ligações (cr! -»4). Uma única cadeia do polímero pade conter vários milhares de unidades de glicose A. amilopectina tem ligações semelhantes à amilose nas unidades entre os pontos de ramificação. (b) No ponto de ramificação, a amilopectina, tem uma ligação (a1-+6). (e) Um agregado de amilose e amilopectina como se acredita que ocorra nos grânulos de amido. As fibras de amilopectina (vermelho) formam estruturas em dupla hélice umas com as outras ou com as fibras de amilose (azul). As unidades de glicose, na extremidade não-redutora das ramificações externas, são retiradas enzimaticamente, uma de cada vez, durante a mobilização intracelular do amido para produção de energia. O glicogênio tem uma estrutura similar, mas é muito mais ramificado e mais compacto minais não-redutores. As enzimas de degradação, que agem so- mente nos terminais não-redutores, podem agir simultaneamen- te em muitos terminais, acelerando a conversão do polímero em monossacarídeo. Por que não estocar a glicose na forma monomérica? Foi cal- culado que os hepatócitos estocam glicogênio equivalente à con- centração de 0,4M de glicose. A concentração real de glicogênio, que é insolúvel e contri ouco para a osmolaridade do citosol, é cerca de 0,014uM. Se o citosol contivesse 0,4M de glicose, a os- molaridade poderia ser perigosamente alta, levando à entrada de água por osmose, o que poderia romper a célula (veja Fig. 4-11). Além disso, com uma concentração intracelular de glicose de 0,4M e uma concentração externa aproximadamente igual a 5mM (a concentração sangúínea em um mamífero), a variação da energia livre para a captação da glicose pela célula seria contra um gra- diente de concentração muito alto que poderia ser proibitivo. A estrutura tridimensional do amido é mostrada na Figura 9-16 e pode ser comparada com a estrutura da celulose descrita a seguir, A celulose e a quitina são homopolissacarídeos estruturais A celulose, uma substância fibrosa, resistente e insolúvel em água, é encontrada na parede celular dos vegetais, particularmente em troncos, galhos e em todas as partes lenhosas. A celulose consti- tui a maior parte da massa da madeira, e o algodão é celulose quase pura, Como a amilose e as cadeias principais da amilo- pectina e do glicogênio, a molécula de celulose é um homopo- lissacarídeo linear e não-ramificado, de 10 a 15 mil unidades de D-glicose. Mas há uma diferença muito importante: na celulose as unidades de glicose têm a configuração f (Fig.9-17), enquanto na amilose, na amilopectina e no glicogênio a glicose está na configuração a. As unidades de glicose na celulose estão unidas por ligações glicosídicas do tipo (1-»4). Essa diferença confere à celulose e à amilose estruturas tridimensionais e propriedades físicas muito diferentes entre si. A estrutura tridimensional das macromoléculas dos carboi- dratos está baseada nos mesmos princípios fundamentais da estrutura dos polipeptídeos: as subunidades com uma estrutura mais ou menos rígida, ditada por ligações covalentes, formam as estruturas macromoleculares tridimensionais, que são esta- bilizadas por interações fracas. Como os polissacarídeos têm muitos grupos hidroxila, a formação de pontes de hidrogênio tem uma influência especialmente importante em suas estrutu- ras. Os polímeros de B-D-glicose, como a celulose, podem ser representados por uma série de anéis piranosídicos rígidos co- nectados por um átomo de oxigênio que faz ponte entre dois átomos de carbono (ligação glicosídica). Existe rotação livre para a ligação C—O que liga as duas unidades (Fig. 9-17a). À confor- mação mais estável para o polímero é aquela na qual cada “ca- deira” é rodada 180º em relação às subunidades vizinhas, for- mando uma cadeia reta e estendida. Com várias cadeias estendi- das lado a lado, uma rede estabilizadora de pontes de hidrogénio inter e intracadeias produz fibras supramoleculares retas, estáveis e de grande resistência à tensão (Fig. 9-17b). A resistência à tensão da celulose tem feito dela uma substância muito útil para as civi- lizações através dos milênios. Muitos produtos manufaturados, incluindo o papel, papelão, raiom, placas de isolamento e outros materiais de empacotamento e construção, são derivados da celu- lose. A quantidade de água contida nesses materiais é baixa, por- que as ligações de ponte de hidrogênio intercadeias de celulose saturam sua capacidade de formação desse tipo de ligação. HO) AL o CH/OH Unidades de D-glicose ligadas (01-54) (a) Figura 9-16 - A estrutura do amido (amilose). (a) Em sua conformação mais estável, de “cadeiras” rigidas adjacentes, este polissacarídeo apresenta à cadeia curva de forma diferente da linear da celulose (veja Fig. 9-17). (b) Desenho em escala de um segmento da amilase. A conformação das Igações (u'1--»4) da amilose, da amilopectina e do glicogênio forçam estes polimeros a assumirem uma estrutura helicoidal estreitamente compacta. Estas estruturas compactas produzem os grânulos densos de estoca- gem de amido ou glicogênio vistos na maioria das células (veja Fig. 9-14). OHELEITITIQH 6 09 / MI TITIHO ; oH 4 OH Unidades de D-glicose ligadas (1-4) ta) (b) Figura 9-17 - A estrutura da celulose. (a) Duas unidades monossacarídicas de uma cadeia de celulose; as unidades de D-glicose têm ligação (31-54). As estruturas rigidas em “cadeira” podem girar, uma em relação à outra. (b) Desenho em escala de segmentos de duas cadeias paralelas de celulose , mostrando a conformação das unidades de D-glicose e as interligações proporcionadas pelas pontes de hidrogênio, Na unidade de hexose à esquerda, embaixo, estão mostrados tados os átomos de hidrogênio; nas outras três unidades de hexoses, todos os átomos de hidrogênio ligados a carbono foram omitidos, em benefício da clareza, já que eles não participam da formação das pontes de hidrogênio, Diferentemente das fibras lineares produzidas pelas ligações (B1-»4) em polímeros, como a celulose, a conformação mais favorável para ligação (a. 1-4) dos polímeros de D-glicose, como o amido e o glicogênio, é uma estrutura helicoidal compacta e estabilizada por pontes de hidrogênio (Fig. 9-16). O glicogénio e o amido, ingeridos na dieta, são hidrolisados por a-amilases, enzimas da saliva e das secreções intestinais que rompem as ligações glicosídicas (0:14) entre as unidades de glicose. A celulose não pode ser usada pela maioria dos animais como uma fonte de energia, porque falta-lhes uma enzima que hidrolise as ligações (31-54). Os térmitas, ou cupins, digerem facilmente a celulose (e, portanto, a madeira), mas isso só ocor- re porque o seu trato intestinal abriga um microrganismo sim- biótico, Trichonympha, que secreta uma enzima chamada celu- lase, a qual hidrolisa as ligações (B1->4) entre as unidades de glicose. Os fungos e as bactérias da madeira em decomposição também produzem celulase. Os únicos vertebrados que conse- guem utilizar a celulose como alimento são os bovinos e outros animais ruminantes (ovelhas, cabras, camelos, girafas). O estó- mago extra desses animais (rúmen) contém protistas e bactérias que secretam celulase. A quitina é um homopolissacarídeo linear composto por unidade de N-acetil-D-glucosamina em ligação 3 (Fig.9-18). Sua única diferença química com a celulose é a substituição de um grupo hidroxila em C-2 por um grupo amino acetilado. A q) na forma fibras estendidas similares àquelas da celulose e, como a celulose, não é digerível por animais vertebrados. A quitina é o principal componente do exoesqueleto duro de aproximadamen- te 1 milhão de espécies de artrópodos, por exemplo, insetos, la- gostas e caranguejos. Provavelmente, depois da celulose, é o po- lissacarídeo mais abundante na natureza, 235 Os glicosaminoglicanos são componentes da matriz extracelular O espaço extracelular nos tecidos de animais multicelulares é preenchido com um material geleificado, chamado de matriz extracelular ou, ainda, conhecido como substância fundamen- tal, que mantém unidas as células de um tecido e fornece uma via porosa para a difusão dos nutrientes e do oxigênio para as células individuais. A matriz extracelular é composta por uma rede de heteropolissacarídeos e por proteínas fibrosas interco- nectadas, tais como colágeno, elastina, fibronectina e laminina. Os heteropolissacarídeos, chamados glicosaminoglicanos, são uma família de polímeros lineares compostos por unidades re- “petitivas de dissacarídeos (Fig. 9-20). Um dos monossacarídeos que compõem os dissacarídeos é sempre a N-acetilglucosamina ou a N-acetilgalactosamina, o outro na maioria dos casos é um ácido urônico, usualmente o ácido D-glucurônico ou o ácido L-idurônico. Em alguns glicosaminoglicanos, uma ou mais das hidroxilas do açúcar aminado estão esterificadas com sulfato. A combinação desses grupos sulfato e dos grupos carboxilato das unidades de ácido urônico dá aos glicosaminoglicanos uma alta densidade de cargas negativas. Para minimizar as forças repulsi- vas entre os grupos carregados vizinhos, essas moléculas assu- mem uma conformação estendida quando em solução. O pa- Glicosamino- Dissacarídeo repetitivo Nº de dissaca- glicano rídeos por cadeira Hialuronato * oo D, o HA =50,000 HH ) H H OH Glea GleNAC Condroitina 4-sulfato 20-60 H 5; Ga GalNAcASO; Queratana CH,OSO; sulfato —o Ê cH,0H H Os H -25 2H Ho 4 aa H A, o H H po H OH | CH, Gal GIeNACOSO; Figura 9-20 - Unidades repetitivas de alguns glicosaminoglicanos comuns da matriz extracelular. As moléculas são copolimeros alternan- do unidades de ácido urônico e de aminoaçúcar com ésteres de sulfato em algumas das posições. Os grupos ionizados, carbosilato e sulfato (verme- lho), dão a esses polímeros sua caracteristica de carga altamente negativa. 237 drão característico das unidades sulfatadas e não-sulfatadas nos glicosaminoglicanos estabelece um reconhecimento específico por uma variedade de proteínas ligantes que se associam ele- trostaticamente com essas moléculas. Os glicosaminoglicanos estão ligados a proteínas extracelulares para formar os proteo- glicanos (discutidos a seguir). O glicosaminoglicano ácido hialurônico (hialuronato em pH fisiológico) contém unidades alternadas de ácido D-glucurô- nico e N-acetilglucosamina (Fig. 9-20). Os hialuronatos têm massas moleculares acima de 1 milhão; tendo até 50.000 unida- des repetitivas compostas por dissacarídeos. Eles formam solu- ções altamente viscosas e claras, as quais funcionam como lu- brificantes nos fluidos sinoviais das juntas e conferem ao hu- mor vítreo dos olhos dos vertebrados sua consistência semelhante à da gelatina (do grego hyalos que significa “vidro”; hialurona- tos podem ter uma aparência vítrea ou translúcida). O hialuro- nato é também um componente essencial da matriz extracelular das cartilagens e dos tendões, nos quais contribui para sua elas- ticidade característica e para a resistência à tensão, como resul- tado das fortes interações que apresenta com os outros compo- nentes da matriz. A hialuronidase, uma enzima secretada por algumas bactérias patogênicas (causadoras de doenças), pode hidrolisar as ligações glicosídicas do hialuronato, tornando os tecidos mais suscetíveis à invasão pelas bactérias. Em muitos organismos, uma enzima semelhante, no esperma, hidrolisa uma capa externa de glicosaminoglicano que envolve o óvulo, per- mitindo a penetração do espermatozóide. Outros glicosaminoglicanos diferem do hialuronato em dois aspectos: eles são geralmente polímeros muito curtos e covalen- temente ligados a proteínas específicas (proteoglicanos, discuti- dos a seguir). A condroitina sulfato (do grego chondros que si nifica “cartilagem”) contribui para a resistência à tensão na car- tilagem, tendões, ligamentos e paredes da aorta. A dermatana sulfato (do grego derma, “pele”) colabora com a flexibilidade da pele e também está presente nos vasos sangúíneos e válvulas do coração. A queratina sulfato (do grego keras, “cornos”) não pos- sui ácido urônico e a quantidade de sulfato é variável. Esse polí- mero está presente na córnea, cartilagens, ossos e em uma varie- dade de estruturas calosas formadas por células mortas: chifres, cabelos, cascos, unhas e garras. A heparina (do grego hêpar, “fi- gado”) é um anticoagulante natural, sintetizado nos mastócitos e liberado no sangue, no qual inibe a coagulação pela ligação e estimulação simultânea da proteína anticoagulante, antitrom- bina III. A interação entre a heparina e a antitrombina III é for- temente eletrostática; a heparina é a macromolécula biológica mais bem conhecida, que contém a mais alta densidade de carga negativa. A heparina purificada é rotineiramente adicionada a amostras de sangue colhidas para análises clínicas e sangue doa- do para transfusão, a fim de evitar a coagulação. A Tabela 9-2 resume a composição, as propriedades, a ocor- rência e a função dos polissacarídeos descritos neste tópico. Glicoconjugados: Proteoglicanos, Glicoproteínas e Glicolipídios Além das suas importantes funções como armazenamento ener- gético (amido, glicogênio) e materiais estruturais (celulose, quiti- na, peptideoglicano), os polissacarídeos (e oligossacarídeos) são portadores de informações, eles servem como indicadores de en- dereçamento para algumas proteínas e como mediadores para in- terações específicas célula-célula e célula-matriz extracelular. As moléculas que contêm carboidratos específicos agem no reconhe- cimento e na adesão célula-célula, na migração celular durante 238 Tabela 9-2 - Estrutura e função de alguns polissacarídeos Tamanho (número Polímero Tipo” Unidade repetitiva! de unidades Funções monossacarídicas) Amido Armazenagem de energia: nos vegetais Amilose Homo fa: 1-+4)GIc, linear 50-5.000 Amilopectina Homo fa:1-»4)Glc,com Até 10º ramos (a1»6)Glc a cada 24-30 unidades Glicogênio Homo (71-54)GIc, com até 50.000 Armazenagem de energia: nas bactérias e células ramos (a 1 »6)Glc animais a cada 8-12 unidades Celulose Homo (B1-+4)Glc Até 15.000 Estrutural; nos vegetais, fornece ngidez e resistência às paredes celulares Quitina Homo (B1-+4)GIcNAC Muito grande Estrutural; em insetos, aranhas e crustáceos, fornece rigidez e resistência ao exoesqueleto Peptideoglicano Hetero; 4)MurZAc(p1-+4) Muito grande Estrutural; em bactérias, fornece rigidez e ligado com GINAc(p1 resistência ao envoltório celular peptídeo Hialuronato Hetero; AGIA 3] até 100.000 Estrutural: em vertebrados, matriz extracelular da glicosaminoglicano) ácido GINAcp1 pele e tecido conjuntivo; viscosidade e lubrificação nas articulações * Cada polimera é classificado como homopolissacarídeo (homo) ou como heteropolissacarídeo (hetero). *Os nomes abreviados para os peptideaglicanos e hialuronato indicam que o polimero contém unidades repetitivas de dissacarideos, com o GICNAC de uma unidade dissacarídica ligada (1 -» 4) à primeira unidade da próxima unidade dissacarídica desenvolvimento, no revestimento sangúíneo, na resposta imune ena cicatrização de lesões, apenas para citar algumas de suas fun- ções. Na maioria dos casos, o carboidrato sinalizador é covalente- mente ligado à proteína ou ao lipídio para formar um glicoconju- gado, que é a molécula biologicamente ativa. Proteoglicanos são macromoléculas da superfície da célula ou da matriz extracelular, nos quais uma ou mais cadeias de gli- cosaminoglicanos estão ligadas covalentemente a uma proteína de membrana ou a uma proteína secretada. A parte formada pelo glicosaminoglicano comumente é a maior fração (em massa) da molécula de proteoglicano, predominando na estrutura, e é fre- quentemente o principal sítio da atividade biológica. Em mui tos casos, a atividade biológica é resultante de múltiplos sítios de ligação, ricos em possibilidades de realizar pontes de hidro- gênio e interações eletrostáticas com outras proteínas da super- ficie celular ou da matriz extracelular. Os proteoglicanos são o maior componente dos tecidos conectivos, tais como cartilagens, nos quais suas muitas interações não-covalentes com outros pro- teoglicanos, proteínas e glicosaminoglicanos produzem rigidez e elasticidade. Glicoproteínas têm um ou vários oligossacarídeos de com- plexidade variada ligados covalentemente à proteína. Elas são encontradas no lado externo da membrana plasmática, na ma- triz extracelular e no sangue. No interior das células, elas são encontradas em organelas específicas, tais como o complexo de Golgi, os grânulos secretores e os lisossomos. As frações oligos- sacarídicas das glicoproteínas são menos repetitivas do que as ca- deias de glicosaminoglicanos dos proteoglicanos, elas são ricas em informações, compondo sítios altamente específicos para o reco- nhecimento e a ligação de alta afinidade com outras proteínas. Glicolipídios são lipídios de membrana nos quais os gru- pos hidrofílicos da “cabeça” são oligossacarídeos, que, como nas glicoproteínas, agem como sítios específicos para o reconheci- mento pelas proteínas ligadas a carboidratos. A glicobiologia, o estudo da estrutura e função de glicocon- jugados, é uma das mais ativas e excitantes áreas da bioquímica e da biologia celular. Nossas discussões utilizam apenas alguns poucos exemplos para ilustrar a diversidade estrutural e de ati- vidades biológicas dos glicoconjugados. No Capítulo 20, discu- tiremos a biossíntese dos polissacarídeos, incluindo os glicosa- minoglicanos e, no Capítulo 27, a síntese das cadeias oligossa- carídicas das glicoproteínas. Os proteoglicanos são macromoléculas da superfície celular e da matriz extracelular, contendo glicosaminoglicanos As unidades básicas dos proteoglicanos consistem em uma “pro- teína central” ligada covalentemente a glicosaminoglicano(s). Por exemplo, a estrutura laminar da matriz extracelular (lâmina ba- sal), que separa grupos organizados de células, contém uma fa- mília de proteínas centrais (M, de 20.000 a 40.000), cada uma com várias cadeias de heparana sulfato ligadas covalentemente. A heparana sulfato é estruturalmente similar à heparina, porém apresenta menor grau de esterificação por grupos sulfato. O ponto de ligação é comumente um resíduo de Ser, no qual o glicosaminoglicano é ligado por meio de pontes feitas por um trissacarídeo (Fig. 9-21). O resíduo de Ser é geralmente da se- qiência -Ser-Gly-X-Gly- (onde X é qualquer resíduo de ami- noácido), embora, nem todas as proteínas com essa sequência tenham glicosaminoglicano ligado. Muitos proteoglicanos são secretados na matriz extracelular, mas alguns são proteinas in- tegrais de membrana. Por exemplo, a proteína central sindecana (M, 56.000) tem um domínio transmembrana único e um do- mínio extracelular de reconhecimento formado por trés cadeias de heparana sulfato e duas de condroitina sulfato ligadas, cada (Bi) (Bla (BIS3APIS3 (pia) I Condroitina sulfato : Proteína central > Figura 9-21 — A estrutura de um proteoglicano mostrando a ponte de trissacarídeo. Um trissacarídeo Igante típico (azul) conecta um gii- cosaminoglicano — no caso a condroitina sulfato (laranja) — por meio de um resíduo de Ser (vermelho) da proteina central. A unidade de xilose do terminal redutor do ligante é unida pelo seu carbono anomérico ao grupo hidroxila do resíduo de Ser. 240 tracelulares. Essas interações não servem meramente para anco- rar as células à matriz extracelular, mas também para produzir vias que dirigem a migração das células nos tecidos em desen- volvimento e, por meio das integrinas, transportam informa- ções em ambas as direções por meio da membrana plasmática. Os proteoglicanos da matriz são essenciais na resposta das células a certos fatores de crescimento extracelulares, Por exem- plo, o fator de crescimento do fibroblasto (FGF), uma proteína sinalizadora extracelular que estimula a divisão celular, primei- ro se liga a uma porção da heparana sulfato das moléculas de sindecana na membrana plasmática da célula-alvo. A sindecana, então, “apresenta”o FGF ao receptor específico para FGF na mem- brana plasmática, e somente assim o FGF pode interagir produti- vamente com esses receptores para iniciar a divisão celular. As glicoproteínas são conjugados ricos em informações contendo oligossacarídeos As glicoproteínas são conjugados de carboidrato-proteína nos quais a parte dos carboidratos são menores e estruturalmente mais variados do que os glicosaminoglicanos nos proteoglica- nos. Os carboidratos estão ligados com o grupo hidroxila do resíduo de Ser ou Thr (O-ligados) por meio do carbono anomé- rico por ligação glicosídica, ou por meio de ligação N-glicosí ca com o nitrogênio da função amida de um resíduo de Asn (N- ligado) (Fig. 9-25). Algumas glicoproteinas têm uma cadeia ol gossacarídica simples, mas muitas têm mais do que uma; o carboidrato pode constituir de 19% a 70% ou ainda mais da massa da glicoproteína. As estruturas de um grande número de oligos- sacarídeos O e N-ligados de uma variedade de glicoproteínas são conhecidas; a Figura 9-25 mostra alguns exemplos típicos. Como veremos no Capítulo 12, a superficie externa da mem- brana plasmática tem muitas glicoproteínas unidas covalente- mente a oligossacarídeos de complexidade variada. Uma das gli- coproteínas de membrana mais bem caracterizadas é a glicofo- rina A da membrana de eritrócito (veja Fig. 12-10). Ela contém 60% da massa em carboidrato, formado por 16 cadeias oligos- sacarídicas (totalizando 60 a 70 unidades monossacarídicas) covalentemente ligadas a resíduos de aminoácidos próximos ao aminoterminal da cadeia polipeptídica. Quinze das cadeias oli- gossacarídicas são O-ligadas a resíduos de Ser ou Thr, e uma é N-ligada a um resíduo de Asn. Muitas das proteínas secretadas pelas células eucarióticas são glicoproteínas, incluindo a maioria das proteínas do sangue. Por exemplo, imunoglobulinas (anticorpos) e certos hormônios, tais como hormônio estimulante do folículo, hormônio luteinizan- te e hormônio estimulante da tireóide. Muitas proteínas do lei- te, incluindo a lactoalbumina e algumas proteínas secretadas pelo pâncreas (por exemplo a ribonuclcase), são glicosiladas, bem como a maioria das proteínas contidas em lisossomos. Às vantagens biológicas da adição de oligossacarídeos às proteínas não estão completamente entendidas. Os agregados altamente hidrofílicos de carboidratos alteram a polaridade e a solubilidade das proteínas com as quais estão conjugados. As cadeias de oligossacarídeos ligadas a proteínas recém-sintetiza- das no complexo de Golgi podem também influenciar a sequên- cia dos eventos que se processam durante o enovelamento do polipeptídeo que determina a estrutura terciária da proteína (veja Fig. 27-36). As interações estéricas entre peptídeo e oli- gossacarídeo podem impedir uma rota de enovelamento e fa- vorecer outra. Quando numerosas cadeias de oligossacarídeos carregados negativamente são agregadas em uma única região de uma proteína, a repulsão entre as cargas favorece então, nes- sa região, a formação de uma estrutura estendida, semelhante a um bastão. A densidade e a carga negativa das cadeias de oligos- sacarídeos também protegem algumas proteínas do ataque de enzimas proteolíticas. Além desses efeitos fisicos globais sobre a estrutura protéi- ca, existem efeitos biológicos mais específicos das cadeias de oli- gossacarídeos nas glicoproteínas. Já havíamos notado anterior- mente as diferenças entre as sequências lineares ricas em infor- mações dos ácidos nucléicos e das proteínas e a regularidade monótona dos homopolissacarídeos como a celulose (veja Fig. 3-23). Os oligossacarídeos ligados às glicoproteínas em geral não são monótonos, porém são muito ricos em informação estrutu- Figura 925 - Oligossacarídeos ligados em glicoproteínas. (a) Os oligossacaríde- os O-ligados têm uma ligação glicosídica do grupo hidroxila dos resíduos de Ser ou Thr (sombreado em rosa), representada aqui por GalNAc como açúcar no terminal redutor do oligossacarídeo. Uma cadeia simples e uma cadeia complexa são mos- tradas. (Relembre que NeuSAc — ácido N- acetilneuramínico — é comumente chama- do ácido siálico). (b) Os oligossacarideos Nº- ligados têm uma ligação N-glicosil ao nitrogênio da amida no resíduo de Asn (sombreado em verde), representado aqui por GleNAc como açúcar terminal. Três ti- GalNAC pos comuns de cadeias oligossacarídicas que são Nligadas nas glicoproteinas são Exemplos: mostradas. Uma descrição completa da es- trutura do oligossacarídeo requer especifi- cação da posição e estereoquímica (a ou f) de cada ligação glicosídica (a) O-ligados NH ds He GleNAc Asn Exemplos: O GloNac O Man Ocal O NeusAc YFuc | VW GalNAc ral. Considere as cadeias de oligossacarídeos na Figura 9-25, elas são típicas daquelas encontradas em muitas glicoproteínas. As mais complexas delas mostram conter 14 unidades de monos- sacarídeos, de quatro tipos diferentes, variadamente unidos en- tre si com ligações (152), (153), (154), (156), (253) e (2-6), alguns com a configuração « e outros com a configura- ção . O número de permutações e combinações possíveis dos tipos de monossacarídeos e de ligações glicosídicas em um oli- gossacarídeo desse tamanho é astronomicamente grande. Cada um dos oligossacarídeos na Figura 9-25 apresenta, portanto, uma face única, reconhecível por enzimas e por receptores que inte- ragem com ele. Inúmeros » conhecidos nos quais a mes- ma proteína produzida em dois tecidos tem diferentes padrões de glicosilação. A proteina humana interferon IEN-B1, produ- zido nas células do ovário, por exemplo, tem cadeias oligossa- carídicas que diferem da mesma proteina produzida em células epiteliais mamárias. O significado biológico dessas glicoformas teciduais não é conhecido, mas de alguma maneira as cadeias oli- gossacarídicas representam um marcador específico de tecido. sos Glicolipídios e lipopolissacarídeos são componentes de membrana As glicoproteinas não são os únicos componentes celulares que possuem cadeias de oligossacarídeos complexos; alguns lipídios também contêm cadeias de oligossacarídeos ligadas covalente- mente. Os gangliosídeos são lipídios de membrana das células de eucarioto, nos quais, o grupo da cabeça polar, a parte do lipí- n210 241 dio que forma a superficie externa da membrana, é um oligossa- carídeo complexo contendo ácido siálico (Fig. 9-9) e outras uni- dades de monossacarídeos. Alguns dos oligossacarídeos que fa- zem parte dos gangliosídeos, tais como aqueles que determi- nam os grupos sangúíneos humanos (veja Fig. 11-12), são idênticos àqueles encontrados em certas glicoproteínas que, consequentemente, também contribuem para a determinação dos tipos de grupos sangúíneos. Como os oligossacarídeos que fazem parte das glicoproteinas, esses lipídios de membrana são geralmente, talvez sempre, encontrados na face externa da mem- brana plasmática. Lipopolissacarídeos são os componentes principais da mem- brana externa de bactérias gram-negativas como a E. coli e a Sal- monella typhimurium. Eles são os alvos primários dos anticor- pos produzidos pelo sistema imunológico de vertebrados em resposta à infecção bacteriana e são, consequentemente, impor- tantes determinantes do sorotipo de linhagens de bactérias (so- rotipos são linhagens distintas com base nas suas propriedades antigênicas). Os lipopolissacarídeos da S. typhimurium contêm seis ácidos graxos ligados a duas unidades de glucosamina, um dos quais é um ponto de ligação para um oligossacarídeo com- plexo (Fig. 9-26). A E. coli é semelhante, mas apresenta apenas um lipopolissacarídeo. Os lipopolissacarídeos de algumas bac- térias são tóxicos para o homem e outros animais; por exemplo, eles são responsáveis pela perigosa hipotensão sanguínea que ocorre na sindrome do choque tóxico, resultante de infecções provocadas por bactérias gram-negativas. Cadeia “O-específica” Oligossacarídeo central Lipídio Figura 9-26 - Lipopolissacarídeos bacterianos. (a) Diagrama esque- mático do lipopolissacarídeo da membrana externa de Salmonella typhi- murium. Kdo é o ácido 3-desoxi-D-mano-ocutulosônico, previamente chamado de ácido cetodesoxioctônico; Hep é o L-glicero-D-mano-hepto- se, AbeOAc é a abequose (a 3,6-didesoxi-hexose) acetilada em uma de suas hidroxilas. Existem seis ácidos graxos na porção de lipídio A da mo- lécula. Diferentes espécies bacterianas têm diferenças sutis na estrutura dos hpopolissacarídeos, mas elas têm em comum uma região lipídica (li- pídio A), um oligossacarídeo central e uma cadeia “O-específica”, a qual é o principal determinante do sorotipo (imunorreativo) da bactéria. Fora da membrana da bactéria gram-negativa S. typhimunum e E. coli estão contidas muitas moléculas lipopolissacarídicas, sendo a superfície das células virtualmente coberta de cadeias O-específicas. (b) Modelo mole- cular espaço-cheio do lipopolissacarídeo de £. coli. A posição da sexta cadeia de ácidos graxos não foi definida no estudo cristalográfico, por ta) isso não está mostrada, 243 Glicoproteina ligante de integrina Integrina P-selectina Célula do endotélio capilar Glicoproteína ligante de P-selectina EE | Figura 9-27 - Função das interações lectina-ligante no movimento de linfócitos nos sítios de uma infecção ou lesão. A «circulação de um linfócito T através de um capilar é diminuída pelas interações transitórias entre as moléculas de selectina P da membrana plasmática das células endoteliais do capilar e as proteinas com sítios de ligação para selectina P na superfície das células T. Por meia dessas interações com as moléculas de selectina P sucessivas, as células T rolam ao longo da superfície do capilar. Próximo ao sítio de inflamação, interações fortes entre integrina da superfície do capilar e seus ligantes na superfície de célula T conduzem a uma firme adesão. As células T param a rolagem e, sob a influência de sinais enviados de fora do sitio de inflamação, inicia-se um extravasamento através da parede do capilar — com isso, movem-se em direção ao sítio de inflamação. Figura 9-28 — As células de Helicobacter pylori aderidas à superfície gás- trica. Essa bactéria causa úlcera pelas interações entre a lectina da super- fície da bactéria e a oligossacarideo Le* (um antigeno de grupo sangúí- neo) do epitélio gástrico. podem ligar e entrar nas células, mas elas podem elicitar uma resposta imune que poderia proteger o recipiente quando ex- posto à toxina natural. Várias viroses animais, incluindo o vírus da influenza, li- gam-se a suas células hospedeiras por meio de interações com oligossacarídeos expostos na superfície da célula do hospedeiro. A lectina do vírus da influenza é a proteína HA, a qual descreve- remos no Capítulo 12. A proteína HA é essencial para a entrada ea infecção viral (veja págs. 313-316 e 12-21). A Figura 9-29 resume os tipos de interações biológicas que são mediadas por ligação a carboidratos. Análise de Carboidratos A crescente tomada de consciência da importância da estrutura de oligossacarídeos no reconhecimento biológico tem sido o promotor do desenvolvimento de métodos de análise da estru- tura e estereoquímica de oligossacarídeos complexos. A análise de oligossacarídeos é complicada pelo fato de que, diferentemente dos ácidos nucléicos e proteínas, os oligossacarídeos podem ser ramificados e unidos por uma variedade de ligações. Para serem analisados, os oligossacarídeos são geralmente removidos das proteínas ou lipídios conjugados e então submetidos a etapas de degradação com reagentes específicos que revelam a posição das ligações ou a estereoquímica. A espectrometria de massa e a es- pectroscopia RMN também têm se tornado inestimáveis para decifrar a estrutura de oligossacarídeos. Cadeia de oligossacarídeo Proteína da membrana plasmática P-selectina Figura 9-29 - Funções dos oligossacarídeos no reconhecimento e na adesão à superfície da célula (a) Os oligossacarideos com estruturas únicas (representados por uma cadeia de bolas coloridas), componentes de uma variedade de glicoproteinas ou glicolipídios na superficie externa da membrana plasmática, interagem com alta especificidade e afinidade com as lectinas do meio extracelular. (b) As viroses que infectam as células animais, tais como o virus da influenza, ligam-se às glicoproteinas na célula como primeira etapa de infecção. (e) Toxinas bacterianas, tais como a toxina pertussis escolérica, ligam-se a um glicolipídio de superfície antes de entrar na célula. (d) Algumas bactérias, tais como Helicobacter pylori, aderem para, então, colonizar ou infectar as células animais. (e) As lectinas chamadas selectinas da membrana plasmática de certas células medeiam as interações célula-célula, tais como as dos linfócitos T com células endoteliais da parede dos capilares no sitio de infecção. Os oligossacarídeos componentes de glicoproteínas ou gli- colipídios podem ser liberados por enzimas purificadas — gli- cosidases que removem especificamente oligossacarídeos O ou N-ligados, ou lipases que removem os grupos lipídicos da cabe- ça. As misturas de carboidratos são fracionadas em seus com- ponentes individuais (Fig. 9-30) por muitas das mesmas téci cas utilizadas na separação de proteínas e aminoácidos: preci tação fracionada com solventes, cromatografia de troca iônica e cromatografia de filtração em gel (exclusão molecular) (veja Fig. 5-18). As lectinas altamente purificadas, ligadas covalentemente a um suporte insolúvel, são comumente usadas na cromatogra- fia de afinidade de carboidratos (veja Fig. 5-18c). A hidrólise de oligossacarídeos e polissacarídeos em solução de ácido forte li- bera uma mistura de monossacarídeos que, depois da conver- são para derivados voláteis convenientes, pode ser separada, iden- tificada e quantificada por cromatografia gás-líquida (veja pág. 298), esclarecendo a composição completa do polímero. Por exemplo, para polímeros lineares simples como a amilose, as posições das ligações glicosídicas entre os monossacarídeos são determinadas pelo tratamento do polissacarídeo intacto com io- deto de metila em meio fortemente básico para converter todas as hidroxilas livres para os respectivos éteres metílicos, que são estáveis em meio ácido, então o polissacarídeo metilado é hi- drolisado em meio ácido. Somente os grupos hidroxila livres presentes no monossacarídeo resultante são aqueles que esta- vam envolvidos nas ligações glicosídicas. Para determinar a se- quência das unidades monossacarídicas, incluindo ramificações se elas estiverem presentes, são utilizadas exoglicosidases de es- pecificidade conhecida, para remover as unidades uma de cada vez do terminal(is) não-redutor(es). A especificidade dessas exo- glicosidases frequentemente permite deduzir a posição e a este- reoquímica das ligações. A análise dos oligossacarídeos está cada vez mais ligada à espectroscopia de RMN de alta resolução e espectrometria de massa. As análises de RMN isoladas, especialmente para oligos- sacarídeos de tamanho médio, podem fornecer muitas informa- ções sobre a segiência, a posição da ligação e a configuração do carbono anomérico. Os polissacarídeos e os oligossacarídeos grandes podem ser tratados quimicamente ou com endoglicosi- dases para romper ligações glicosídicas internas específicas, pro- duzindo vários oligossacarídeos menores, mais facilmente ana- lisáveis. Processos automáticos e instrumentos comerciais são usados para as determinações de rotina da estrutura de oligos- sacarídeos, mas a sequência de ramificação dos oligossacarídeos unidos por mais de um tipo de ligação continua sendo um pro- blema mais interessante do que determinar a sequência linear de proteínas e ácidos nucléicos, os quais têm monômeros liga- dos apenas por um tipo de ligação.